Долговременная динамика видов лесных растений с различной шириной экологических ниш на постоянной пробной площади в сосняке с дубом и липой в Воронежском заповеднике

Автор: Зубкова Елена Владимировна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общие проблемы экологии
Статья в выпуске: 5-1 т.14, 2012 года.
Бесплатный доступ
Рассмотрены изменения видового состава лесных растений на постоянной пробной площади, наблюдавшейся 61 год в Воронежском заповеднике; проведено определение параметров местообитания по шкалам Г.Элленберга и Д.Н. Цыганова, а также проанализирована динамика стенобионтных видов. Показано, что по факторам увлажненности и кислотности почв существенных изменений условий на пробной площади не обнаружено, уменьшения доли растений-стенобионтов по этим факторам среды не наблюдалось; выявлено увеличение богатства почв азотом в результате жизнедеятельности биоценоза и замещение растений более бедных мест обитания на растения более богатых мест обитания.
Экологические шкалы растений, экологические факторы, лесные сукцессии, стенобионты, эврибионты
Короткий адрес: https://sciup.org/148201474
IDR: 148201474 | УДК: 57.055
Long-term dynamics of species of forest plants with different width of ecological niches at permanent plot in pine forest with oaks and lime in Voronezh nature reserve
Changes of species composition of forest plants at permanent plot established 61 year ago in Voronezh Strict Nature Reserve is discussed. Evaluation of site parameters using Ellenberg and Tsyganov ecological scales has been done. Dynamics of stenobionts (specialists) during these years is also described. It is shown that there are no significant changes in soil moisture and acidity, stenobionts portion in relation to these factors is constant in time. Nevertheless soil fertility increased being a result of biocoenos life activity. Plant species specific for more rich sites substitute those ones specific for poor sites.
Текст научной статьи Долговременная динамика видов лесных растений с различной шириной экологических ниш на постоянной пробной площади в сосняке с дубом и липой в Воронежском заповеднике
Д анные многолетних наблюдений на постоянных пробных площадях в лесных сообществах являются уникальными и позволяют проверить гипотезы о направленности процессов при сукцессионных изменениях в лесных сообществах.
Нами ранее [1, 2] были проанализированы данные по изменениям видового состава лесных сообществ растений на разных сукцессионных стадиях в: Печоро-Илычском заповеднике, в лесах на северо-востоке Костромской области и в заповеднике «Брянский лес».
В работе была использована программа EcoScaleWin [3, 4], позволившая получить сводную информацию об экологических условиях изучаемого участка с использованием экологических шкал Д.Н. Цыганова [5] и Г. Элленберга [6]. Экологические шкалы устроены таким образом, что каждый фактор среды в диапазоне условий существования растений разделен на несколько градаций (например, по фактору увлажненности почв от пустынного типа увлажнения до водного всего 23 градации). Для каждого растения в балловом выражении определяется оптимум (шкалы Г. Элленберга) или диапазон условий (шкалы Д.Н. Цыганова) существования вида. Таким образом, мы можем экологическую нишу каждого растения охарактеризовать набором балловых величин по разным факторам среды, что позволяет как сравнивать экологические ниши видов, так и по списку видов получать балловую характеристику экотопа по факторам среды, представленным в шкалах.
Так как шкалы Д.Н. Цыганова являются диапазонными, это позволяет нам разделить все виды на две группы: с узким диапазоном по данному фактору среды – стенобионты или специалисты и остальные – эврибионты или генералисты [7]. Такое деление дает возможность исследовать структурный состав растительных сообществ разных сук-
Зубкова Елена Владимировна , с.н.с., e-mail:
цессионных стадий, в том числе с точки зрения присутствия в них видов широкой и узкой экологической амплитуды и, зная тенденции изменения характеристик экотопа, составлять прогноз об изменении состава сообщества. Нами были предложены следующие критерии выделения стенобионт-ных видов: шкала увлажнения почв (Hd) описывает изменение фактора 23 баллами, по ней к стеноби-онтным были отнесены виды с шириной диапазона не более 7 баллов в любой части шкалы; ширина шкалы богатства почв азотом (Nt) – 11 баллов, сте-нобионтные - виды с шириной диапазона не более 5; шкала кислотности почв (Rc) 13 баллов, стено-бионтные – с шириной диапазона не более 5 баллов; шкала освещённости-затенения (Lc) – 11 баллов, стенобионтные виды – с диапазоном не более 4 баллов. Анализ свойств шкал Д.Н. Цыганова опубликован ранее [8].
