Evaluation of the effect of oxidative stress on roots of red beet ( Beta vulgaris L.)
")
Author: Kolesnikova E.V., Ozolina N.V., Nurminsky V.N., Nesterkina I.S., Sitneva L.A., Lapteva T.I.
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 4 т.10, 2014.
Free access
In the present investigation it was studied the effect of oxidative stress on root of red beet. The degree of stress exposure was assessed at various levels: in tissues of roots (conductometric method), on isolated vacuoles (time-lapse video recording), and transport activity of the vacuolar membrane enzyme, namely V-H +-ATPase (fluorescent probe method). The obtained results allow to conclude the negative effects of oxidative stress on different levels of plant organization. There were several effects: a significant increase of outflow of electrolytes from the tissues of roots, a great decrease of half-life time of isolated vacuoles, and a 2 times reduction of transport activity of tonoplast H +-ATPase. Thus, the methods used in this study can be applied to assess the intensity of the impact of oxidative stress on plants.
Enzyme activity, electrolyte outflow, root of red beet, oxidative stress, stability of vacuoles
Short address: https://sciup.org/14323909
IDR: 14323909
Text of the scientific article Evaluation of the effect of oxidative stress on roots of red beet ( Beta vulgaris L.)
Как известно, влияние неблагоприятных факторов внешней среды на растения приводит к нарушению метаболизма и существенному снижению продуктивности растений. Изучение изменений, которые происходят в ответ на стрессовые воздействия и способы их преодоления является одной из основных задач биологии (Shakirova, 2001). Любые абиотические стрессы приводят к окислительному – одному из наиболее неблагоприятных видов стресса для растительной клетки (Menshikova et al., 1993). Существует достаточно большое количество методов, позволяющих оценить величину негативного воздействия, вызванного стрессом, часть из которых была использована в настоящем исследовании. Цель данного исследования состояла в оценке влияния окислительного стресса на разных уровнях организации растительного организма. Для этого была поставлена задача провести сравнительную оценку воздействия окислительного стресса начиная с тканевого уровня (выход электролитов) и кончая молекулярным (активностью мембраносвязанного фермента), используя методы кондуктометрии, цейтраферной видеосъёмки и спектрофлуориметрию.
MATERIALS AND METHODS
Объектом исследования служили корнеплоды и изолированные вакуоли столовой свёклы ( Beta vulgaris L.) сорта Бордо, выращенной на опытном участке института. Корнеплоды находились на стадии покоя, и хранились при +4°С. Изолированные вакуоли и везикулы вакуолярных мембран получали с помощью метода (Salyaev et al ., 1981). Окислительный стресс создавали реакцией Фентона, приводящей к образованию гидроксильных радикалов. Для этого к тканевым дискам, в суспензию изолированных вакуолей или к осадку мембранных везикул добавляли 50 мМ
Н 2 О 2 и 1 мМ FeSO 4 , после чего инкубировали 30 мин. В контрольном варианте инкубацию проводили в дистиллированной воде, а полученные результаты принимали за 100%.
Для оценки устойчивости ткани корнеплодов к окислительному стрессу использовали кондуктометрический метод (Grischenkova, Lukatkin, 2005). Диски ткани корнеплодов инкубировали 30 минут в дистиллированной воде и в растворе 50 мМ Н 2 О 2 . Сразу после окончания стрессового воздействия определяли проницаемость клеточных мембран по выходу электролитов из тканевых дисков в дистиллированную воду с использованием кондуктометра (ОК-104, Radelkis) с платиновым электродом. Затем раствор с растительным материалом доводили до кипения (для разрушения мембран) и давали ему остыть до комнатной температуры, после чего объём доводили до исходной величины и по электропроводности определяли полный выход электролитов. Степень выхода электролитов рассчитывали в процентах от полного выхода.
Влияние окислительного стресса на барьерные свойства мембран (стабильность мембран) изучали с использованием экспериментальной установки собственного изготовления, позволяющей получать серии компьютерных изображений (цейтраферная видеосъемка, частота кадров 0.1 мин-1 (1 кадр за 10 мин)), отражающих динамику процесса разрушения вакуолей (Nurminsky et al., 2004). С помощью компьютерной обработки данных строили график зависимости количества сохранившихся вакуолей (%) от времени и рассчитывали значения периода полураспада изолированных вакуолей, в течение которого разрушаются 50% вакуолей (Т1/2).
