Effect of salinity stress on chlorophyll content, proline, water soluble carbohydrate, germination, growth and dry weight of three seedling barley (Hordeum vulgare L.) cultivars
 cultivars")
Author: Movafegh Somayeh, Razeghi Jadid Roghie, Kiabi Shadi
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 4 т.8, 2012.
Free access
Salinity is a serious environmental constraint to crop production in many parts of the world and the development of crops with improved salt tolerance is proposed as part of solution to this problem. This research was performed out in order to study the effects of different salinity levels on germination, growth, dry weight, proline, water soluble carbohydrate and chlorophyll content of three barley (Hordeum vulgare L.) cultivars named Jonoob (INC-54), Reyhan (INC-45) & Nosrat (INC -47).The experiment was carried out using factorial based on completely randomized design with three replications. Seven old seedlings after germination were transferred to Hoagland nutrient solution under the effect of salinity levels (0, 50, 150 and 250 mM NaCl)in during seven days. Data variance analysis showed that seed germination of three barley cultivars was significantly (PReyhan>Nosrat. The results showed that, increasing in salinity decreased all growth parameters. Salinity stress decreased shoot and root length, root dry weight and chlorophyll contents in every three cultivars. But decreasing of chlorophyll was less in Nosrat compared to two other items. Proline content and soluble carbohydrate were increased in all of the three cultivars with enhance of NaCl concentration. By increasing of salinity stress accumulation of proline and soluble sugar content in leaves of Nosrat cultivar was more than other cultivars. As saltiness increases resistance natural responses in this plant gets better considering less decrease in chlorophyll amount and strategy of more production about praline and sugar solution compared to two other items.
Barley, chlorophyll, dry weight, germination, proline, salinity, soluble carbohydrate
Short address: https://sciup.org/14323682
IDR: 14323682
Text of the scientific article Effect of salinity stress on chlorophyll content, proline, water soluble carbohydrate, germination, growth and dry weight of three seedling barley (Hordeum vulgare L.) cultivars
The soil salinity affects large areas of the world’s cultivated land, causing significant reductions in crop yield (Grewal, 2010). The soil salinity is one of the most problem affecting agriculture productions fertility in the world an estimated 6% of total global land area, which equates to between 400 and 950 × 106 hectares and nearly half of all irrigated land being adversely affected by high salinity. High satl stress disrupts the homeostasis of water potential and ion distribution at both the cellular and the whole-plant levels (Wenxue, 2003).
Barely ( Hordeum vulgare L.), is one of the most important cereal production in all over the world which it has main role in human and herd feeding (Walia, 2006). This plant with scientific name of Hordeum vulgare is from Gramineae (Poaceae) family and Hordeum specie. From under cultivating level, barely is the fourth cultivated cereal in the world which this title due to it's vast consistency in arid and semi- arid situation and water tensions and salinity (Gill 1999).
The germination phase is the most important and sensitive phases in salinity tension (Dizaji, 1998). Salinity by reducing of water potential and special poisonous ions such as: Na+, Cl- and reducing of its necessary nutrition calcium and potassium has negative influence on seed germination (Chesson, 2004). Salinity stress increase ions poisonous, changing enzymes activation and seeds hormones in germination phases (Othman, 2006). Salinity by competing in nutrition element absorption causes to growth reduction. Different experiments on cultivating plants explained this subject which increasing of salinity can reduce micro- stem and micro- root length and also dry weight of these bodies in comparison with instance sample meaningfully (Chesson, 2004). Plants have many behaviors against with salinity indicating. Osmoregulation in plants is one of the effective and meaningful mechanisms against with salinity stress (Paneknyat, 2007). The most important of these osmoregulators solutions including of soluble sugars (such as: sakaroz, glucose, fructose, terhaloz, rafinoz (Prabhjot, 2001). Insoluble sugar (starch, amylose, amilopectin,) and Tetra amino acid such as proline (Shuji, 2002). Proline and hydroxyproline in structural proteins are clearly distinguished from free proline, which serves to regulate osmotic adjustment. proline transporter (HvProT) was highly expressed in the apical region of barley roots under salt stress (Ueda, 2007). Increasing of aminoacid concentration which contributes to osmoregulation, is due to several factors such as: inhibition of proline analysis, inhibition of enter of proline to protein or increasing of protein analysis which accompany with growth reduction (Kao, 1981).
