Epipactis palustris (L.) Crantz на золоотвале Нижнетуринской ГРЭС и в естественном местообитании
 Crantz на золоотвале Нижнетуринской ГРЭС и в естественном местообитании")
Автор: Филимонова Елена Ивановна, Глазырина Маргарита Александровна, Лукина Наталия Валентиновна, Борисова Галина Григорьевна, Чукина Надежда Владимировна, Малева Мария Георгиевна, Грошева Светлана Николаевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 3 (172), 2018 года.
Бесплатный доступ
Целью данной работы являлось изучение пространственной и возрастной структур ценопопуляций, а также морфофизиологических параметров и микоризы корней Epipactis palustris (L.) Crantz, семейство Orchidaceae Juss. (дремлик болотный, сем. Орхидные), произрастающих на золоотвале Нижнетуринской государственной районной электростанции (г. Нижняя Тура, Свердловская область) и в естественном фитоценозе (прибрежная зона озера Багаряк, Сысертский район, Свердловская область). Установлено, что плотность ценопопуляций E. palustris колеблется в значительных пределах: на золоотвале - от 1 до 54 особей на м2, в контроле - от 5 до 73 особей на м2. Средняя плотность в 1,4 раза выше в ценопопуляции естественного растительного сообщества. Обе ценопопуляции являются нормальными полночленными. По критерию «дельта-омега» они относятся к молодым. Индекс восстановления был выше в естественной ценопопуляции. Морфологический анализ показал достоверные различия генеративных особей E. palustris из изученных популяций по высоте, числу цветков и числу прицветников. Высота генеративных растений, произрастающих на золоотвале, в среднем в 1,2 раза меньше, но цветков они образуют в среднем в 1,9 раза больше, чем в естественном сообществе. По толщине листа и содержанию фотосинтетических пигментов достоверных различий между популяциями не выявлено.
Золоотвал, ценопопуляции, морфофизиологические параметры, микориза
Короткий адрес: https://sciup.org/14751315
IDR: 14751315 | УДК: 581.5:581.1 | DOI: 10.15393/uchz.art.2018.127
Epipactis palustris (L.) Crantz of Nizhneturinskaya power station ash dump and natural habitats
The purpose of this research was to study the spatial and age structure of E. palustris coenopopulations. We also studied the morpho-physiological parameters and mycorrhizal roots’ association of Epipactis palustris (L.) Crantz, the family of Orchidaceae Juss., which grows on the ash dump of Nizhneturinskaya state district power station (Nizhnyaya Tura, Sverdlovsk region) and in the natural habitat (the coastal area of Bagaryak Lake, Sysertskii district, Sverdlovsk region). It is observed that the density of E. palustris coenopopulations varies considerably from 1 to 54 individuals per m2 on the ash dump and from 5 to 73 individuals per m2 in the control site. The average density was 1.4 times higher in natural habitats. Both coenopopulations were normal polynomials. They refer to the young population by the criterion of ‘delta-omega’. The recovery index was higher in the natural coenopopulation. The morphological analysis showed significant differences in generative E. palustris from the studied sites in the height, the number of flowers and the number of bracts. The height of generative plants growing on the ash dump was on the average 1.2 times smaller. At the same time they formed an average of 1.9 times higher number of flowers than the ones growing in the natural habitat. Moreover, the leaf thickness and the content of photosynthetic pigments did not reveal any significant differences between the populations from both sites.
Текст научной статьи Epipactis palustris (L.) Crantz на золоотвале Нижнетуринской ГРЭС и в естественном местообитании
Проблема сохранения и восстановления биологического разнообразия как основы устойчивости биогеоценозов является особенно актуальной для индустриально развитых регионов России, в том числе для Среднего Урала, где сосредоточены значительные площади нарушенных промышленностью земель. Восстановление растительности на подобных территориях происходит крайне медленно, видовой состав формирующихся растительных сообществ обеднен. Вместе с тем в отечественной и зарубежной литературе появляются сведения о поселении редких видов в техногенных местообитаниях в условиях сниженного фитоценотического стресса [2], [6], [12], [16], [18]. К таким растениям зачастую относятся и представители семейства Orchidaceae Juss. Оценка состояния популяций и выявление механизмов устойчивости редких видов Orchida-ceae возможны лишь при комплексном изучении их биологических и экологических особенностей, структуры и динамики ценопопуляций, консортивных связей с другими компонентами биоценоза. Комплексный подход позволяет прогнозировать последующее развитие конкретных популяций и разрабатывать наиболее эффективные меры охраны каждого вида.
Целью работы являлось изучение пространственной и возрастной структур ценопопуля-ций, а также морфофизиологических параметров и микоризы корней особей Epipactispalustris (L.) Crantz, семейство Orchidaceae Juss. (дремлик болотный, сем. Орхидные), произрастающих на золоотвале Нижнетуринской государственной районной электростанции (НТГРЭС) и в естественном фитоценозе (около озера Багаряк, Сы-сертский район, Свердловская обл.) (контроль).
