Факторная ординация лесных экосистем на зональной границе (на примере Комсомольского-на-Амуре заповедника)
")
Автор: Петренко Полина Сергеевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общие проблемы экологии
Статья в выпуске: 3 т.15, 2013 года.
Бесплатный доступ
С помощью теоретико-информационного метода бинарной ординации рассчитаны матрицы совместного распределения геоморфологических, почвенных и фитоценотических свойств экосистем заповедника «Комсомольский» по 17 геокомпонентным показателям. На этой основе проведен анализ ведущих локальных механизмов формирования экосистем заповедника, рассматриваемых на уровне межкомпонентных ландшафтных связей.
Лесная экосистема, бинарная ординация, геокомпонентные признаки, экологический оптимум
Короткий адрес: https://sciup.org/148201761
IDR: 148201761 | УДК: 911.52+91.001.5+502.4
Factor ordination of forest ecosystems at zonal boundary (for example of Komsomolsky state nature reserve)
This paper outlines experience in building combined distribution’s matrixes of geomorphological, soil, and phytocenosis characters within 17 geocompanent indices for the territory of Komsomolsky reserve. The calculation was made with the use the theory information method of binary ordination. On this base main inside landscape organizing mechanisms of reserve’s ecosystems were analyzed. They were shown as interrelation landscape links for local level.
Текст научной статьи Факторная ординация лесных экосистем на зональной границе (на примере Комсомольского-на-Амуре заповедника)
Государственный природный заповедник «Комсомольский» расположен на зональной границе бореальных и суббореальных ландшафтов Юга Дальнего Востока [6] и входит, таким образом, в систему зональных геоэкотонов, по определению [5], столь характерных для восточной окраины Евразийского материка [8, 12, 16]. Заповедник охватывает один из сохранившихся еще в Нижнем Приамурье крупных массивов темнохвойных и хвойно-широколиственных лесов, насыщенных третичными реликтовыми растениями. Здесь соседствуют представители трех основных флор [18]: охотско-камчатской (ель аянская), восточно-сибирской мерзлотно-таежной (лиственница даурская) и маньчжурской (липа амурская, кедр кокорейский). Предполагается, что, благодаря своему экотонному положению, лесные сообщества заповедника должны находиться в критических состояниях, далеких от оптимума их функционирования и структурной устойчивости, и отличаться повышенной чувствительностью к внешним воздействиям. Такие пограничные фитоценологические комплексы являются благоприятными объектами для биоэкологиче-ского и геосистемного мониторинга [2] на основе изучения межкомпонентных ландшафтных связей как проводников внешних воздействий на состояние гео(эко)-систем [8, 10].
С целью количественной оценки моносистем-ной организации природно-территориальных комплексов заповедника «Комсомольский» автором были проведены на его территории крупномасштабные ландшафтно-экологические исследования по методологии, разработанной в Институте экологии Волжского бассейна РАН [9, 10]. С
Петренко Полина Сергеевна , научный сотрудник государственного природного заповедника «Комсомольский».
помощью теоретико-информационного метода бинарной ординации удалось вскрыть некоторые механизмы формирования почвенных и фитоце-нотических компонентов природных экосистем заповедника. В настоящем сообщении излагаются результаты проведенных исследований.
ОБЪЕКТЫ И МЕТОДЫ АНАЛИЗА
На территории заповедника было заложено 23 пробных площади в различных типах локальных местоположений – геотопах, по определению [11, 15]. Геотопы образуют систему местных ландшафтных сопряжений [3], или катен, – от элювиальных и трансэлювиальных типов местоположений до аккумулятивных и супераквальных. Одновременно пробные площади охватывали наиболее типичные для равнинных и низкогорных ландшафтов Нижнего Приамурья растительные формации. Объектами исследования послужили следующие растительные комплексы в ранге фито-ценомеров локального уровня, согласно [16]: 1) мелколиственные леса с примесью широколиственных пород (Мл); 2) кедровошироколиственные и широколиственные леса (Шл); 3) лиственничные и широколиственнолиственничные леса (ЛиШЛл); 4) темнохвойные елово-пихтовые леса (ЕПл); 5) осоково-сфагновая марь с вересковыми кустарничками и вейником (МСМ); скалистые луга (СЛ). Под лесными сообществами распространены почвы буроземного типа: в южнотаежных и подтаежных лесах буротаежные (в том числе иллювиально-гумусовые), а в субнеморальных – бурые почвы (в том числе бурые горно-лесные).