Proline has conservative effect on nitrate reductase activation of cellular membrane in barely plant leaf under low- water situation seems that cellular membrane resistance is one of the best physiologic indices against with environmental stress and is as a parameter for plants selection with high resistance is high (Neto, 2004 , Tantau, 2004).Carbohydrate soluble aggregations in leaves under stress indicating of it's involve in plant consistency process (Kerepesi, 2000).The reduction of photosynthesis activity is one of the main reasons in salinity influence of plants in which is due to chlorophyll reduction and also reduction of Co 2 absorption and photosynthesis capacity (Fransisco, 2002).
MATERIALS AND METHODS
For studying of salinity stress effect on germination and physiological factors of Barley ( Hordeum vulgare L.) seeds were prepared in Karaj plant Institute, Iran. Related study was carried out in 2010 at Islamic Azad University of Tonekabon under controlled condition. This experiment was carried out using factorial based on completely randomized design with three replications. For studying of salinity tension effects on germination phase seeds was put in 5% sodium hypochlorite solution of for 20 min and then by distilled water washed for antiseptic of it. Seeds was cultivated in Petri on wet filter paper soaked with the four levels of salinity include 0, 150, 50,250 m/mol chloride sodium. In every dish putted 30 seeds. Then plates were putted in 20 centigrade in incubator(250C, 12 h of light and 12 h of dark), after 3 days germinated seeds enumerated. For evaluation of physiological parameters , Seven old seedlings after germination on wet filter paper soaked with distilled water in incubator, were transferred to Hoagland nutrient solution under the effect of salinity levels (0, 50, 150 and 250 mM NaCl) in during seven days. The Hoagland solution was continuously aerated changed twice a week (table 1). PH was 6-6.5. After passing of 3 days, salinity care, 0, 50, 150, 250 m/mol, NaCl imposed on them for one week. Plants were grow under controlled situation (16 hours light on 250C, 8 hours darkness on 200C and environment moisture 60-70%) (Yousfi, 2007). Then physiological parameters were evaluated. After passing of stress period, proline was evaluated by Bates method
(1973), sugar by Nelson method (1943), chlorophyll by Lichtenthaler method (1987). Other parameters included measurement of root/shoot dry weight, root height and aerial or atmospheric body. The dry matter was determined by drying the plants in an oven at 65-70 0C for 48 hours to a constant weight. Data Analysis, variance and comparison of mean by Duncan experiment and SPSS software was done and drawing of diagrams was used from Excel software.
RESULTS
Salinity and germination : statistical analysis of studied behaviors showed that by increasing of salinity, germination percent will reduce.
Based on the table of comparison, the highest percent of germination belongs to Jonoob type with 98.3% in control level and 75% in 250m/mol. The least percent of germination belonging to Nosrat type with 93.3% in control level and 40% in 250m/mol level.
Salinity and growth: variance analysis table shows that the length of atmospheric body and root and root/shoot dry weight in different salinity levels and in comparison with three types of them, they have meaningful difference together.(P<0/01) salty condition had less effect on Nosrat's air organs height and dry weight compared to other items and the lengths of Reyhan's and Jonoob's stem under salty condition was significantly decreased (table 4). Considering to the table of comparison, shortest height of air organ belongs to 250 mgr/lit salt and the highest belongs to control level. This matter applies to all three items under study. (Table 3) Decrease of dry weight in shoot and root are noticeable respectively compared to highest level of saltiness (250 mgr/lit salt) and results show that dry weight of three kinds of barely under study was dramatically decreased under salty condition. (Table 2 and 3)
Salinity and carbohydrate solution: Based on a variance analysis table, carbohydrate solution in atmospheric phase in different levels of salinity in comparison with three types and also interacting effect of types and salinity, had meaningful difference ( p ≤0.01) sugar solution of stem with increasing of salinity increased (fig1).
Salinity and proline rate : Result showed that in every level of salinity and in comparison with three types of barely (Jonoob, Nosrat, Reyhan) it was meaningful difference ( p ≤0.01) saltiness had a considering effect on increase in root's praline in
150 and 250mmol compared to control level and 50 mmol NaCl level. Salinity induced increase in content of leaves and root's proline in Nosrat (0/455a) (0/427a) compared to two other items.