-
E. palustris внесен в Красные книги 49 регионов России, в том числе Республики Карелии, Архангельской области, Республики Коми, Ханты-Мансийского автономного округа, Тюменской, Курганской, Челябинской областей и др. [2], [5], [9]. В Свердловской области данный вид имеет статус уязвимого (II категория) [10].
МAТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
E. palustris (сем. Orchidaceae Juss.) - евразийский бореальный геофит, поликарпическое, многолетнее явнополицентрическое вегетативноподвижное длиннокорневищное летнезеленое травянистое растение [2], [17]. Вид характеризуется интенсивным вегетативным размножением путем ветвления корневища и обособления его частей (партикуляции), вследствие чего числен- ность вида в местах произрастания обычно высока. Реже данный вид размножается семенами [11].
-
E. palustris - светолюбивое растение (2-я ступень шкалы Элленберга, 4–5-я ступени шкалы Ландольта) [19], [20]. Встречается на сырых почвах (8-я ступень шкалы Элленберга, 4-5-я ступени шкалы Ландольта). Растет на нейтральных и щелочных почвах (8-я ступень шкалы Элленберга, 4-5-я ступени шкалы Ландольта), кальце-фил [3].
В естественных условиях предпочитает заболоченные редколесья, заболоченные и сырые луга, болота разных типов, в том числе низинных, расположенных на выходах известняков или гипсов [2], [6], [11]. К факторам, снижающим численность вида, относятся повышенная антропогенная нагрузка и осушение болот.
На территории Свердловской области ранее было известно всего 8 местообитаний E. palustris . В 2016 году нами обнаружено новое местообитание данного вида на золоотвале НТГРЭС.
Исследования проводили в июле 2017 года на Среднем Урале. Район исследований находится в умеренно-континентальной бореальной климатической зоне. Типично лесной район, где коренная растительность представлена лиственничнососновыми травяно-кустарничковыми лесами среднетаежного типа и сосновыми лесами с примесью Larix sibirica Ledeb. и с Tilia cordata Mill. в подлеске, травяными южнотаежного типа.
Золоотвал НТГРЭС расположен в 19 км от г. Нижняя Тура (Нижнетуринский городской округ, Свердловская обл.), на месте бывшего Вогульского болота и занимает около 440 га. Золо-отвал заливался пульпой с 1992 года посекционно, работы на последнем участке завершены в 2015 году. После завершения работы каких-либо рекультивационных мероприятий на нем не проводилось.
Зола, складируемая в золоотвалы, является специфическим субстратом, обладающим рядом особенностей. По механическому составу она представлена фракциями песка и пыли с примесью измельченного шлака. Для золы характерна низкая влагоемкость, слабая теплопроводность, щелочная реакция среды, следовые количества или полное отсутствие азота, недостаточное содержание калия и в некоторых случаях – фосфора в доступной для растений форме. Содержание микроэлементов выше, чем в почве [4].
Значительная территория золоотвала обводнена и при небольшой глубине (менее 0,5 м) зарастает хвощово-рогозовыми сообществами с участием видов рода Salix. На свежих подсыхающих переувлажненных участках формируются фитоценозы с доминированием Marchantia polymor-pha L. и редким подростом Salix sp., со временем заменяющихся ивняками хвощово-вейниковыми зеленомошно-маршанциевыми.
На подсохших участках золоотвала наблюдаются разные стадии формирования лесной растительности. Зарастание золоотвала осуществляется за счет заноса семян из окружающих его со всех сторон сосновых лесов и заболоченных территорий. Антропогенное влияние на процесс самозарастания минимально.
В качестве контроля изучена ценопопуляция E. palustris в естественном фитоценозе на территории гидрологического и зоологического памятника природы «Болото Багаряк», расположенного на северном заболоченном берегу озера Багаряк (дер. Космаково, Сысертский район, Свердловская обл.).
Сбор фактического материала выполнен по общепринятым методикам [1]. Обследование территории проводили детально-маршрутным методом. Для изучения пространственной и возрастной структур ценопопуляций E. palustris случайным образом закладывали по 60 учетных площадок (S = 0,25 м2). Учитывали плотность особей вегетативного происхождения (рамет) E. palustris . Возрастные состояния E. palustris выделяли в соответствии с описанием онтогенеза М. Г. Вахрамеевой с соавторами [3].
Большая часть рамет E. palustris была обследована в природных условиях, меньшая - выкапывалась для анализа в камеральных условиях. Морфологический анализ особей проводился по следующим параметрам: высота особи (см), число листьев (шт.), длина листа (мм), ширина листа (мм). У генеративных особей кроме этого анализировали: длину цветоноса (см), длину соцветия (см), число цветков в соцветии (шт.), число прицветников (шт.), общую площадь листьев срединной формации (см2 на особь), среднюю площадь листа срединной формации второго типа (см2).
