Древнейшие фоссилизированные микроорганизмы Фенноскандинавского щита

Автор: Светов С.А., Медведев П.В.
Журнал: Вестник геонаук @vestnik-geo
Статья в выпуске: 4 (208), 2012 года.
Бесплатный доступ
Впервые в истории палеонтологических исследований на Фенноскандинавском щите обнаружены древнейшие фоссилизированные микроорганизмы в кремнистых осадочных породах (хемогенных силицитах), приуроченных к нижней части разреза мезоархейской (3.0—2.9 млрд лет) протоокеанической ассоциации Ведлозерско-Сегозерского зеленокаменного пояса (Центральная Карелия). Выявленные микрофоссилии представлены окремнёнными фрагментами чехлов нитчатых микроорганизмов, обитавших в морских глубоководных условиях вблизи гидротермальных систем.
Фенноскандинавский щит, мезоархей, микрофосcилии, ведлозерско-сегозерский зеленокаменный пояс
Короткий адрес: https://sciup.org/149129070
IDR: 149129070
Oldest fossilized microorganisms of Fennoscandian shield
Oldest fossilized microorganisms in chemically precipitated siliceous rocks with age ca. 3 Ga were discovered on Fennoscandian shield for the first time. The rocks are situated in the low part of oceanic volcanic succession in Vedlozero-Segozero greenstone belt (central Karelia). Microfossils which were found there are interpretated as silicified fragments of filamental microorganisms inhabitated zone of hydrothermal activity in a deep water environment.
Текст научной статьи Древнейшие фоссилизированные микроорганизмы Фенноскандинавского щита
Обнаружение остатков ископаемых бактерий в палеоархейских, начиная с 3.5 млрд лет, зеленокаменных комплексах Земли является уникальным достижением современной геологии. Древнейшие на сегодняшний день остатки микробиоты обнаружены в кремнистых породах, метаморфизованных в зеленосланцевой фации из вулканогенных разрезов зеленокаменных поясов Западной Австралии (серия Варравуна, 3.49 — 3.31 млрд лет) и Южной Африки (серии Онвервахт, 3.50—3.26 млрд лет и Фиг Три, 3.26—3.23 млрд лет). Подробнее об этих находках и об обосновании возраста вмещающих пород можно узнать в обзоре [11] и атласе [1]. Следует отметить, что все найденные древнейшие микрофоссилии связаны с микробиальными постройками (строматолитами и др.). Большинство специалистов трактует палеоар-хейские микрофоссилии как остатки коккоидных и нитчатых бактерий, обладавших хемолитотрофным метаболизмом и не нуждавшихся в солнечном свете и кислороде. За последние 15 лет многочисленные минеральные псевдоморфозы по микроорганизмам были обнаружены в метеоритах, относящихся к классу углистых хондритов [1, 14]. Несмотря на продолжающие ся дебаты по обоснованию биологической природы древнейших микро-фоссилий [1, 4, 12], эти находки позволяют существенно расширить временные рамки, в которых обнаружены следы архейской жизни.
На территории Фенносканди-навского щита самыми древними находками являются сфероморфные акритархи и фрагменты нитчатых микроорганизмов, выделенные методом мацерации (химического растворения кремнезёма плавиковой кислотой) из метаосадочных пород мезоархейско-го возраста (2.85—2.86 млрд лет) районов р. Семчь и оз. Совдозеро в Центральной Карелии [12]. В настоящее время эти находки подтверждены данными электронной микроскопии и дополнены новыми разновидностями микрофоссилий, обнаруженными в осадочных породах возрастом 2.80— 2.70 млрд лет в Хизоварской структуре (Парандово-Тикшозерский зеленокаменный пояс) на севере Карелии [1].
В этой работе приводятся результаты палеонтологического изучения древнейших (3.0—2.9 млрд лет) на Фенноскандинавском щите осадочных породных ассоциаций Ведлозер-скоегозерского зеленокаменного пояса (Хаутаваарская структура), Цент ральной Карелии, представленных хемогенными силицитами (рис. 1, а).
Мезоархейский Ведлозерско-Сегозерский зеленокаменный пояс представляет собой сохранившийся реликт аккреционной системы, сложенный несколькими стратотектоническими ассоциациями, образованными в контрастных геодинамиче-ских обстановках, сходных с современными конвергентными зонами перехода океан-континент [7]. Древнейшие комплексы зеленокаменного пояса представлены вулканитами ко-матиит-базальтового состава с прослоями глубоководных кремнистых и углеродистых осадков [10], интерпретируемыми как фрагмент океанического плато, а также базальт-андезит-дацитовой ассоциацией, определяемой как островодужный комплекс [6].