Salinity and chlorophyll: Variance analysis indicates that saltiness effect on cultivars chlorophyll content is meaningful. Table 3 indicates that chlorophyll content of a, b and a+b had noticeable decrease by increasing of salinity in all cultivars. The highest rate of chlorophyll was observed in nutrient solution without salt and in high rates of saltiness (150 and 250 mM) the highest rate of chlorophyll decrease was observed. Considering diagram, chlorophyll was less in Nosrat compared to two other items (fig. 2).
Table 1. Contents of nutrient solution used for irrigation of barely seedlings
|
Element name |
Resource |
Main solution(g/L) |
Suitable content for 1 litere of solution |
|
Potassium di hydrogen phosphate |
KH 2 PO 4 |
34.025 |
1mg/L |
|
Potassium nitrate |
KNO 3 |
25.275 |
5mg/L |
|
Calcium nitrate |
Ca(NO 3 ) 2 .4H 2 O |
59.025 |
5mg/L |
|
Magnesium sulfate |
MgSO 4 .7H 2 O |
61.6 |
2mg/L |
|
Boric acid |
H 3 BO 3 |
0.715 |
1mg/L |
|
Manganese chloride |
MnCl 2 .4H 2 O |
0.4525 |
|
|
Zinc sulfate |
ZnSO 4 .5H 2 O |
0.055 |
|
|
Copper sulfate |
CuSO 4 .5H 2 O |
0.02 |
|
|
Molybdic acid |
H 2 MoO 4 .H 2 O |
0.0225 |
|
|
Coordination complex |
Na 2 EDTA |
0.5 |
1mg/L |
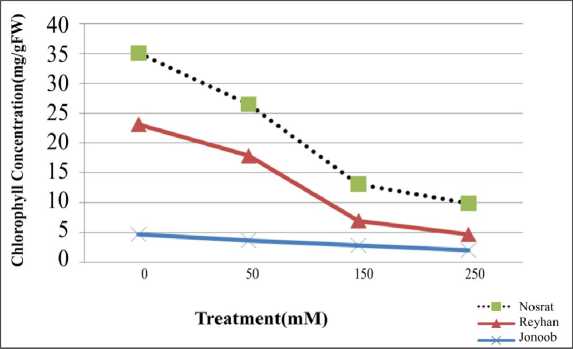
Fig2.Infraction effect of salinity and cultivar on total chlorophyll
Table 2 . Varience analysis of studied parameters
Table 3. Comparsion of cultivars
|
cultivar |
Jonoob (1) |
Reyhan (2) |
Nosrat (3) |
|
Shoot height (cm) |
16.8250 b |
14.900 c |
18.733 a |
|
Root height (cm) |
7.8417 ab |
7.3417 b |
9.2750 a |
|
Shoot dry weight (mg/gr) |
0.0150000 b |
0.0139167 b |
0.0175833 a |
|
root dry weight (mg/gr) |
0.0055833 b |
0.0054167 b |
0.0070000 a |
|
chlorophyll a (mg/g) |
0.18942 b |
0.50867 a |
0.51900 a |
|
chlorophyll b (mg/g) |
0.11458 c |
0.33167 a |
0.23975 b |
|
Chlorophyll (a+b) (mg/g) |
3.3831 c |
9.8269 a |
8.0009 b |
|
leaves sugar (mg/g) |
0.20067 b |
0.23958 b |
0.44150 a |
|
Root sugar (mg/g) |
0.10775 a |
0.16900 a |
0.21550 a |
|
leaves proline(mM/g) |
0.21675 b |
0.19692 b |
0.45567 a |
|
Root proline (mM/g) |
0.27900 b |
0.15392 c |
0.42792 a |
|
Germination persentage |
86.917 a |
75.833 b |
67.083 b |
|
salinity |
Control (1) |
50m M (2) NaCl |
150mMNaCl (3) |
250mMNaCl (4) |
|
Shoot height (cm) |
21.8556 a |
17.7667 b |
15.1222 c |
12.5333 d |
|
Root height (cm) |
11.4444 a |
9.0000 b |
6.8667 bc |
5.3000 c |
|
Shoot dry weight (mg/gr) |
0.0224444 a |
0.0175556 b |
0.0134444 c |
0.008556 d |
|
root dry weight (mg/gr) |
0.0087778 a |
0.0071111 b |
0.0050000 c |
0.0031111 d |
|
chlorophyll a (mg/g) |
0.67111 a |
0.50833 b |
0.25733 c |
0.18600 c |
|
chlorophyll b (mg/g) |
0.38600 a |
0.28578 b |
0.14178 c |
0.10111 c |
|
Chlorophyll (a+b) (mg/g) |
11.7019 a |
8.8553 b |
4.3980 c |
3.3259 c |
|
leaves sugar (mg/g) |
0.08800 d |
0.17944 c |
0.29511 b |
0.61311 a |
|
Root sugar (mg/g) |
0.06267 b |
0.07156 b |
0.22378 ab |
0.29833 a |
|
leaves proline(mM/g) |
0.12533 c |
0.20222 c |
0.33067 b |
0.50089 a |
|