По каждой пробной площади, характеризующей определенный биогеоценоз (ландшафтную фацию), получены эмпирические данные, включающие 16 геокомпонентых признаков, объединенных в 4 блока: геоморфологический, почвенный, фитоценотический и гидротермический.
Сбор фактического материала проводился согласно общепринятым методам полевых ландшафтных и биогеоценологических исследований [1, 4, 13 и др.]. Информационная мера сложности почвенного профиля, а также меры флористического разнообразия древесного яруса и травянистого покрова вычислялись по известной формуле К. Шеннона [7].
Согласно рекомендациям [10, 14], каждый качественный и количественный признак был подразделен на 4–6 градаций, с присвоением соответствующих баллов. Качественный признаки представлены в таблице. Для квантования количественных признаков использована зависимость:
i = ( ^ max * ^ min )/ n
, ( 1)
где: i – величина интервала; x max и x min – соответственно максимальное и минимальное значения признака в данной совокупности (выборке); n – число групп. Полученные значения были округлены до целых чисел.
С помощью метода бинарной ординации [15, 17] находились экологические ниши состояний изучаемого явления A в множестве состояний того или иного фактора B . Для этого по матрице вероятностей (частот) совместной встречаемости различных значений А и В вычислялись частные коэффициенты связи по формуле [7]:
C ( aj b j ) =
p ( ai / bj ) p ( ai )
где: p ( a i /b i ) – условная вероятность i -го состояния явления А при j -том состоянии фактора
В; p(ai) – априорная вероятность i-го состояния явления А. Связь считается значимой при С(ai/bi)>1. Градации фактора с максимальными значениями С(ai/bi) образуют экологический оптимум (обозначается символом «+»), остальные градации относятся к «размытой» части ниши (с символом «• »). В области экологического оптимума система рассматривается как наиболее устойчивая к внешним воздействиям [14]. Кривая,
проведенная через экологические доминанты явления, дает общую картину его ординации по данному фактору.
В итоге были получены матрицы распределения бинарных отношений системы «фактор-явление» (рис. 1–5) для признаков почвенного и фитоценотического блоков экосистем.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В почвенном блоке одним из важных признаков «явления» служит мощность гумусового профиля. Она увеличивается в направлении сверху вниз по катене. По-видимому, здесь сказывается процесс гравитационного горизонтального переноса гумуса из более крутых и высоко располо- женных склоновых геотопов в более низкие вы-положенные. Из общей тенденции выделяется лишь резкое увеличение мощности гумусового профиля в трансэлювиальном местоположении (рис. 1 а), что можно объяснить более глубокой здесь инфильтрацией почвенных растворов вглубь повышенно скелетной почвенногрунтовой толщи.
Следующий признак – мощность органоминерального горизонта А 1 . При большой и средней влажности почвы мощность данного горизонта гораздо больше, нежели в условиях сухой почвы (рис. 1 б , в ). В целом такая зависимость обусловлена, по-видимому, преобладающим тяготением лиственной древесной растительности, поставляющей наибольшее количество гумуса в почву, к местам со средними показателями почвенной влажности.
Оптимальное развитие мощности горизонта А 1 соответствует наименьшим значениям вертикального температурного градиента в почве (рис. 1 г ). Большая мощность органогенного горизонта развивается при наименьших температурных перепадах. Многие виды травянистых и кустарничковых растений плохо переносят резкое снижение температуры с глубиной почвы (рис. 1 д , е ; 2 а ), что характерные, как известно, по большей части для горных территорий.
Информационная меры сложности почвенного профиля имеет слабо выраженную тенденцию к увеличению при возрастании влажности почвы (рис. 2 б ).
Перейдем к основному объекту анализа – фи-тоценотическому блоку экосистем. Коэффициент атмосферного увлажнения Высоцкого-Иванова в регионе (по данным ст. Комсомольск-на-Амуре) составляет около 1,4, т.е. территория заповедника находится в условиях фонового избыточного увлажнения и недостатка тепла, что и определяет господство здесь длительносезонномерзлой и островной многолетней мерзлоты [18]. В условиях муссонно-континентального климата, свойственного умеренному поясу тихоокеанской окраины Евразийского материка, дефицит тепла сказывается на протяжении всего года, в том числе и в вегетационный период. Это непосредственно отражается на связях биоразнообразия лесных сообществ, и прежде всего самого древостоя, с летними термоэдафическими условиями. Более высоким температурам почвы соответствует и большее разнообразие древесных пород, причем этот рост наиболее существен в диапазоне наиболее низких температур (рис. 2 а , б ). В области минимальных температур почвы (5–10о на глубинах 30–40 см), особенно характерных для супер-аквальных маревых ландшафтов, породное разнообразие деревьев находится в пессимуме (≤ 1,4 бит), в то время как температурам 15–20о соответствует диапазон разнообразия до 2,0–2,8 бит.