Мафитовый комплекс Хаутаваар-ской структуры в современном эрозионном срезе выделен стратифицированной лавово-пирокластической толщей высокомагнезиальных пород ко-матиит-базальтовой ассоциации мощностью около 2.5 км [8]. Осадочные породы — силициты, туффиты, графити-стые алевролиты широко распространены в разрезе в виде цемента в подушечных лавах коматиитов, базальтов и отдельных маломощных (до 3 м) про-

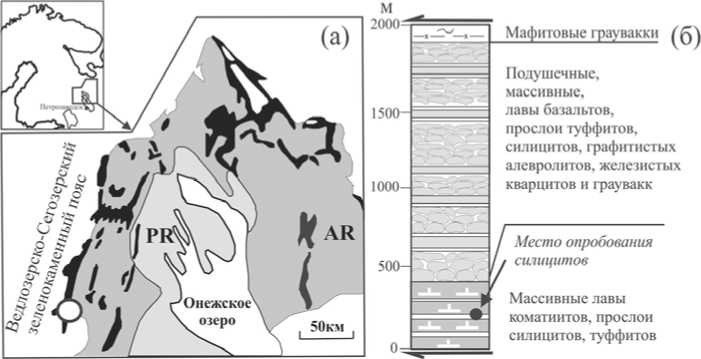
Рис. 1. Упрощенная схема геологического строения палеоархейского Ведлозерского блока и положения зеленокаменных структур в его обрамлении: а черным цветом показаны мезоархейские зеленокаменные структуры, темно-серым архейские комплексы Ведлозерского блока, светло-серым перекрывающие палеопротерозойские породные ассоциации. Район работ (Хаутаваарская структура) обозначен кругом; б — стратиграфическая схема мезоархейской коматиит-базальтовой ассоциации Хаута-ваарской структуры (от остального разреза отделена тектоническими зонами)
слоев туффитов и силицитов между массивными лавовыми потоками (рис. 1, б). Время формирования коматиит-базальтовой ассоциации по данным SmNd метода (по породе в целом) составляет 2921 ± 55 млн лет [7].
Изучаемые нами мезоархейские силициты представляют собой афанитовые кремнистые и алюмокремнис-тые породы хемогенной природы, маркирующие в разрезе мафитовой ассоциации перерывы в вулканической деятельности [8]. Силициты характеризуются массивной, тонкослоистой, реже конкреционной текстурой, имеют серый, зеленовато-серый, розоватый или черный цвет, обусловленный присутствием в осадочных породах хлорита, окислов железа и тонкодисперсного графитистого вещества. В минеральном составе силицитов преобладает волокнистый халцедон (частично перекристаллизованный в ходе регионального метаморфизма в кварц), слагающий породу на 75—80 %. В виде примесей присутствуют серицит, альбит, калиевый полевой шпат, биотит, мусковит. В качестве акцессорных минералов выявлено небольшое количество зерен терригенного циркона (чаще осколков, имеющих слабо окатанные, корродированные формы), а также редкие минеральные агрегаты ниобатов группы фергюсонита-самарскита. Исследуемые породы метаморфизованы в условиях хлорит-серицитовой субфации зеленосланцевой фации регионального метаморфизма [3].
Химический состав 15 проб силицитов определен методом «мокрой»
химии в Институте геологии КарНЦ РАН (г. Петрозаводск). Содержание петрогенных элементов (мас. %) в кремнистых породах варьируются в следующих пределах: SiO2 — 75.20—86.23, TiO2 — 0.03—0.28, Al2O3 — 8.10—11.41, Fe2O3 — 0.14—1.52, FeO — 0.10—0.36, MnO — 0.01—0.05, MgO — 0.21—0.57, СаO — 0.14—0.98, Na 2 O — 0.50—2.10, K2O — 3.15—4.47, H2O — 0.06—0.22, п.п.п 0.30—1.72.
Оценка времени формирования осадочных прослоев и соответственно коматиитового разреза проведена на основе прецизионного изучения цирконов из слабоизмененного слоя силицитов (залегающего в пачке с тонким переслаиванием мафитовых туффитов и силицитов) в средней части коматиитового разреза Хаутаваар-ской структуры (рис. 1, б). Из слоя силицитов была отобрана проба весом 10 кг, из которой удалось выделить около 20 зерен циркона, имеющих торпедовидный, копьевидный, гиацинтовый и изометричный облик. Цвет зерен варьируется от розоватого, желтоватого до сероватого. Размер зерен колеблется от 0.35 х 0.10 до 0.15 х 0.11 мм. Микрозондовое исследование циркона показало наличие в них микровключений ильменита, апатита, олигоклаза и фрагментарно сохраненное обрастание циркона каймами плагиоклаза (лабрадор-андезин). Для большинства зерен характерна магматическая зональность и хорошая сохранность ядерных частей кристаллов. Именно такие кристаллы были изучены на прецизионном вторичноионном микрозонде высокого разре шения SHRIMP-II в Центре изотопных исследований ВСЕГЕИ. В результате датирования шести зерен цирконов установлено, что Th/U-отноше-ние для данных зерен варьируется в пределах 0.42—1.32, их изохронный возраст составляет 2917.2 ± 8.7 млн лет (рис. 2). Таким образом, полученные временные значения могут рассматриваться как возможное древнее ограничение времени формирования высокомагнезиальной ассоциации Хаутаваарской структуры, что хорошо согласуется с другими геологическими и геохронологическими данными [7, 9].