Были определены индексы возрастности (∆) [15], эффективности (и) [7] и восстановления (1в) ценопопуляций [8].
Для исследования характеристик фотосинтетического аппарата (толщина эпидермиса, мезофилла; содержание фотосинтетических пигментов) отбирали по 10 сформированных листьев с 10 генеративных особей данного вида из каждого местообитания.
Измерение толщины листа (эпидермиса и мезофилла) проводили в 30 повторностях при помощи системы Simagis Mesoplant (ООО «СИАМС», Россия) с использованием светового микроскопа Meiji MT 4300L («Meiji Techno», Япония).
Содержание фотосинтетических пигментов (хлорофиллов a, b и каротиноидов) определяли спектрофотометрически согласно [21] на Jasco V-650 («Jasco Inc.», США) в ацетоновых экстрактах (80 %).
Содержание общего азота и фосфора в листьях E. palustris измеряли колориметрически после мокрого озоления растительного материала смесью кислот: H 2 SO 4 и HClO4. Определение общего азота проводили с помощью реактива Несслера, а общего фосфора - с молибдатом аммония в кислой среде. Анализ проводили в трех биологических и трех аналитических повторностях.
Для изучения микоризы были отобраны корни 5 особей E. palustris в каждой ценопопуляции. Были приготовлены поперечные срезы корней, которые затем просматривали в поле зрения микроскопа при 100-кратном увеличении [13].
Собранный материал обработан стандартными методами математической статистики. Для обработки полученных данных использовали программные пакеты MS Office (Excel) и Statistica 6.0. Достоверность различий оценивали по критерию Манна – Уитни при уровне значимости р < 5 %.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Первая ценопопуляция E. palustris обнаружена в ивняке хвощово-вейниковом зеленомошно-маршанциевом, формирующемся на золоотвале НТГРЭС. В ценозе доминируют Salix myrsini-folia Salisb., S. triandra L., S. pentandra L., реже S. phylicifolia L. Общее проективное покрытие (ОПП) кустарникового яруса составляет 35–45 % (табл. 1). Высота ивняка в среднем достигает 0,7 м, варьируя от 0,5 до 0,8 м. На участке встречается подрост Betula pubescens Ehrh. (коэффициент встречаемости (КВ) = 62,5 %), B. pendula Roth (КВ = 52,5 %), Populus tremula L. (КВ = 20,0 %), Pinus sylvestris L. (КВ = 7,5 %) с высотой особей от 0,1 до 0,5 м. Распределение растений крайне неравномерное, ОПП травянистого яруса в среднем составляет 45 %, варьируя от 20 до 70 %. Наиболее равномерным распределением и высоким обилием характеризуются доминирующие виды: Equisetum fluviatile L. (КВ = 97,5 %, сор3), Calamagrostis epigeios (L.) Roth (КВ = 92,5 %, сор3), Epipactis palustris (КВ = 68 %, сор1–сор2), Melilotus albus Medik. (КВ = 47,5 %, sp–сор1), группами встречаются Phragmites australis (Cav.) Trin. ex Steud. (сор1 gr), Eriophorum polystachion L. (sp gr), Juncus articulates L. (sp gr), Trifolium pratense L. (sp gr), Erigeron politus Fries (sol–sp). На поверхности золы повсеместно формируется комплексный зеленомошно-маршанциевый покров, образованный многолетними наслоениями отмерших и вегетирующих талломов Marchan-tia polymorpha (покрытие 60–65 %, КВ = 95 %) и мхов Polia nutans (Hedw.) Lindb., Brium palles-cens Schleich. ex Schwaerg., Bryoerythrophyllum recurvirostre (Hedw.) Chen, Drepanocladus aduncus (Hedw.) Warnst и др. Встречаются крупные пятна Peltigera rufescens (Weis.) Humb. Формируется органогенный торфяной горизонт мощностью от 2 до 5 см.
Таблица 1
Характеристики местообитаний E. palustris
|
Характеристики |
Золоотвал НТГРЭС |
Контроль |
||
|
Х ср. |
lim |
Х ср. |
lim |
|
|
ОПП древесного яруса, % |
– |
– |
25 |
20–30 |
|
ОПП кустарникового яруса, % |
40 |
35–45 |
27 |
25–30 |
|
ПП древесно-кустарникового подроста, % |
16 |
5–35 |
20 |
0–75 |
|
ОПП травянистого яруса, % |
45 |
20–70 |
75 |
45–95 |
|
ОПП мохового яруса, % |
65 |
25–100 |
70 |
45–100 |
|
Количество видов в сообществе, шт. |
42 |
– |
36 |
– |
|
Видовое богатство шт. / 0,25 м2 |
7 |
4–10 |
5,4 |
3–9 |
Кроме E. palustris в растительном сообществе обнаружены другие редкие в Свердловской области виды сем. Orchidaceae: группы особей Dac-tylorhiza incarnata (L.) Soo (КВ = 12,5 %, sol gr) и Malaxis monophyllos (L.) Sw. (KB = 5 %, un-sol).