Анализ динамики стенобионтных видов1 на трёх ранее рассмотренных территориях показал, что 1) по ходу сукцессии увеличивалась доля стенобион-тов по фактору увлажненности почв и 2) отсутствовало закономерное изменение относительного числа стенобионтов по фактору богатства почв азотом [1]. Было найдено, что изменение экотопиче-ских условий по факторам увлажненности почв и богатства почв азотом, оцененные по шкалам Д.Н.Цыганова [5] и Г.Элленберга [6], в ходе сукцессии показывают сходные тенденции. Так, по фактору увлажненности почв от первой к последней из изученных стадий экотопические условия остались постоянными; по фактору богатства почв проведенный анализ по шкалам обоих авторов показал, что произошло увеличение богатства почв. Это было показано как при расчете по общему списку видов сообществ, так и по изменению положения среднего значения левой границы диапазонов у стенобионтных видов. Выяснилось, что средние значения левых границ обитания стенобионтных видов по фактору увлажненности почв не меняются, а по фактору богатства почв азотом левая граница смещается в сторону увеличения богатства почвы. При эндогенных сукцессиях, приводящих к изменениям свойств среды под влиянием жизнедеятельности видов, слагающих сообщество, само сообщество также претерпевает изменения, при которых в нём будут накапливаться виды-стенобионты по фактору среды, не претерпевающему существенных изменений (в нашем случае по фактору увлажненности почв). Виды, стенобионты по фактору среды, претерпевающему изменения, будут частично выпадать из сообщества, теряя свои позиции при изменениях среды.
С целью обнаружения сходства полученных ранее результатов при обработке большого количества геоботанических описаний с динамикой одного и того же растительного сообщества в его многолетнем развитии, был проведён анализ динамики стенобионтных видов на постоянной пробной площади. По причине большой длительности сукцессионных стадий в лесных сообществах получить данные об изменениях видового состава на закартированных пробных площадях в ходе нескольких сукцессионных стадий крайне сложно. В данной работе нами были проанализированы уникальные материалы, собранные на постоянной пробной площади, заложенной в 1937 году в Воронежском заповеднике – ПП No.10 (816 м2) [10, 11] и любезно предоставленные сотрудником заповедника Е.В. Стародубцевой. Растительный покров на этой пробной площади развивался без значительных антропогенных воздействий в течение 61 года, за которые сообщество прошло один сукцессионный шаг - из сосняка с дубом и липой мелкотравного преобразовалось в сосняк с дубом и липой сложный (бореально-неморальный), классифицированный нами по геоботаническим описаниям согласно базе данных «Ценофонд» Л.Б.Заугольновой и О.В.Морозовой [9].
ИСТОРИЯ ИССЛЕДОВАНИЙ ПРОБНОЙ ПЛОЩАДИ ДРУГИМИ АВТОРАМИ
Описание пробной площади приводится по статье Л.С. Травниковой [10] и публикации Е.А. Стародубцевой с соавторами [11].