Root proline (mM/g) |
0.11344 b |
0.20633 b |
0.38644 a |
0.44156 a |
|
Germination persentage |
95.556 a |
84.222 a |
69.444 b |
57.222 b |
DISCUSSION different levels of salinity on growth and dry
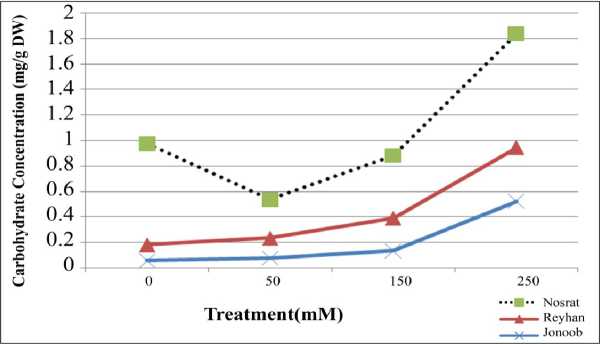
Fig Un traction effect of salinity and cultivar on carbohydrate of leaves
Salinity on germination and growth parameters: Different studies showed that the increasing of salinity had meaningful reduction on germination percentage in variety of plants such as Panicum miliaceaum (Alizadeh et al, 2007), Hordeum vulgare L. (Youzung, 2006), Elymus junceus (Askarian, 2005), Triticum aestivum (Al-Ansari, 2003). Under saltiness tension low moisture would lead to stop of metabolism while germination and changes seeds harmonic activities (Othman, 2006). Salinity due to solution's osmotic potential reduction, toxic ions production and imbalance in nutritional elements, causes decrease of germination..Sodium Chloride has a direct negative effect on embryo's growth and finally leads to decrease in germination percentage and delay in appearance of roots in seeds. Researchers found out that extension of embryo's lengths is highly stopped by high rates of salt (Alizadeh et al, 2007). Salinity due to osmotic reduces water absorption and due to aggregation of ions such as Na+ and Cl- can lead to imbalance in nutrition and toxicity (Shokohifard, 1989).
Root as a filter controls ions passing and prepares convenience proportion of Na+ and K+ ions for plant activity, so root consumes much energy against with tension and this matter can reduce the applicability of root and also absorption of nutrition elements for atmospheric bodies and as a result, the root and atmospheric body dry weight reduce. Disordering in absorption system and transferring of material under environmental stress by preparing of inconvenience proportion K/Na has negative effect on physiological process and it creates toxicity. In a such situation, plant by some mechanisms such as ions leakage to out of root, absorption by woody vessel cells can maintain salt in a low grade in cytoplasm and this reason can be caused either reducing of root length or becoming corked (Safarnejad, 2007).
Salinity on soluble carbohydrate and proline: Plants develop different mechanisms in order to face environmental tensions. Gathering of osmoregulatores is one of the plant's first responses to saltiness tension. Such as alcoholic sugars (glycerol, inozytol) and simple sugars (froktoz, glucose), mixed sugars (Terholoz, Rafinoz, froktans) and proline. (Shuji, 2002). In many studies there has been an emphasis on accumulation of polyamines and carbohydrates as an effective mechanism in different plants under salinity stress which can infer to meaningful increasing on Oriza sativa L. (Amirjani, 2010), Sorghum bicolor L. (Faheed, 2005), Sesbania grandiflora (Dhanapackiam et al, 2010), portulaca oleracea L. (Yazici, 2007) , Lycopersicon esculentum (Amini, 2005) , Vicica faba (Neto, 2004).