Вторая ценопопуляция E. palustris (контроль) произрастает на образовавшейся на месте выходов известняковых жил прогалине, рассекающей заболоченный лес северного берега озера Багаряк. Растительность прогалины отличается по видовому составу и высоте деревьев от окружающего лесного массива. Высота древесного яруса составляет 2,0–4,0 м, сомкнутости нет, ОПП – 20–30 %. В древесном ярусе преобладают Pinus sylvestris , Betula humilis Schrank, B. pubescens , B. р endula , имеющие многочисленный подрост. В кустарниковом ярусе наиболее часто встречаются Salix myrsinifolia , S. р entandra , ОПП – 25–30 %.
В травянистом ярусе (ОПП 75 %) доминируют Carex dioica L. (КВ = 85 %, сор2), C. panacea L., C. vesicaria L. (КВ = 40 %, сор1 gr), Phragmites australis (КВ - 82 %, сор2); Galium palustris L. (КВ = 24 %, sp), Epilobium palustre L. (КВ = 14 %, sol–sp), Eriophorum polystachion (sol gr); единично встречаются Pyrola rotundifolia L., Angelica sylves-tris L., Filipendula ulmaria (L.) Maxim., Potentilla erecta (L.) Raeusch., Dactylorhiza incarnata (sol gr), Neottianthe cucullata (L.) Schlechter (un). Повсеместно развит моховый покров, ОПП которого составляет 70 %.
E. palustris на исследуемом участке име-ет высокое обилие (сор1–сор2) и встречаемость (КВ = 50 %).
Сравнение видового состава изученных растительных сообществ по коэффициенту общности Съеренсена (Кс = 31 %) выявило их малое сходство. Анализ структуры экологических групп растений (по отношению к увлажнению) пока- зал, что на участке золоотвала НТГРЭС преобладают виды переувлажненных местообитаний (гигрофиты и мезогигрофиты), их доля составляет 47,7 %, доля мезофитов - 45,2 %, ксероме-зофитов – 7,1 %. В естественном местообитании (контроль) доля гигрофитов - 69,4 %, мезофитов – 30,6 %. Сравнение видового состава по структуре жизненных форм (по Раункиеру) показало, что и на золоотвале, и в естественном местообитании преобладают геофиты, гемикриптофиты и фанерофиты (33,3; 28,6; 26,2 и 38,9; 38,9; 16,6 соответственно), что также свидетельствует о формировании болотной растительности.
Таким образом, общим для исследованных местообитаний E. palustris является переувлаж-ненность субстрата, развитие гигрофитной растительности, хорошо развитое моховое покрытие, щелочная реакция среды (рН = 7,3–7,4).
Исследования ценопопуляций E. palustris показали, что как на золоотвале НТГРЭС, так и в контроле они имеют групповой тип распределения особей в пространстве (отношение дисперсии к среднему числу особей на площадке составило соответственно 14,3 и 16,4).
Плотность ценопопуляций E. palustris колеблется в значительных пределах: на золоотва-ле – от 1 до 54 особей на м2, в контроле – от 5 до 73 особей на м2. Средняя плотность особей на золоотвале составляет 16,5 особи на м2, что в 1,4 раза ниже, чем в ценопопуляции естественного растительного сообщества (23,4 особи на м2) (табл. 2). Для сравнения плотность популяций E. palustris , произрастающих на урбанизированных территориях в черте г. Тверь, составляла 40,8–41,7 особи на м2, максимальная 54–76 особей на м2 [12]. В природных популяциях данные по плотности весьма неоднозначны, варьируют от единичных особей до 100 особей на м2 [17].
Таблица 2
Характеристика ценопопуляций E. palustris
|
Местонахождение |
Плотность, особей/м2 |
Индексы ценопопуляций |
|||
|
lim |
Х ср. ± m x |
Δ |
ω |
J . |
|
|
Золоотвал НТГРЭС |
1–54 |
16,5 ± 4,0 |
0,23 |
0,57 |
1,00 |
|
Контроль |
5–73 |
23,4 ± 5,1 |
0,13 |
0,38 |
3,68 |
Примечание. Δ – индекс возрастности [15], ω – индекс эффективности [7], Jв – индекс восстановления ценопопуляций [8].
При изучении возрастной структуры цено-популяций E. palustris , произрастающих на исследуемых объектах, были выделены следующие возрастные состояния особей: ювенильные (j), имматурные (im), виргинильные (v) и генеративные (g). Генеративные особи представлены двумя возрастными состояниями: молодые (g1) и средневозрастные (g2) растения. В работе не учитывались ведущие подземный образ жизни проростки (протокорм).