Пробная площадь была заложена и закартирована в 1937 г. ботаником заповедника М.В. Николаевской в сосняке дубняковом 110-ти лет на бурой лесной песчаной почве. Пробная площадь расположена на второй надпойменной террасе реки Воронеж на вершине песчаного бугра и занимает его пологий северо-западный склон с волнистым рельефом. В предшествующий период формирование современных старых сосняков происходило под пологом дуба, который неоднократно вырубался, и на момент закладки площади сообщество было представлено в первом ярусе сосной обыкновенной ( Pinus sylvestris L.), во втором – дубом черешчатым
(Quercus robur L.) с примесью берёзы повислой (Betula pendula Roth) и клёна платановидного (Acer platanoides L.). Полнота насаждения была низкой. Так как кроны практически не перекрывались, то освещённость под пологом была достаточной для роста многовидового подроста, в состав которого входили: клен платановидный (Acer platanoides L.), клён татарский (Acer tataricum L.), берёза повислая (Betula pendula Roth), крушина ломкая (Frangula alnus Mill.), дуб черешчатый (Quercus robur L.), бересклет бородавчатый (Euonymus verrucosa Scop.), рябина обыкновенная (Sorbus aucuparia L.), липа сердцевидная (Tilia cordata Mill.), тополь дрожащий (Populus tremula L.). В нижнем ярусе было отмечено большое число боровых и луговых видов [11]. Кроме геоботанического описания растительности были составлены картограммы распределения на площади древесных растений и травяно-мохового покрова [10]. В 1955 году геоботаником И.М. Хомяковой было отмечено разрастание подлеска из липы, в результате чего уменьшилась освещённость на уровне почвы, увеличилась площадь «мертвой подстилки из листьев широколиственных пород», что привело к исчезновению лишайников, уменьшению числа мхов, боровых и луговых видов и увеличению обилия неморальных видов, таких как звездчатка ланцетолистная (Stellaria holostea L.), осока волосистая (Carex pilosa Scop.). Были взяты пробы почвы под разными видами растений, проведён анализ почвенных образцов, в числе прочего, на содержание гумуса [10]. В итоге показано большее содержание гумуса в слое почвы 2-7 см горизонта А1 под растениями неморальной группы, такими как: осока волосистая (9.78%), звездчатка (8,75%), липа (7,9%), бересклет (7,55%) по сравнению с его содержанием под куртинами мха (5.8%) и куртинами бореального вида – костяники (6,82%) [10]. Впоследствии, липа и лещина заняли доминирующее положение во втором ярусе, что привело к существенному уменьшению освещённости, и, как следствие, к практически полному выпадению боровых видов и значительному выпадению луговых видов. Число видов трав сократилось с 59 в 1937 году до 23 в 1988 г. К 1988 г. мхи, наблюдаемые ранее на площади (4 вида в 1927 г., 1 вид в 1955 г.), исчезли полностью. Одновременно с этим происходило разрастание крон сосны. Исследование, проведенное в 2001 г., показало, что по сравнению с 1937 г. число стволов сосны сократилось с 23 до 13 вследствие усыхания, однако, общее проективное покрытие крон за счет их разрастания увеличилось более чем в три раза [11]. Число стволов дуба практически не изменилось однако проективное покрытие крон увеличилось почти в семь раз. Разрастание крон привело к увеличению площади пересечений крон в первом и втором ярусах и усыханию деревьев в третьем ярусе (69%). С 1974 г. на пробной площади наблюдалось полное отсутствие мхов, ранее имевших приуроченность к окнам в древесном пологе. Общее число видов на пробной площади с 1937 к 1998 г. сократилось с 78 до 36 [11].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ ПО ЭКОЛОГИЧЕСКИМ ШКАЛАМ И
ОБСУЖДЕНИЕ
Нами было проведено сравнение тенденций изменения параметров экотопа ПП-10 (освещённость-затенение, увлажненность почв, богатство почв, кислотность почв) по шкалам Д.Н. Цыганова (методом пересечения большинства интервалов) и Г. Элленберга (рис. 1А, Б, В, Г). В работе была использована программа EcoScaleWin [3, 4].