Gathering these osmoregulatores in leafs during environmental tensions including saltiness indicates their role in adjustable process. Proline is one of the well-known osmoprotectants and its accumulation is widely observed in various organisms under salt stress. Proline accumulation is regulated by multiple factors, such as its synthesis, catabolism, utilization for protein synthesis and transport from other tissues. Proline gathering in environmental tension such as saltiness could be due to denovo synthesis which reason of proline biosynthesis in salty stress is because of increase in level of 1-D– Piroline – 5 – carboxylate synthesis enzyme
(Grewal, 2010). Different roles has been presented related to proline gathering in plant tissues in environmental stress that could lead to adjust osmosis, integration of plasma membrane, energy source, carbon and nitrogen source, destroyer of free radicals of hydroxyl and creation of structural proteins (Dhanapackiam, 2010). Ueda et al (2007) demonstrated that proline transporter (HvProT) was highly expressed in the apical region of barley roots under salt stress. Salt stress increased proline and hydroxyproline contents in the cell wall fraction of the root apical region, suggesting increment of proline utilization. Expression of the genes encoding cell wall proteins (proline rich protein and extensin) and cellulose synthase was induced in barley roots by salt stress. These findings indicated that free proline transported by HvProT presumably behaved as a component of cell wall synthesis in the apical region of barley roots under salt stress.
Sugars are important molecules that influence on physiological response, regulator genes in metabolism, photosynthesis and defensive reactions. Sugars regulate expression of some genes such as genes coding of amylum constructor proteins, sacarosis metabolism and defensive genes (Amini, 2005). Therefore it is possible that increase in sugar solution of Nosrat cultivar was stimulated by osmotic regulation and in result caused some resistant genes to express. Another thesis in relation to carbohydrate gathering under saltiness is their role in cleaning types of active oxygen (free radical) which are produced by tension. Generally increase of solution sugars during tension process could be related to amylum analysis, sugar synthesis in a non-photosynthesis pathway, not transferring these combinations and decrease in their transaction by leaves and other parts (Ueda et al, 2007).
Salinity on chlorophyll: Under salinity, photosynthesis in leaves surface level decreases due to closing leaf's ostium, decrease in carbon dy-oxide transaction (Wallia, 2006), limitation of leaf's extension and chlorophyll molecules destruction by sodium (Innocenti, 2009). As the chlorophyll content changes by saltiness tension would also be meaningfully effected on final photosynthesis level, perspire speed and ostium transaction (Dhanapackiam, 2010).In many studies, decrease in chlorophyll content under salinity stress mentioned which can imply to its increase in Zea mays (Chaum et al, 2009), Paulowina imperialis (Dhanapackiam, 2010), Hordeum vulgare L. (Grewall, 2010), Oriza sativa L. (Amirjani, 2010) Gossypium hirsutum L. (Desingh et al, 2007). Beinsan (2003) reported that decrease in leaf's chlorophyll content was due to increase in activity of chlorophyll destroying enzyme named chlorophylls enzyme that would lead to destruction in chlorosplast and instability of protein complexity of pigments.(Faheed, 2005). One of other reasons of decrease in photosynthesis could be considered as the gathering in sugars and increase in their density in mesophyll cells which are in fact a kind of feedback limitation on photosynthesis (Galiba, 1994).
ACKNOWLEDGEMENTS
This work was supported by Islamic Azad University of Tonekabon Branch of Iran.
References Effect of salinity stress on chlorophyll content, proline, water soluble carbohydrate, germination, growth and dry weight of three seedling barley (Hordeum vulgare L.) cultivars
- Abasi,F. (2007) Effect of salinity and drought on growth parameters of two species of plant (Aeluropus logopoides, Aeluropus littoralis). Science Journal of Islamic Azad University. J.66:121-138.
- Al-Ansari, F.M. (2003). Salinity tolerance during germination of two arid-land varients of wheat. Seed Sci & Technol. 31:597-603.
- Alizade Banat, G., Ghasemi, K., Taghizadeh, S.,(2007) Invastigation of salinity and temperature effects on germination, seedling growth and ion relation of Panicum miliaceaum. Pajouhesh and Sazandegi. J.74:115-122
- Amini, F., and Ehsanpour, A.A.(2005). Soluble proteins, praline, carbohydrates and Na+/K+ changes in two tomato (Lycopersicon esculentum Mill.) cultivars under invitro salt stress. Amercan Journal of Biochemistry and Biotechnology. 1(4): 212-216.