Воздействие окислительного стресса на транспортную функцию протонной помпы определяли на фракции вакуолярных мембран (Salyaev et al., 1981). Уровень протон-транслоцирующей активности Н+-АТФазы тонопласта оценивали по изменению рН везикул тонопласта при помощи флуоресцентного зонда акридинового оранжевого. За сдвигами рН следили по изменению интенсивности флуоресценции этого зонда при длине волны возбуждающего и испускаемого света 493 и 540 нм соответственно. Эксперименты проводили на спектрофлуориметре (RF-5301PC, Shimadzu). В кювету поэтапно вносили 2.5 мл инкубационного раствора и 0.05 мл осадка везикул тонопласта. Инкубационный раствор содержал: 20 мМ Трис/МЭС, 50 мМ КСl, 280 мМ маннит, 5 мкМ акридиновый оранжевый, 3 мМ МgCl2, рН 7.3. Содержание белка определяли по методу Бредфорд (Bradford, 1976). В среднем на 0.05 мл образца приходилось 20-30 мкг белка. В среду инкубации вносили 3 мМ АТФ (Трис). Для доказательства специфичности проводимых исследований во всех экспериментах использовали общепринятый ингибитор Н+-АТФазы – 50 мМ КNO3, который подавлял активность этого фермента, а также протонофор 10 мкМ карбонилцианид 3-хлорофенилгидразон (CCCP), который полностью предотвращал тушение флуоресценции, возникающее при функционировании фермента. Транспортная активность измерялась в общепринятых единицах (%ΔF/мг белка/мин), где ΔF - тушение флуоресценции акридинового оранжевого (Braun et al., 1986).
В работе применяли следующие реактивы фирмы “Sigma”: 2-N-морфолино-этансульфоновую кислоту (МЭС), аденозинтрифосфат (АТФ, натриевая соль), Трис, 2-меркаптоэтанол, карбонилцианид 3-хлорофенилгидразон (СССР). Акридиновый оранжевый фирмы “Aldrich”. Остальные реактивы были отечественного производства квалификации ХЧ.
На графиках представлены средние арифметические значения величин и их стандартные отклонения, которые были получены в пяти независимых экспериментах, подсчитанные с помощью программы Microsoft Excel.
RESULTS AND DISCUSSION
На первом этапе исследований измеряли выход электролитов из дисков ткани корнеплодов. Этот параметр позволяет оценить проницаемость клеточных мембран и их устойчивость к стрессу. При действии неблагоприятных факторов повышается утечка электролитов из тканей, что свидетельствует о нарушении избирательной проницаемости мембран растительной клетки (Prikhodko, 1977). Это может быть связано с изменениями в структуре мембран или в состоянии их липидной составляющей (Bertin et al., 1996). Проведённые эксперименты показали, что при воздействии окислительного стресса на высечки из корнеплодов столовой свёклы существенно (более чем в 2 раза) увеличивался выход электролитов (Рис. 1), что говорит о значительных нарушениях структуры мембран, которые произошли в ответ на это стрессовое воздействие.
Сходные результаты были получены и при оценке стабильности изолированных вакуолей, которая исследовалась методом цейтраферной видеосъёмки. Результаты показывают, что при окислительном стрессе период полураспада вакуолей уменьшался более чем в 2 раза (Рис. 2), то есть темпы их разрушения были очень высокими. Исходя из того, что стабильность липидного бислоя и клеточной мембраны в целом определяется липидными порами (Antonov et al., 1999), разрыв тонопласта, прежде всего, является следствием изменений в липидном матриксе мембраны. Причины нарушения целостности липидного бислоя могут быть различны, например, перекисное окисление липидов, гиперактивация катаболических ферментов, изменения в липид-белковых взаимодействиях и др. Эти процессы приводят к образованию стрессовых пор, их последующему расширению и разрыву мембраны. Стрессирование и разрыв мембраны могут быть также вызваны нарушением водно-осмотического равновесия вакуоли, вследствие разрушения крупных молекул внутри вакуоли (например, при гидролизе белков) и повышения осмотической концентрации внутривакуолярного содержимого, что должно привести к поступлению воды в вакуоль, увеличению ее объема и фатальному растяжению мембраны. В наших экспериментах при окислительном стрессе, вызванном пероксидом водорода, нарушалась структура тонопласта, что приводило к формированию и расширению липидных пор в мембране, и в результате чего, она разрушалась за короткое время. Возможно, именно с нарушением барьерной функции тонопласта, которая отслеживалась в этих экспериментах, и связан более высокий выход электролитов из ткани корнеплодов, как это было показано ранее.