Нами установлено, что все изученные цено-популяции E. palustris являются нормальными полночленными. Возрастной спектр ценопопуля-ции данного вида с золоотвала НТГРЭС является двухвершинным, с пиком в имматурном и генеративном состояниях; контрольной ценопопуля-ции – одновершинным, с пиком в имматурном состоянии (рис. 1). По исследованиям И. В. Та-таренко [14], базовый возрастной спектр E. pa-lustris является левосторонним, в котором преобладают ювенильные и имматурные особи. Для данного вида свойственно омоложение за счет партикуляции, что наблюдается в естественном местообитании. Анализ индексов возрастности и эффективности показал, что ценопопуляции
E. palustris являются молодыми (см. табл. 2). Это подтверждается также индексом восстановления ценопопуляции (1в), представляющим собой отношение плотности подроста к плотности генеративных растений (1в на золоотвале НТГРЭС - 1,0, а в контроле – 3,7) (см. табл. 2).
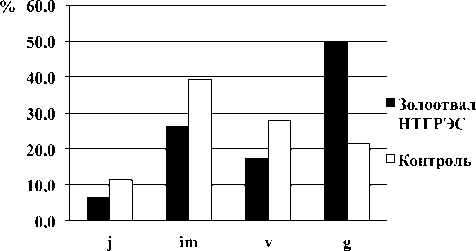
возрастные состояния
Рис. 1. Возрастные спектры ценопопуляций E. palustris
Морфологический анализ показал, что вегетативные особи изученных ценопопуляций достоверно не отличаются (табл. 3).
Таблица 3
Некоторые морфометрические показатели виргинильных особей E. palustris
|
Показатель |
Местонахождение |
Х ср. ± m x |
lim |
C v , % |
|
Высота особи, см |
Золоотвал НТГРЭС |
21,9 ± 0,7 |
18,5–26,5 |
12 |
|
Контроль |
21,1 ± 0,9 |
14,3–25,9 |
17 |
|
|
Общее число листьев, шт. |
Золоотвал НТГРЭС |
9,9 ± 0,2 |
9–11 |
8 |
|
Контроль |
9,2 ± 0,3 |
7–11 |
13 |
|
|
Число листьев срединной формации, шт. |
Золоотвал НТГРЭС |
7,2 ± 0,3 |
6–8 |
13 |
|
Контроль |
6,9 ± 0,3 |
5–9 |
17 |
|
|
Длина листьев срединной формации, см |
Золоотвал НТГРЭС |
7,6 ± 0,4 |
0,7–13,3 |
47 |
|
Контроль |
7,5 ± 0,4 |
0,3–13,8 |
54 |
|
|
Ширина листьев срединной формации, см |
Золоотвал НТГРЭС |
1,5 ± 0,1 |
0,05–3,2 |
50 |
|
Контроль |
1,4 ± 0,1 |
0,01–3,0 |
48 |
Выявлены достоверные различия генеративных особей E. palustris из нарушенного и естественного местообитаний по высоте, числу цветков и числу прицветников (табл. 4). Высота генеративных растений, произрастающих на зо-лоотвале, в среднем в 1,2 раза меньше, чем в естественном сообществе. Вместе с тем число цветков они образуют в среднем в 1,9 раза больше, что может быть связано с лучшим обеспечением на золоотвале минеральными веществами.
Общая площадь листьев срединной формации в 1,2 раза больше на золоотвале.
Изучение анатомической структуры листа E. palustris показало, что у данного вида выражен гомогенный тип строения мезофилла, характерный для гигрофитов. Толщина листовой пластинки составляет в среднем 195 мкм (см. табл. 4).
Фототрофная ткань представлена губчатым мезофиллом, который состоит из 5-6 слоев клеток и в среднем составляет 70 % от толщины листовой пластинки. Эпидермис хорошо выражен. Достоверных различий между растениями исследованных популяций по толщине листовой пластинки, толщине мезофилла и эпидермиса не выявлено, что свидетельствует об одинаковых условиях освещенности и сходном гидрологическом режиме.