Балловые оценки экотопа на основании списков видов по разным срокам исследования показали сходные тенденции по шкалам Д.Н. Цыганова и Г. Элленберга. По шкале Д.Н. Цыганова по фактору освещённости-затенения с 1937 по 1998 гг. произошло незначительно усиление затенённости - в направлении от режима полуоткрытых пространств (4 балл) к режиму светлых лесов (балл 4.5). По шкале Г. Элленберга также диагностируется некоторое увеличение доли видов, относящихся к «полутеневым растениям» (получающим более 10%, но менее 50% от полной освещенности); так как шкалы освещённости-затенения Д.Н. Цыганова и Г. Элленберга разнонаправленные (по шкале Д.Н. Цыганова балл «1» присвоен самым светлым местообитаниям, а по шкале Г. Элленберга – самым затенённым), то на рисунке 1А результаты по обеим шкалам совпадают. По фактору увлажнённости почв по шкале Д.Н. Цыганова, если сравнивать 1937 и 1998 г., увлажненность почв немного усилилась от сыро-лесолугового типа режима увлажнения (балл 11) в сторону влажно-лесолугового режима увлажнения (балл 12). По шкале Г. Эллен-берга увлажнение значимо не меняется и соответствует увлажнению на свежих почвах (5 балл) (рис 1 Б). Кислотность почв по шкале Д.Н. Цыганова также существенно не меняется – тип режима слабокислых почв (7 балл); по шкале Г. Элленберга условия определяются как благоприятные для растений, обитающих в основном на слабокислых почвах (рис. 1 В). По фактору богатства почв условия меняются наиболее значительно, богатство почв увеличивается (рис. 1Г); по шкале Д.Н. Цыганова от режима бедных азотом почв (балл 5) до режима достаточно обеспеченных азотом почв (балл 7); по шкале Г. Элленберга от бедных почв в сторону почв со средним содержанием минерального азота.
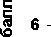
о 5.9
□4
о 5.7
□ 3.5
о 5.5
□ 4.5
о 5.4
□ 4.5
_ □ 12 □ 12.5 □ 12
□ 11
О 4.8 О 4.6 О 5.1 О 5.1
1957 1988 1998
1937 1957 1988 1998
годы наблюдений
годы наблюдений
□Ц о Эл
□Ц о Эл
А
Б
В
□7 □7 □7
О 5.7 о 5.2 96 о 5.4
1957 1988 1998
годы наблюдений
Ei Ц о Эл
Г
Рис. 1. Оценка экотопических условий на пробной площади ПП-10 Воронежского заповедника по шкалам Д.Н.Цыганова (Ц) и Г.Элленберга (Эл): А – шкала освещённости-затенения (Ц) и светолюбия (Эл) (шкалы разнонаправленные по убыванитю-возрастанию затенённости); Б – шкала увлажнённости почв; В – кислотности почв; Г – богатства почв: по горизонтальной оси - годы исследования, по вертикальной – баллы
Таким образом, наиболее существенные изменения на ПП-10 произошли по фактору богатства почв в сторону его увеличения.
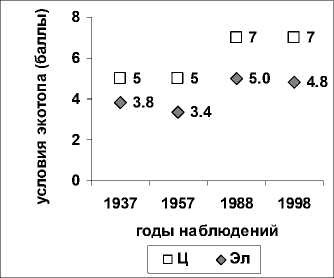
Для оценки изменения структуры сообщества на ПП-10 было проанализировано изменение крайней левой границы диапазонов стенобионтных видов по факторам увлажненности почв, богатства почв азотом и кислотности почв (рис.2). Стенобионтные виды по определённому фактору среды нас интересуют по той причине, что по закону Либиха выживание растений определяет фактор, находящийся в минимуме. Из этого следует, что виды, стеноби- онтные по определённому фактору, и будут наиболее отзывчиво реагировать на изменение этого фактора.