Для сравнительной оценки влияния окислительного стресса на молекулярном уровне было проведено изучение изменения транспортной активности протонной помпы вакуолярной мембраны – Н+-АТФазы, которая являются одними из основных мембранных белков тонопласта и обеспечивает поступление в вакуоль большого числа метаболитов (Martinoia еt al ., 2007). На долю Н+-AТФазы приходится 15% всех белков тонопласта (Maeshima, 2001). Эксперименты показали, что в условиях окислительного стресса транспортная активность этого фермента падает более чем на 40%, что говорит о существенном нарушении метаболических процессов (Рис. 3). Можно предположить, что пероксид водорода вызывает окисление сульфгидрильных групп гидрофильного комплекса фермента, вследствие чего и происходит частичное инактивирование Н+-AТФазы (Seidel et al ., 2012). Кроме, того на уровень активности протонной помпы могло оказать влияние и изменение липидного окружения (Los et al., 2007), что приводит к перестройке конформации фермента и тем самым сказывается на активности (Palladina, Simchuk,
Проведённые эксперименты показали, что окислительных стресс оказывал значительное негативное влияние на разных уровнях организации растения: заметно нарушались барьерные свойства мембран растительных клеток, резко падала стабильность изолированных вакуолей и активность мембранных транспортных систем. Устойчивость к этому виду стресса у корнеплодов столовой свёклы была незначительной. Все использованные в экспериментах методы могут применяться для оценки интенсивности воздействия окислительного стресса на растительные объекты.
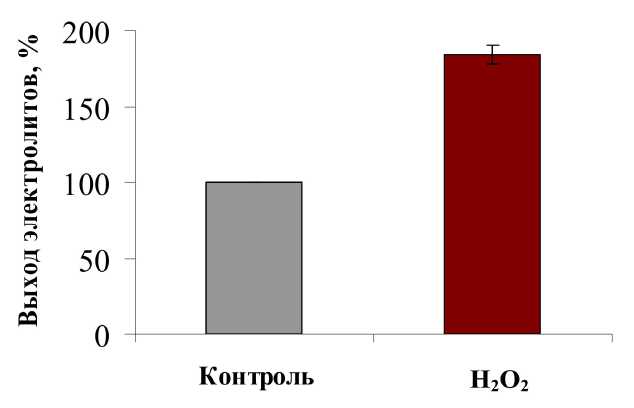
Figure 1 . Влияние окислительного стресса на выход электролитов из дисков корнеплодов столовой свёклы в присутствии пероксида водорода (50 мМ), % от контроля.
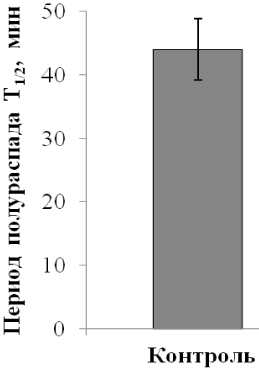

Figure 2 . Влияние окислительного стресса на период полураспада изолированных вакуолей (Т 1/2 ). Результаты получены методом цейтраферной видеосъемки.