E. palustris относится к так называемым зеленым орхидеям, фотосинтезирующим в течение надземных стадий развития. Функционирование фотосинтетического аппарата растений определяется количеством фотосинтетических пигментов и их соотношением. По содержанию хлорофиллов и каротиноидов между растениями
Таблица 4
Морфометрические и анатомические показатели генеративных особей E. palustris
|
Показатель |
Местонахождение |
Х ср. ± m x |
lim |
C v , % |
|
|
Высота особи*, см |
Золоотвал НТГРЭС |
45,1 ± 2,5 |
27,5–56,2 |
19 |
|
|
Контроль |
54,2 ± 3,3 |
38,0–73,5 |
21 |
||
|
Общее число листьев, шт. |
Золоотвал НТГРЭС |
9,4 ± 0,2 |
8–11 |
8 |
|
|
Контроль |
8,8 ± 0,2 |
8–10 |
7 |
||
|
Число листьев срединной формации, шт. |
Золоотвал НТГРЭС |
6,4 ± 0,2 |
6–8 |
11 |
|
|
Контроль |
6,3 ± 0,2 |
5–7 |
10 |
||
|
2 >s 8 н о О Щ S S к S s у с8 Ц 5 к § S ^ Л м О О =5 "^ |
1 типа* |
Золоотвал НТГРЭС |
2,3 ± 0,3 |
0,5–4,9 |
54 |
|
Контроль |
3,9 ± 0,5 |
1,2–7,8 |
54 |
||
|
2 типа* |
Золоотвал НТГРЭС |
8,0 ± 0,5 |
3,5–11,5 |
33 |
|
|
Контроль |
10,2 ± 0,4 |
6,6–14,7 |
21 |
||
|
3 типа |
Золоотвал НТГРЭС |
9,3 ± 0,3 |
5,6–11,6 |
19 |
|
|
Контроль |
8,8 ± 0,6 |
1,7–13,3 |
34 |
||
|
О S И s t; щ s cd s g щ ч 2 s % g О Рч Он S ° ° 3 |
1 типа |
Золоотвал НТГРЭС |
1,9 ± 0,3 |
0,8–3,2 |
48 |
|
Контроль |
2,2 ± 0,2 |
1,2–3,8 |
35 |
||
|
2 типа* |
Золоотвал НТГРЭС |
3,5 ± 0,2 |
1,6–4,8 |
26 |
|
|
Контроль |
2,9 ± 0,1 |
1,9–3,8 |
18 |
||
|
3 типа* |
Золоотвал НТГРЭС |
2,0 ± 0,1 |
0,6–3,5 |
37 |
|
|
Контроль |
1,6 ± 0,1 |
0,6–2,8 |
39 |
||
|
Длина цветоноса, см |
Золоотвал НТГРЭС |
8,8 ± 0,4 |
5,4–11,1 |
17 |
|
|
Контроль |
9,9 ± 1,0 |
4,2–16,5 |
34 |
||
|
Длина соцветия, см |
Золоотвал НТГРЭС |
12,1 ± 1,2 |
5,1–18,2 |
35 |
|
|
Контроль |
9,7 ± 1,2 |
4,1–15,7 |
42 |
||
|
Число цветков*, шт. |
Золоотвал НТГРЭС |
13,9 ± 1,6 |
4–24 |
41 |
|
|
Контроль |
7,4 ± 0,8 |
3–11 |
34 |
||
|
Число прицветников*, шт. |
Золоотвал НТГРЭС |
16,8 ± 1,5 |
11–27 |
30 |
|
|
Контроль |
9,3 ± 0,8 |
6–14 |
29 |
||
|
Длина прицветников, см |
Золоотвал НТГРЭС |
1,5 ± 0,1 |
0,2–6,0 |
66 |
|
|
Контроль |
1,5 ± 0,1 |
0,2–5,7 |
66 |
||
|
Ширина прицветников, см |
Золоотвал НТГРЭС |
0,4 ± 0,01 |
0,1–1,1 |
43 |
|
|
Контроль |
0,4 ± 0,02 |
0,1–0,9 |
40 |
||
|
Общая площадь листьев срединной формации, см2/особь |
Золоотвал НТГРЭС |
72,9 ± 10,0 |
21,3–115,0 |
43 |
|
|
Контроль |
62,6 ± 5,7 |
39,3–94,2 |
29 |
||
|
Средняя площадь листа срединной формации второго типа, см2 |
Золоотвал НТГРЭС |
17,1 ± 1,3 |
4,6–26,1 |
38 |
|
|
Контроль |
13,8 ± 1,1 |
2,6–27,0 |
37 |
||
|
Толщина эпидермиса, мкм |
Золоотвал НТГРЭС |
55,1 ± 3,0 |
31,9–66,7 |
19,5 |
|
|
Контроль |
63,0 ± 3,7 |
42,5–85,0 |
21,1 |
||
|
Толщина мезофилла, мкм |
Золоотвал НТГРЭС |
145,2 ± 6,1 |
127,5–189,1 |
15,0 |
|
|
Контроль |
127,8 ± 3,1 |
106,3–148,8 |
8,7 |
||
Примечание. * – различия между популяциями достоверны при р < 0,05.
E. palustris с золоотвала и естественного фитоценоза достоверных различий также не было выявлено (рис. 2).
При этом содержание хлорофилла а у E. pa-lustris изученных популяций было в среднем в 2,5 раза выше по сравнению с хлорофиллом b, что является характерным для наземных растений и гигрофитов с надводными листьями.
Важную роль при характеристике рабо-ты пигментного комплекса играет отноше-ние хлорофиллов к каротиноидам. Это соотношение варьирует в широких пределах (2–7)
и определяется разными факторами, но чаще всего условиями освещения [22]. Известно, что это соотношение реагирует на изменение условий среды. Отношение суммы хлорофиллов к каротиноидам в листьях E. palustris как на золоотвале, так и в естественном местообитании было примерно одинаковым (в среднем 6,5), что еще раз свидетельствует о сходных условиях освещенности, водообеспечения и степени обеспеченности питательными веществами.