Для палеонтологического исследования был выполнен отбор проб из тех же выходов силицитов, по которым ранее осуществлялось геохронологическое датирование (рис. 1, б). В работе использованы штуфы породы, не имеющие контакта с дневной поверхностью, без корок выветривания, трещиноватости и любых проявлений вторичных процессов (ожелезнения, карбонатизации и т. д.). Глубина отбора проб от дневной поверхности варьировалась от 15 до 20 см. Электронно-микроскопически анализировались сколы силицитов площадью от 2 до 4 кв. см, приготовленные непосредственно перед исследованием. В стерильных лабораторных условиях образцы со свежими сколами промывались последовательно в воде (15 минут с ультразвуковой обработкой) и позже в спирте для исключения поверхностного загрязнения. В общей сложности было подготовлено 10 проб с различными плоскостными сечениями (как вкрест напластованию, так и параллельно).
Исследования образцов проводились на сканирующем электронном микроскопе VEGA IILSH (Tescan) с энергодисперсионным микроанализатором INCA Energy 350 (Oxford instruments), с использованием BSE и SE детекторов при HV=20 кВ в Институте геологии КарНЦ РАН (г. Петрозаводск).
В результате электронно-микроскопического исследования было установлено присутствие в мезоархейских кремнистых осадочных породах нескольких разновидностей фоссилизи-рованных микроорганизмов (рис. 3, а—е). Наибольшее количество находок связано с обнаружением фрагментов нитевидных трубчатых микрофосси-лий длиной от 30 до 600 мкм, диаметром от 5 до 20 мкм и толщиной стенок 0.5—1 мкм. Кроме того, выявлены
0.66
0.62 ■■
Точки: 3.1,4.1, 5.1, 6.1, 7.1, 8.1 Пересечения:
СКВО = 0.15
0.58
0.54
0.50
2600,
0.46
14 16
2°7рь/235и
3_1 (2905+20 млн лет)
4_1 (2929±29 млн лет)
5_1 (2907+7 млн лет)
7 1 (2917±7 млн лет) .
- 8_1 (2919±9 млн лет)
6_1 (2934±12 млн лет)
Рис. 2. Диаграмма цирконов с конкордией из слоя силицитов (залегающего в пачке с тонким переслаиванием мафитовых туффитов и силицитов) в средней части коматиитового разрезаХаутаваарской структуры. Морфология зерен цирконов и положение точек датирования на вторично-ионном микрозонде высокого разрешения SHRIMP-II. В скобках, рядом с номером точек, приведены значения 207Pb/206Pb возраста цирконов
многочисленные деформированные (скрученные, сплющенные, разорванные) фрагменты отдельных чехлов (рис. 3, д—е). В единичных случаях обнаружены нижние части колоний нитевидных трубчатых микрофоссилий (собранных в пучки), представляющих в поперечном сечении округлые агрегаты размером до 60 х 100 мкм, крепящиеся к породе пленкообразным бактериальным агрегатом (рис. 3, г). Вероятнее всего, данные находки являются фоссилизированными (замещёнными кремнезёмом) чехлами хе-молитотрофных микроорганизмов (рис. 3, ав), обитавших в глубоководных обстановках в зоне гидротермальной активности, связанной с ультраосновным (коматиит-базальтовым) подводным вулканизмом.
Выявленные в мезоархейских сили-цитах биоформы морфологически подобны нитевидным микроорганиз
100 мкм мам, установленным в корах выветривания палеопротерозойского (~2.4 млрд лет, район оз. Паанаярви, Северная Карелия) и архейского возрастов (~2.8 млрд лет, район оз. Воронье, Лехтинская структура, Северная Карелия), а также бактериальным формам, найденным в углистом хондритовом метеорите Мурчисон (CM2) [1].