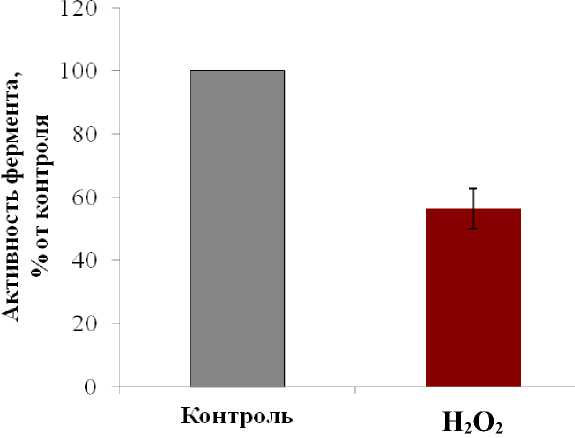
Figure 3 . Транспортная активность Н+-АТФазы тонопласта в присутствии пероксида водорода (50 мМ), % от контроля.
ACKNOWLEDGMENT
Исследование выполнено при финансовой поддержке РФФИ в рамках научных проектов № 14-04-31103 мол_а и № 14-04-31080 мол_а.
В работе использовалось оборудование Байкальского аналитического центра (ЦКП) ИНЦ СО РАН.
References Evaluation of the effect of oxidative stress on roots of red beet ( Beta vulgaris L.)
- Antonov V.F., Anosov A.A., Bogatyreva N.E. et al. (1999) Lipid pores: stability and membrane permeability. 2nd Congress of biophysicists, Moscow, Russia, 23-27 Aug., 1999: abstracts. dokl., V. 2, 476-477
- Bertin P., Bouharmont J., Kinet J. (1996) Somaclonal variation and improvement in chilling tolerance in rice. Changes in chilling-induced electrolyte leakage. Plant Breeding. 115(4): 268-272
- Braun Y., Hassidim M., Lerner H., Reinhold L. (1986) Studies on H+-translocating ATPases in plants of varying resistance to salinity. Plant Physiol., 81: 1050-1056
- Brеdford D.P. (1976) A rapid and sensitive method for the quantitation of protein utilising the principl of protein-dye binding. Anal. Biochem., 72: 248-254
- Grischenkova N.N., Lukatkin A.S. (2005) Determination of resistance of plant tissues to abiotic stresses using conductometric method. Povolzhskiy journal of ecology, 1: 3-11
- Los D.A. (2007) Perception of stress signals by biological membranes. In the book: Problems of regulation in biological systems. Biophysical aspects. Ed. Rubin A.B. M-Izhevsk: center "Regulatory and chaotic dynamics", Institute of computer science, 329-360
- Maeshima M. (2001) Tonoplast transporters: organization and function. Annu Rev. Plant Physiol. Plant Mol. Biol., 52: 469-497
- Martinoia E., Maeshima M., Neuhaus H.E. (2007) Vacuolar transporters and their essential role in plant metabolism. Journal of Experimental Botany, 58(1): 83-102
- Menshikova E.E., Zenkov N.A. (1993) Antioxidants and inhibitors of radical processes. Biology Bulletin Reviews, 113(4): 183-205
- Nurminsky V.N., Korzun, A.M., Rosinov SV, Salyaev R.K. (2004) Computer time-lapse video of fractions of isolated vacuoles. Biomedical chemistry, 50: 180-187
- Palladina T.A., Simchuk E.E. (1990) Effect of sterols on the activity of H+-ATPase of plasma membranes of root cells of maize. Dokl. USSR, 314(4): 1018-1020
- Prikhodko N.V. (1977) Changes in permeability of cell membranes as a general level of nonspecific mechanisms of plant responses to external influences. Physiology and biochemistry of cultivated plants, 9(3): 301-309
- Salyaev R.K., Kuzevanov V.A., Chaptagaev C.B, Kopytchuk V.N. (1981) Isolation and purification of vacuoles and vacuolar membranes from plant cells. Russian Journal of Plant physiology, 28: 1295-1305
- Seidel T., Scholl S., Krebs M., Rienmuller F., Marten I., Hedrich R., Hanitzsch M., Janetzki P., Dietz K.-J., Schumacher K. (2012) Regulation of the V-type ATPase by redox modulation. Biochem. J., 448: 243-251
- Shakirova F.M. (2001) Nonspecific resistance of plants to stress factors and its regulation. Ufa: Gelem, 160 p