3 10
q to 5
to 5
^ ^
годы
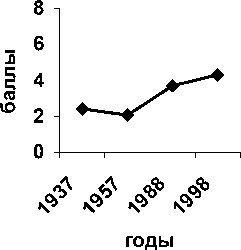
годы
А
Б
В
Рис. 2. Левая граница стенобионтных видов растений на пробной площади ПП-10 Воронежского заповедника по факторам: А – влажности почв (Hd), Б – богатства почв азотом (Nt), В) – кислотности почв; по горизонтальной оси – годы исследования, по вертикальной – баллы по шкалам Д.Н. Цыганова
Таблица 1. Виды стенобионтные по фактору богатства почв азотом (Nt)
|
Год наблюдений |
Диапазон* |
Вид |
||||
|
1937 |
1957 |
1988 |
1989 |
мин |
макс |
|
КУСТАРНИКИ**
|
есть |
есть |
нет |
нет |
1 |
5 |
Genista tinctoria L. |
|
есть |
нет |
нет |
нет |
7 |
11 |
Rubus caesius L. |
|
КУСТАРНИЧКИ |
||||||
|
есть |
есть |
нет |
нет |
1 |
5 1 |
Vaccinium vitis-idaea L. |
|
ТРАВЫ |
||||||
|
есть |
нет |
нет |
нет |
7 |
10 |
Aegopodium podagraria L. |
|
нет |
нет |
есть |
есть |
7 |
10 |
Chelidonium majus L. |
|
есть |
есть |
нет |
нет |
1 |
5 |
Ajuga genevensis L. |
|
есть |
есть |
есть |
есть |
5 |
9 |
Brachypodium sylvaticum (Huds.) see Palisot |
|
есть |
нет |
нет |
нет |
1 |
5 |
Campanula rotundifolia L. |
|
есть |
есть |
есть |
есть |
3 |
7 |
Convallaria majalis L. |
|
есть |
есть |
нет |
есть |
5 |
9 |
Fragaria vesca L. |
|
есть |
нет |
нет |
нет |
1 |
5 |
Hieracium pilosella L. |
|
нет |
есть |
нет |
нет |
1 |
5 |
Hieracium umbellatum L. |
|
есть |
нет |
нет |
нет |
1 |
5 |
Molinia caerulea (L.) Moench |
|
нет |
есть |
есть |
есть |
3 |
7 |
Luzula pilosa (L.) Willd. |
|
есть |
есть |
нет |
нет |
1 |
5 |
Orthilia secunda (L.) House |
|
есть |
есть |
есть |
нет |
1 |
5 |
Peucedanum oreoselinum (L.) Moench |
|
есть |
нет |
нет |
нет |
3 |
7 |
Polygonatum multiflorum (L.) All. |
|
есть |
есть |
есть |
есть |
3 |
7 |
Rubus saxatilis L. |
|
есть |
нет |
нет |
нет |
1 |
5 |
Rumex acetosella L. |
|
есть |
нет |
нет |
нет |
1 |
5 |
Steris viscaria (L.) Raf. |
|
есть |
есть |
нет |
нет |
1 |
5 |
Viola canina L. |
|
есть |
нет |
нет |
нет |
1 |
5 |
Viola hirta L. |
*Диапазон условий жизни видов по шкале богатства почв азотом Д.Н.Цыганова (1983).
**Жизненные формы даны по определителю [12].
Исследование показало менее существенное смещение среднего значения левой границы стено-бионтных видов по факторам увлажненности почв (рис. 2А) и кислотности почв (рис. 2В) по сравне- нию с фактором богатства почв азотом (рис.2Б) По фактору богатства почв (табл. 1) смещение границы произошло в сторону более богатых мест обитания; что является следствием исчезновения с площадки значительного числа стенобионтных видов более бедных мест обитания (13 видов), например, таких как, дрок красильный (Genista tinctoria L.) и ортилия однобокая (Orthilia secunda (L.) House) и внедрением видов, обитающих в богатых почвенных условиях, таких как: чистотел большой (Chelidonium majus L.), ожика волосистая (Luzula pilosa (L.) Willd.) (табл. 1). Древесных видов стенобионтов по фактору богатства почв азотом отмечено не было (Таб. 3).