|
5 Н |
Мощность гумусового профиля, см |
с г е |
1 1 |
Мощность горизонта Ai, см |
||||||||||||||||||
|
<15 |
16-25 |
26- 35 |
36-45 |
46-60 |
<9 |
10-15 |
16-21 |
34-38 |
||||||||||||||
|
3 |
• |
• |
1 |
+_____ |
||||||||||||||||||
|
Тэ |
• |
__- + |
2 |
|||||||||||||||||||
|
Та |
ч • |
3 |
||||||||||||||||||||
|
А |
4 |
• |
||||||||||||||||||||
|
Saq |
• |
• |
5 |
• |
||||||||||||||||||
|
со |
Мощность гумусового профиля, см |
о О 1_ 1— ш |
Мощность горизонта А^ см |
|||||||||||||||||||
|
<15 |
16- 25 |
26- 35 |
36-45 |
46-60 |
<9 |
10-15 |
16-21 |
34-38 |
||||||||||||||
|
1 |
1 |
+__ |
||||||||||||||||||||
|
2 |
2 |
|||||||||||||||||||||
|
3 |
3 |
|||||||||||||||||||||
|
4 |
• |
4 |
/ |
• |
||||||||||||||||||
|
5 |
5 |
• |
||||||||||||||||||||
|
5 о 1— СП |
Глубина проникновения корней травянистых растений, см |
а- го ° § Е |
Сырая надземная фитомасса травостоя, г/м3 |
|||||||||||||||||||
|
ID- 25 |
26-40 |
41-55 |
56 70 |
71 - 85 |
50 300 |
301 - 550 |
551 - 800 |
801 -1100 |
1101 -1350 |
|||||||||||||
|
0,03-0,06 |
• |
• |
5-9 |
|||||||||||||||||||
|
0,06 - 0,09 |
+ |
9,1 - 12 |
< • |
• |
||||||||||||||||||
|
0,09-0,12 |
12,1 - 16 |
• |
||||||||||||||||||||
|
0,12-0,16 |
+/ |
+ |
16,1 - 18 |
+ |
||||||||||||||||||
Рис. 1. Матрицы распределения бинарных отношений системы «фактор-явление»
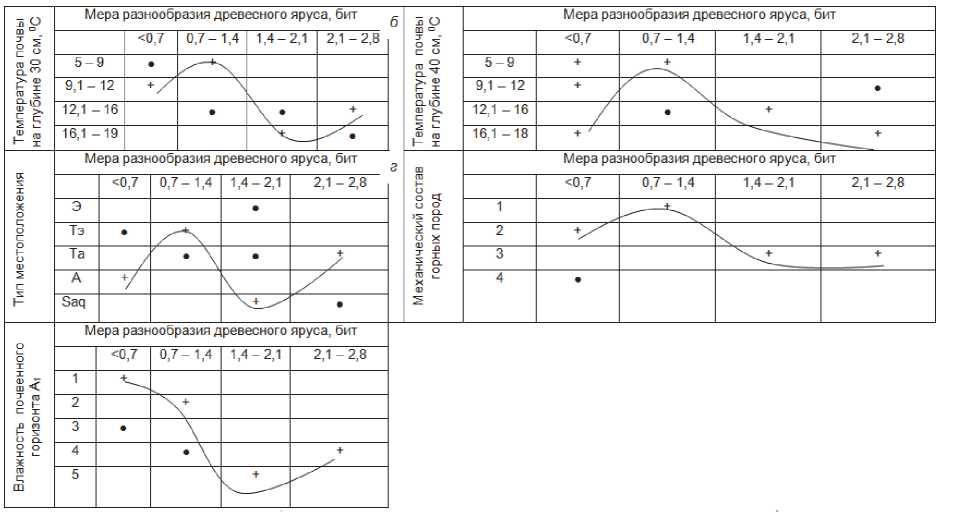
Рис. 2. Матрицы распределения бинарных отношений системы «фактор-явление»
Минимальным разнообразием древесного яруса отличаются лесные сообщества на трансэлювиальных и элювиальных местоположениях (рис. 2 в ), где температура почвы снижается с глубиной наиболее резко (рис. 3 а ) и гумусовый профиль развит сравнительно слабо (см. рис. 1 а ). Максимальное флористическое богатство древостоя свойственно лесам предгорных трансаккумулятивных и высокопойменных супераквальных геотопов низменных равнин, где почвы летом прогреваются в наибольшей степени (рис. 3 г , д ), при средней мощности гумусового профиля. Однако в слабо дренируемых заболоченных лесах
(марях), с их островной многолетней мерзлотой, существует весьма узкий набор древесных пород (ивы, березы), поскольку древесный ярус находится в пессимальных условиях произрастания.