- Amirjani,M.R. (2010). Effect of NaCl on some Physiological Parameters of Rice. EKBS 3(1):06-16.
- Askarian, M. (2004). The effects of salinity and salinity and dryness on germination and seedling establishment in Elymus junceus and Kochia prostrate. Pajouhesh and Sazandegi. J. 64: 71-77.
- Bates, L.S., Waldran, R.P., Tear, I.D., (1973) Rapid determination of tree proline for water studes, Plant Soil, 39, 205-208.
- Beinsan, C., Camen, D., Sumalan, R., and Babau, M. (2009). Study concerning salt stress effect on leaf area dynamics and chlorophyll contein in four bean local landraces from Banat area. 44th Croatian & 4th International Symposium on Agriculture, February 16 -20, 2009. (Editors Florijan?ic?, T.;Luz?aic?, R.) pp. 416-419.
- Cha-um S, Kirdmanee C (2009). Effect of salt stress on proline accumulation, photosynthetic ability and growth characters in two maize cultivars. Pak. J. Bot. 41: 87-98.
- Chesson, P., Gebauer, L.E., Schwinning, S., Huntly, K., Wiegand, N.K., Ernest M.S.K, Sher, A., NovoPlansky A., and Weltzin, J.F., (2004). Resource Pluses, Species interactions, and diversity maintenance in arid and semi-arid environments. Ecologia 141: 236-253.
- Desingh, R., Kanagaraj, G. (2007) Influence of salinity stress on photosynthesis and antioxidative systems in two cotton varieties.Gen Appl. Plant Physiol. 33(3-4): 221-234.
- Dhanapackiam, S., and Ilyas, M. (2010). Effect of salinity on chlorophyll and carbohydrate contents of Sesbania grandiflora seedlings. Indian Journal of Science and Technology. 3(1): 64-66.
- Dizaji, N. M., Nasemie, H., and Garjani, A., (1998) Study on the anti-inflammatory effects of stachys inflata in carrageenan and formalin-induced paw oedema in the rat, Naunyn Schmiedbergs. Archives of Pharamacology Vol 358(1): 39-51.
- Ekiz, H., and Yilmaz, A. (2003). Determination of the salt tolerance of some barley genotypes and the characteristics affecting tolerance. Turk J Agric For. 27: 253-260.
- Faheed, F.A., Hassanein, A.M. and Azooz, M.M. (2005). Gradual increase in NaCl concentration overcomes inhibition of seed germination due to salinity stress in Sorghum bicolor (L.) Acta Agronomica Hungarica, 53(2), 229-239.
- Francisco G., Jhon L., Jifon S., Micaela C., James P. S. (2002) Gas exchang, Chlorophyll and nutrient contents in relation to Na+ and Cl-accumulation in sunburst mandarin grafted on different root stocks, Plant Sci. 35: 314-320.
- Galiba, G. (1994). In vitro adaptation for drought and cold hardiness in wheat. In J. Janik (ed). Plant Breeding. Rev. 12:115-161.
- Gill, K.S. (1999). Effect of soil salinity on grain filling and grain development in barley. Biologia plantarum. 21 (4): 241-244.
- Grewal, H.S. (2010). Water uptake,water use efficiency,plant growth and ionic balance of wheat, barely,canola and chickpea plants on a sodic vertosol with variable subsoil NaCl salinity. Agricultural Water Management 97: 148-156.
- Hala, M., El-Bassiouny, S., and Bekheta, M. A. (2005). Effect of salt stress on relative water content, lipid peroxidation, polyamines, amino acids, and ethylene of two wheat cultivars. International Journal of Agriculture and Biology. 3: 363-368.
- Innocenti, E.D., C. Hafsi, L. Guidi and F. Navari (2009). The effect of salinity on photosynthetic activity in potassium -deficient barley species. Journal of plant physiology. 66: 1968-1981.
- Kao, C.H. (1981). Senescense of rice leaves. VI. Comparative study of the metabolic changes of senescing turgid and water-stressed excised leaves. Plant and Cell Physiology. 22: 683-685.
- Kerepsi I. and Gabor Galiba. (2000). Osmotic and salt stress-Induced Alteration in Soluble Carbohydrate Content in wheat seedlings. Crop Science, 40; 482-487.