Расшифровка балловых обозначений шкалы богатства почв азотом Д.Н. Цыганова (1983) приводится в таблице 2. Буквенные обозначения в таблице для удобства заменены цифрами [4].
Таблица 2. Шкала богатства почв азотом (Nt)
|
Тип режима |
Балл |
|
Безазотных почв |
1 |
|
Промежуточный между 1 и 3 |
2 |
|
Очень бедных азотом почв |
3 |
|
Промежуточный между 3 и 5 |
4 |
|
Бедных азотом почв |
5 |
|
Промежуточный между 5 и 7 |
6 |
|
Достаточно обеспеченных азотом почв |
7 |
|
Промежуточный между 7 и 9 |
8 |
|
Богатых азотом почв |
9 |
|
Промежуточный между 9 и 11 |
10 |
|
Избыточно богатых азотом почв |
11 |
Интересна судьба на пробной площади дрока красильного ( Genista tinctoria L.): причину его исчезновения достаточно уверенно можно связать с увеличением богатства почвы азотом за пределы толерантности этого вида. Так, дрок красильный оценивается по шкале богатства почв азотом, как вид, встречающийся в диапазоне условий от 1 (без-азотные почвы) до 5 (бедные азотом почвы), в то время, как показатель богатства почв ПП-10 в 1955 году 5 баллов (бедные азотом почвы) к 1988 году стал равен 7, что соответствует по таблицам Д.Н. Цыганова типу режима «достаточно обеспеченных азотом почв».
Для понимания вклада в динамику стенобионт-ных видов разных жизненных форм (деревьев, кустарников, кустарничков и трав) было определено их число и относительная доля по годам наблюдений. О мхах и лишайниках, встреченных на ПП-10, данные в шкалах Д.Н. Цыганова о толерантности по факторам отсутствуют, поэтому в расчетах они не учитывались. Число видов разных жизненных форм от первого (1937 г.) к последнему сроку наблюдений (1998 год) приводится в таблице 3.
Таблица 3. Число видов разных жизненных форм
|
Число видов |
1937 г. |
1957 г. |
1988 г. |
1998 г. |
|
Всего |
80 |
48 |
37 |
36 |
|
Деревьев |
11 |
6 |
10 |
11 |
|
Кустарников |
8 |
6 |
5 |
4 |
|
Кустарничков |
2 |
1 |
0 |
0 |
|
Трав |
54 |
33 |
22 |
21 |
|
Мхов |
4 |
2 |
0 |
0 |
|
Лишайников |
1 |
0 |
0 |
0 |
По фактору увлажненности почв (рис. 3) доля стенобионтных видов среди деревьев практически постоянна; доля стенобионтов среди кустарников возрастает, доля стенобионтов среди трав можно считать близкой к постоянной.

0 -------9-
ООО
1937 г. 1957 г. 1988 г. 1998 г.
годы наблюдений
—в—деревьев кустарников кустарничков —Ж—трав
Рис. 3. Доля стенобионтных видов разных жизненных форм по фактору увлажненности почв; по вертикальной оси процент видов стенобионтов от числа видов определённой жизненной формы в определённый срок наблюдения; по горизонтальной оси – сроки наблюдений
По фактору богатства почв азотом (рис. 4) сте-нобионтных видов среди деревьев нет; доля стено-бионтов среди кустарников также возрастает, сте-нобионтов среди кустарников и кустарничков к 1988 году в сообществе не остается; среди трав доля стенобионтов близка к постоянной.