Геологические данные, полученные в результате предыдущих исследований Ведлозерско-Сегозерского зеленокаменного пояса [510] позволяют нам провести интерпретацию условий формирования кремнистых породных серий. На основе детального петрологического и изотопно-геохимического изучения коматиит-ба-зальтовой ассоциации Хаутаваарской структуры (содержащей прослои силицитов) установлена принадлежность коматиитов к Al-недеплетиро-ванному типу Мунро (аналогичных комплексу Мунро Тауншип, Канада [13]), формировавшихся в плюмовом режиме [7,13]. Широкое распространение в ассоциации с коматиитами мощных (1500—1800 м) толщ толеитовых подушечных лав, содержащих между потоками прослои туфов и глубоководных осадочных пород — силицитов, графитистых алевролитов с горизонтами железистых кварцитов, позволяет предполагать образование коматиит-базальтового комплекса в океанических обстановках внеосевого вулканизма (удаленного от спре-динговых центров), подобных современным океаническим плато [7, 10].
Формирование силицитов, происходившее в перерывах между фазами вулканической активности, было обусловлено химическим осаждением кремнезема из метастабильных коллоидных растворов, связанных с деятельностью гидротермальных систем в придонных областях.
Вполне вероятно, что флюидные растворы глубинной циркуляции в архейских бассейнах, подобно современным морским гидротермальным системам, характеризовались повышенной температурой и, наряду с кремнеземом, были обогащены СО2 и СН4 [2], делая среду потенциально благоприятной для развития термофильных хемо-литотрофных микроорганизмов.
В результате исследования древнейших силицитов Фенноскандинав-ского щита впервые установлено, что на данной территории 3.0—2.9 млрд лет назад существовали специфические сообщества микроорганизмов, представленные трубчатыми нитевидными бактериальными формами, имеющими органическую оболочку и центральную полость. Размеры построек достигали 100 мкм в диаметре при высоте нитевидных отростков 200—600 мкм. Важно подчеркнуть, что основные оазисы жизни были локализованы в зонах гидротермальной активности, связанной, в данном случае, с подводным коматиитовым вулканизмом. В этих условиях развивались хемолитотрофные микроорганизмы, для метаболизма которых не нужны ни солнечный свет, ни кислород, поскольку все необходимые для хемосинтеза компоненты поставлялись с продуктами поствулканической деятельности.
Исследования ведутся в рамках программы фундаментальных исследований № 28 Президиума РАН «Проблемы происхождения жизни и становления биосферы», проект «Реконструкция

20мкм
1 '20 м км
Рис. 3. Электронно-микроскопические изображения древнейших мезоархейских (3.0— 2.9 млрд лет) фоссилизированных микроорганизмов, выявленных в хемогенных си-лицитах Хаутаваарской структуры Ведлозерско-Сегозерского зеленокаменного пояса: а — длинный окремнённый чехол нитчатого микроорганизма; б — увеличенная деталь стенки чехла с обрастанием мелкими гранулами кремнезёма, образовавшимися на стенке лопнувшего чехла в процессе фоссилизации; в — эффект скручивания трубчатого чехла микроорганизма; г — нижняя часть предполагаемой колоний нитевидных трубчатых микрофосилий, представляющая в поперечном сечении округлые агрегаты, крепящиеся к породе пленкообразным бактериальным агрегатом; деформированные чехлы микроорганизмов: д — скрученный и уплощенный; е — с разорванными стенками на обоих концах трубки
древнейших (3.8—3.0млрд лет) условий становления и последующего развития микробиальных сообществ раннего докембрия на примере восточной части Фенноскандинавского щита» и поддержаны грантом РФФИ 11-05-93959-ЮАР_а.
Список литературы Древнейшие фоссилизированные микроорганизмы Фенноскандинавского щита
- Астафьева М. М., Герасименко Л. М., Гептнер А. Р. Ископаемые бактерии и другие микроорганизмы в земных породах и астроматериалах. М.: ПИН РАН, 2011. 172 с.
- Короновский Н. В. Гидротермальные образования в океанах//Соросовский образовательный журнал, 1999. № 10. С. 55-62.
- Металлогеническая эволюция архейских зеленокаменных поясов Карелии. Ч. I./Отв. ред. Ю. И. Лазарев. СПб.: Наука, 1993. 208 с.
- Юшкин Н.П., Асхабов А.М., Анищенко Л.А., Безносова Т.М., Безносов П.А., Бушнев Д.А., Голубев Е.А., Каткова В.И., Камашев Д.В., Ковалева О.В., Лукин В.Ю., Лысюк Г.Н., Лютоев В.П., Ракин В.И., Соболев Д.Б., Тельнова О.П., Цыганко В.С., Шанина С.Н. Происхождение биосферы и коэволюция минерального и биологического миров. Сыктывкар: ИГ Коми НЦ УрО РАН, 2007. 202 с.
- Слабунов А. И., Лобач-Жученко С. Б., Бибикова Е. В. и др. Архей Балтийского щита: геология, геохронология, геодинамические обстановки//Геотектоника, 2006. № 6. С. 1-29.