- Lichtenthaler, H, K. (1987) Chlorophylls and carotenoids: 30 pigments of photosynthetic biomembranes, Methods in Enzymiology 148, 350-382.
- Neto, N.B.M., C.C. Custodio, A.B. Gatti, M.R. Priolli, and V.J.M. Cardoso. (2004). Proline: use as an indicator of temperature stress in bean seeds. Crop Breeding and Applled Biotechnology 4: 330-337.
- Othman, Y., G. Al-Karaki, A.R. Al-Tawaha and A. Al-Horani. (2006) Vaviation in Germination and Jon uptake in Barley Genotypes under Salinity conditions. World Journal of Agricultural Sciences.2 (1): 11-15.
- Paknyat, H. and M. Armion. (2007) Sadium and Proline accumulation as osmoregulators of salinity in tolerance of sugar beet genotypes to salinity. Pakistan Journal of Biological Sciences 10(22): 4081-4086.
- Prabhjot, K.G., Arun D.S., Prabhjeet S., Singh B. (2001) Effects of various abiotic stress on the growth, soluble sugars and water relations of sorghum seedling grown in light and darkness. Bulg. J. Plant Physiol. 27: 72-84.
- Rezaee, M.E., Khavarinejad, R.E., Fahimi, H. (2004). Physiological response of cotton (Gossypium hirsutum L.) plants to soil salinity. Pajuhesh and Sazandegi. J. 62: 81-89
- Safarnejad, A., Salami, M., Hamidi, H., (2007) Morphological characterization of medicinal plants (Plantago ovata, Plantago psyllium) in response to salt stress. Pajouhesh and Sazandegi. J. 75: 152-160
- Shokohifard G., K.H. Sakagam. and S. Matsumoto, (1989). Effect of amending materials on growth of radish plant in salinized soil. J. Plant Nutr. 12: 1195-1294.
- Shuji Y., Ray A. B., Hassagawa P.M. (2002). Salt stress tolerance of Plants, Center for Enviro Stress Physiol.; Purdue Univ. JIRCAS working Report. 102: 25-33.
- Somogy, N, (1952) Notes on sugar determination, J. Biol. Chem., 195, 19-29.
- Tantau, H., C. Blko, B. Brettschneider, G. Melz and K. Dorfling (2004) Improved frost tolerance and winter survival in winter barley (Hordeum Vulgare L.) by in Vitro selection of proline over accumulating lines. Euphytica 139: 19-32.
- Turan M.A., Elkrim A.H.A., Taban N. and Taban S. (2010). Effect of salt stress on growth and ion distribution and accumulation in shoot and root of maize plant. African Journal of Agricultural Research 5(7), 584-588.
- Ueda A., Yuko Yamamato-Yamane and T. Takabe. (2007) Salt stress enhances proline utilization in the apical region of barley roots. Biochemical and Biophysical Research Communications. 355; 61-66.
- Usue P.L., Anabel R., Matie L., Amaia M.P., Alberto M.R. (2009) The impact of salt stress on the water statue of barley plants partially mitigated by elevated CO2. Environmental and Experimental Botany 66: 463-470.
- Walia, H., Wilson, C., and Wahid, A. (2006). Expression analysis of barley (Hordeum vulgare L.) during salinity stress. Funct Integr Genomics. 6: 143-156.
- Wenxue, W., P.E. Bilsborrrow, P. Hooley, D.A. Fincham, E. lombi and B.P. Forster (2003). Salinity induced differences in growth, ion distribution and partitioning in barley between Marythorpee and its derived mutant golden promise. Plant and Soil. 250: 183-191.
- Yazici, I., Turkan, I., Sekmen, A., Demiral, T. (2007). Salinity tolerance of Purslane (Portulaca olaracea L.) is achieved by enhanced ant oxidative system, lower level of lipid proxidation and proline accumulation. J. Env. Exp. Bot. A 453. 104-112.
- Yousfi S., Sehli M., H. Mahmoudi, C. Abdelly, M. Gharsalli (2007). Effect of salt on physiological responses of barley to iron deficiency. Plant Physiology and Biochemistry. 45: 309-314.
- Youzong H., Guoping Z., Feibo W., Jinxin C. and Meixue Z. (2006). Differences in Physiological Traits among Salt Stressed Barley Genotypes. Communications in Soil Science and Plant Analysis, 37: 557-570.