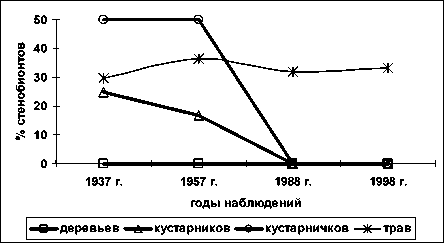
Рис. 4. Доля стенобионтных видов разных жизненных форм по фактору богатства почв азотом; по вертикальной оси процент видов стенобионтов от числа видов определённой жизненной формы в определённый срок наблюдения; по горизонтальной оси – годы наблюдений
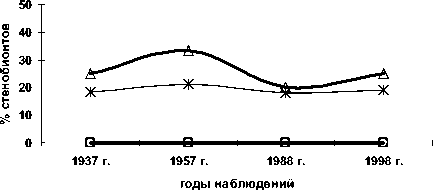
По фактору кислотности почв (рис. 5) стеноби-онтных видов среди деревьев и кустарничков нет; доля стенобионтов среди кустарников и трав, несмотря на некоторые колебания, остается близка к постоянной.
По фактору освещенности-затенения (рис. 6) стенобионтных видов среди деревьев, кустарничков и кустарничков нет; к 1988 году все травы сте-нобионты по фактору освещённости-затенения вы- падают из сообщества, что можно объяснить изменениями освещённости, произошедшими на площади [11].
□ деревьев кустарников о кустарничков -ж-трав
Рис. 5. Доля стенобионтных видов разных жизненных форм по фактору кислотности почв; по вертикальной оси процент видов стенобионтов от числа видов определённой жизненной формы в определённый срок наблюдения; по горизонтальной оси – годы наблюдений
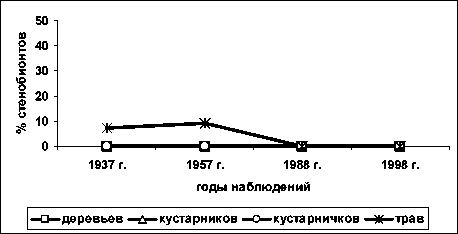
Рис. 6. Доля стенобионтных видов разных жизненных форм по фактору освещённости-затенения; по вертикальной оси процент видов стенобионтов от числа видов определённой жизненной формы в определённый срок наблюдения; по горизонтальной оси – годы наблюдений
Сводный график изменения числа стенобионт-ных видов без разделения на жизненные формы приведён на рис. 7.
Таким образом, несмотря на значительное уменьшение общего числа видов на пробной площади (табл. 3), относительная доля видов, стеноби-онтных по факторам влажности почв и кислотности почв, существенно не изменилась. Вместе с этим по факторам богатства почв и освещённости произошло уменьшение доли стенобионтных видов. Это подтверждает предположение, что при изменениях определённого фактора среды доля растений-стенобионтов по этому фактору уменьшается, а при относительном постоянстве фактора доля стеноби-онтов по фактору остается постоянной или возрастает, что ранее показали наши исследования на больших массивах данных.
ЗАКЛЮЧЕНИЕ
Исследование списков видов, встреченных на постоянной пробной площади, заложенной в 1937 г., растительный покров которой развивался без значительных антропогенных воздействий в течение 61 года, показало увеличение с течением времени богатства почв азотом в результате жизнедея- тельности биоценоза и, как следствие, замещение растений более бедных мест обитания на растения более богатых мест обитания; обнаружено уменьшение доли видов стенобионтов по этому фактору. Отсутствие существенного изменения влажности и кислотности почв пробной площади за рассмотренный период, несмотря на уменьшение числа видов на пробной площади более чем в два раза, не привело к существенному уменьшению доли сте-нобионтов по этим факторам среды.