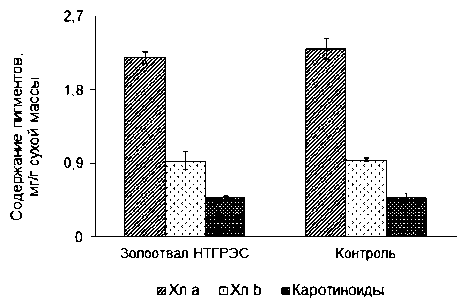
Рис. 2. Содержание фотосинтетических пигментов в листьях генеративных особей E. palustris.
Различия между популяциями недостоверны при p < 0,05
Известно, что E. palustris , населяющий открытые местообитания, вступает в симбиоз с Rhizoctonia -подобными грибами .
По исследованиям М. Г. Вахрамеевой с соавторами [3], у E. palustris грибная инфекция корней очень слабая, но на бедных почвах и при низкой освещенности степень микотрофности возрастает. Интенсивное микоризообразование наблюдается также на очень сырых почвах с низким содержанием гумуса [24]. Нами в корнях E. palustris обнаружена орхидная микориза, представленная пелотонами, занимающая в ценопопу-ляции с золоотвала от 80 до 100 %, в контроле – от 70 до 100 % корней.
Экспериментально показано существенное влияние орхидной микоризы на фосфорное и особенно азотное питание растений [23].
Для E. palustris из изученных популяций было характерно повышенное содержание общего азота: в листьях растений с золоотвала оно составило 5,1 %, а в естественном местообитании – 4,0 % от сухой массы. По содержанию общего фосфора популяции достоверно не отличались (в среднем оно составило 0,3 % от сухой массы).
ЗАКЛЮЧЕНИЕ
На Среднем Урале для Epipactis palustris (редкий вид) установлено новое местообитание в ивняке хвощово-вейниковом зеленомошно-маршанциевом, формирующемся в процессе са-мозарастания на золоотвале Нижнетуринской ГРЭС.
Местообитания E. palustris на золоотвале и в естественном фитоценозе (в прибрежной зоне озера Багаряк) характеризуются переувлажненным субстратом, развитием гигрофитной растительности и щелочной реакцией среды.
Показано, что ценопопуляции E. palustris в растительном сообществе на золооотвале и в естественном фитоценозе имеют высокую плотность: от 1 до 54 и от 5 до 73 особей на м2 соответственно. Средняя плотность особей в 1,4 раза выше в ценопопуляции в естественном растительном сообществе. Обе ценопопуляции являются нормальными полночленными. По критерию «дельта-омега» они относятся к молодым. Индекс восстановления выше в контрольной ценопопуляции. Морфологический анализ показал достоверные различия между генеративными особями E. palustris из изученных популяций по высоте, числу цветков и прицветников. Высота генеративных растений, произрастающих на золоотвале, в среднем в 1,2 раза меньше, но цветков они образуют в среднем в 1,9 раза больше, чем в естественном сообществе. Обильное цветение и плодоношение орхидей в антропогенных местообитаниях отмечается многими авторами [18].
В естественных местообитаниях росту и размножению орхидей часто препятствует развитие травянистого яруса и образование плотной дернины, ограничивающей газообмен между почвой и атмосферой. В нарушенных местообитаниях отсутствие задернения и хорошо развитой подстилки, а также слабая ценотическая конкуренция способствуют прорастанию семян орхидных.
Таким образом, нарушенные территории могут быть временными «убежищами» для сохранения генофонда многих видов сем. Orchi-daceae.
Дальнейшее развитие ценопопуляции E. pa-lustris на золоотвале зависит от особенностей трансформации фитоценоза, изменения гидрологического режима субстрата, а также от антропогенной деятельности на данном объекте.
* Работа выполнена при финансовой поддержке со стороны Министерства образования и науки Российской Федерации в рамках выполнения государственного задания УрФУ № 6.7696.2017/БЧ.
EPIPACTIS PALUSTRIS (L.) CRANTZ OF NIZHNETURINSKAYA POWER STATION ASH DUMP AND NATURAL HABITATS
The purpose of this research was to study the spatial and age structure of E. palustris coenopopulations. We also studied the morpho-physiological parameters and mycorrhizal roots’ association of Epipactis palustris (L.) Crantz, the family of Orchidaceae Juss., which grows on the ash dump of Nizhneturinskaya state district power station (Nizhnyaya Tura, Sverdlovsk region) and in the natural habitat (the coastal area of Bagaryak Lake, Sysertskii district, Sverdlovsk region). It is observed that the density of E. palustris coenopopulations varies considerably from 1 to 54 individuals per m2 on the ash dump and from 5 to 73 individuals per m2 in the control site. The average density was 1.4 times higher in natural habitats. Both coenopopulations were normal polynomials. They refer to the young population by the criterion of ‘delta-omega’. The recovery index was higher in the natural coenopopulation. The morphological analysis showed significant differences in generative E. palustris from the studied sites in the height, the number of flowers and the number of bracts. The height of generative plants growing on the ash dump was on the average 1.2 times smaller. At the same time they formed an average of 1.9 times higher number of flowers than the ones growing in the natural habitat. Moreover, the leaf thickness and the content of photosynthetic pigments did not reveal any significant differences between the populations from both sites.