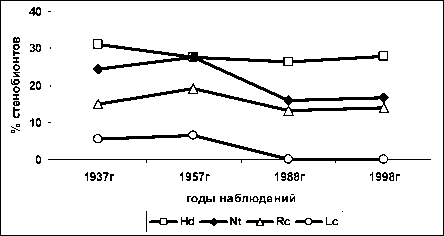
Рис. 7. Суммарная доля стенобионтных видов на пробной площади ПП-10 Воронежского заповедника по шкалам Д.Н.Цыганова: Hd – шкала увлажненности почв, Nt – богатства почв, Rc - кислотности почв, Lc - освещенности-затенения; по горизонтальной оси - годы исследования, по вертикальной – доля участия (%) в списке видов
Выражаю благодарность к.б.н. Е.А. Стародубцевой за предоставленные данные и ценные замечания, проф. А.С. Комарову и к.б.н. Л.Г. Ханиной за консультации и дискуссии по теме и к.б.н. Т.И. Грохлиной за возможность обрабатывать данные программой EcoScaleWin.
Список литературы Долговременная динамика видов лесных растений с различной шириной экологических ниш на постоянной пробной площади в сосняке с дубом и липой в Воронежском заповеднике
- Зубкова Е.В. Изменения соотношения реализованных экологических ниш растений в сообществах при сукцессии//Изв. Самар. НЦ РАН. 2009. Т. 11, № 7(11). С. 1634-1639.
- Комаров А.С., Зубкова Е.В. О стенобионтности и эврибионтности у лесных растений//Современные проблемы популяционной экологии, геоботаники, систематики, флористики: материалы международной научной конференции, посвященной 110-летию А.А. Уранова. Т. 1. Кострома, КГУ им. Н.А.Некрасова, 2011. С. 334-339.
- Грохлина Т.И., Ханина Л.Г. Автоматизация обработки геоботанических описаний по экологическим шкалам//Принципы и способы сохранения биоразнообразия. Материалы II Всероссийской научной конференции. Йошкар-Ола: Мар. гос. ун-т. 2006. С. 87-89.
- Зубкова Е.В., Ханина Л.Г., Грохлина Т.И., Дорогова Ю.А Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoScaleWin:Учебное пособие/Мар. гос. ун-т, Пущинский гос. ун-т. Йошкар-Ола: МарГУ, 2008. 96 с.
- Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука. 1983. 196 с.
- Ellenberg H. Zeigerwerte von Pflanzen in Mitteleuropa [Indicator values of plants in Central Europe]/H. Ellenberg, H.E. Weber, R. Dull, V/Wirth, W. Werner, D. Paulisen//Scripta Geobotanics/V.18. Verlag Erich Goltze KG, Göttin-gen, 1991. 248 s.
- Бигон М., Харпер Дж., Таунсенд К. Экология. Особи, популяции и сообщества: В 2-х т. Т. 1. М.: Мир, 1989. 667 с.
- Зубкова Е.В. О некоторых особенностях диапазонных экологических шкал растений Д.Н.Цыганова//Изв. Самар. НЦ РАН. 2011. Т. 13, № 5. С. 48-53.
- Заугольнова Л.Б., Морозова О.В. Группы типов леса южной тайги и подтаежной зоны Европейской России//Ценофонд лесов Европейской России [Электронный ресурс] URL: http://mfd.cepl.rssi.ru/flora/type_forest3. html.
- Травникова Л.С. Влияние поселения липы под пологом сосняка дубнякового на свойства почвы//Труды Воронежского госзаповедника. Вып. VIII. Воронеж, 1959.С. 264-267.
- Стародубцева Е.А., Лихацкий Ю.П., Трегубов О.В. Динамика лесного покрова на песчаных террасах Воронежского биосферного заповедника//Восточноевропейские леса: история в голоцене и современность. Под. ред. О. В. Смирновой. Кн. 2. М.: Наука, 2004. С. 200-235.
- Губанов И.А., Киселёва К.В., Новиков В.С., Тихомиров В.Н. Иллюстрированный определитель растений Средней России. М.: МКМ, Том 1, 2002. Том 2, 2003. Том 3, 2004.