-
* The research is conducted with the financial support from the Ministry of Education and Science of the Russian Federation within a framework of the state assignment. Project № 6.7696.2017.
Список литературы Epipactis palustris (L.) Crantz на золоотвале Нижнетуринской ГРЭС и в естественном местообитании
- Борисова М. А., Маракаев О. А. Редкие виды растений: практика исследований в природе: Учеб.-метод. пособие. Ярославль: Яросл. гос. ун-т, 2015. 64 с.
- Варлыгина Т. И., Вахрамеева М. Г., Татаренко И. В. Орхидные России (биология, экология и охрана). М.: Товарищество научных изданий КМК, 2014. 437 с.
- Вахрамеева М. Г., Варлыгина Т. И., Баталов А. Е., Тимченко И. А., Богомолова Т. И. Род Дремлик//Биологическая флора Московской области. Вып. 13. М.: Полиэкс, 1997. С. 50-87.
- Глазырина М. А., Лукина Н. В., Филимонова Е. И., Чукина Н. В., Борисова Г. Г., Перевозчикова Е. Н. Fragaria vesca L. на золоотвалах Среднего Урала//Ученые записки Петрозаводского государственного университета. 2017. № 8 (169). С. 35-43.
- Дьячкова Т. Ю., Кузнецов О. Л. Видовой состав сообществ с Epipactis palustris (L.) Crantz на болотах Карелии//Ученые записки Петрозаводского государственного университета. 2017. № 8 (169). С. 49-53.
- Ефимов П. Г. Орхидные северо-запада европейской России (Ленинградская, Псковская, Новгородская области). 2-е изд., испр. и доп. М.: Товарищество научных изданий КМК, 2012. 220 с.
- Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений//Экология. 2001. № 1. С. 3-7.
- Жукова Л. А., Полянская Т. А. О некоторых подходах к прогнозированию перспектив развития ценопопуляций растений//Вестник ТвГУ. 2013. Вып. 32. № 31. С. 160-171.
- Красная книга Республики Карелия. Петрозаводск: Карелия, 2007. 368 с.
- Красная книга Свердловской области: животные, растения, грибы/Отв. ред. Н. С. Корытин. Екатеринбург: Баско, 2008. 256 с.
- Мамаев С. А., Князев М. С., Куликов П. В., Филиппов Е. Г. Орхидные Урала: систематика, биология, охрана. Екатеринбург: УрО РАН, 2004. 124 с.
- Пушай Е. С., Разумова И. С. Изучение и оценка современного состояния популяций Epipactis palustris (L.) Crantz на территории г. Тверь//Вестник ТвГУ. Серия: Биология и экология. 2007. Вып. 4. № 8 (36). С. 106-109.
- Селиванов И. А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 232 с.
- Татаренко И. В. Орхидные России: жизненные формы, биология, вопросы охраны. М.: Аргус, 1996. 207 с.
- Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов//Научные доклады высшей школы. Биол. науки. 1975. № 2. С. 7-34.
- Филимонова Е. И., Лукина Н. В., Глазырина М. А. Орхидные в техногенных системах Урала//Экосистемы, их оптимизация и охрана. 2014. Вып. 11. С. 68-75.
- Чупракова Е. И., Савиных Н. П. Биоморфология Epipactis palustris (L.) Crantz с позиции охраны вида//Научные ведомости. Серия: Естественные науки. 2011. № 9 (104). Вып. 15/1. С. 23-28.
- Adamowski W. Expansion of nativ Orchids in anthropogenous habitats Polish//Botanical Studies. 2006. Vol. 22. P. 35-44.
- Ellenberg H., Weber H. E., Düll R., Wirth V., Werner W., Paulißen D. Zeigerwerte von Pflanzen in Mitteleuropa//Scripta Geobotanics. Vol. 18. Gottingen: Verlag Erich Goltze KG, 1991. 248 s.
- Landolt E. Ökologische Zeigerwerts zur Sweizer Flora//Veroff. Geobot. Zurich: Inst. EtH, 1977. H. 64. 208 s.
- Lichtenthaler H. Chlorophylls and carotenoids: pigments of photosynthetic membranes//Meth. Enzymol. 1987. Vol. 148. P. 350-382.
- Ronzhina D. A., Nekrasova G. F., P’yankov V. I. Comparative characterization of the pigment complex in emergent, floating, and submerged leaves of hydrophytes//Russian J. Plant Physiol. 2004. Vol. 51. No 1. P. 21-27.
- Smith S. E., Read D. J. Mycorrhizal symbiosis (Third Edition). N. Y: Academic Press, 2008. 787 p.
- Summerhayes V. S. Wild orchids of Britain. London, 1951. 290 p.