С геоморфологической ординацией созвучна также зависимость разнообразия пород деревьев от механического состава коренных пород (рис. 2 г). Так, на песчаных и глинистых речных и озерных отложениях разнообразие древостоя минимальное. Оно начинает возрастать на отложениях глинистых сланцев и песчаников, характерных для низкогорной части территории заповедника, занимающей его большую площадь. При перехо- де же к низкогорным водоразделам, с их скальным коренными породами в виде гранодиоритов и диоритовых порфиритов, породный состав древостоя снова обедняется.
Мера разнообразия древесного яруса обратно пропорциональна влажности почвы (рис. 2 д ).
Высокая влажность, характерная для болотных и пойменных ландшафтов, резко снижает разнообразие древесной растительности, нередко до полного ее отсутствия (в случае болот).
|
1 |
Вертикальный температурный градиент почвы °С/см |
6 г |
| ч со |
Мера сложности почвенного профиля, бит |
|||||||||||||||||||
|
0,030,06 |
0,06-0,09 |
0,09-0,12 |
0,120,16 |
<1,171 |
1,172 1,536 |
1,537 - 1,901 |
1,902 -2,266 |
>2,267 |
|||||||||||||||
|
э |
• |
---- |
1 |
+ |
|||||||||||||||||||
|
Тэ |
- |
2 |
|||||||||||||||||||||
|
Та |
3 |
Х^ + |
• |
||||||||||||||||||||
|
А |
4 |
• |
• |
• |
|||||||||||||||||||
|
Saq |
■ |
- |
5 |
• |
• |
||||||||||||||||||
|
ч 5 R |
Глубина проникновения корней травянистых растений, см |
н 2 о 8 S |
Температура почвы на глубине 30 см, °C |
||||||||||||||||||||
|
10-25 |
26 40 |
41-55 |
56-70 |
71-85 |
5-9 |
9,1 - 12 |
12,1 - 16 |
16,1-19 |
|||||||||||||||
|
э |
<110 |
- |
• |
___- |
|||||||||||||||||||
|
Тэ |
+ \ |
• |
111 -220 |
+ __ |
|||||||||||||||||||
|
Та |
• / |
* |
221 - 330 |
||||||||||||||||||||
|
А |
+ / |
551 - 660 |
Х + |
||||||||||||||||||||
|
Saq |
• |
||||||||||||||||||||||
|
2 2 4 |
Температура почвы на глубине 40 см, °C |
е |
О 5 с О 3 5 |
Мера разнообразия травянистого яруса, бит |
|||||||||||||||||||
|
5-9 |
9.1-12 |
12,1-16 |
16,1-19 |
2,396 - 2,881 |
2,882 - 3,366 |
3,367-3,851 |
3,852 -4,336 |
||||||||||||||||
|
<110 |
Л |
5-9 |
4 X |
• |
|||||||||||||||||||
|
111 -220 |
+ / |
9.1 - 12 |
• х |
+ |
|||||||||||||||||||
|
221 - 330 |
//+ |
12,1 - 16 |
X • |
||||||||||||||||||||
|
551 - 660 |
16.1 - 19 |
Xt > |
|||||||||||||||||||||
|
661 - /80 |
+ |
||||||||||||||||||||||
Рис. 3. Матрицы распределения бинарных отношений системы «фактор-явление»
|
1 = 5 ГО |
Мера разнообразия древесного яруса, бит |
б |
1 4 h 5 го -О |
Сырая надземная фитомасса травостоя, г/м3 |
||||||||||||||||||||||
|
<0,7 |
0,71,4 |
1,42,1 |
2,1 2,8 |
г |
50 - 300 |
301 - 550 |
551 -800 |
801 -1100 |
||||||||||||||||||
|
2,396 -2,881 |
10-25 |
• |
||||||||||||||||||||||||
|
2,882 - 3,366 |
26-40 |
+ |
||||||||||||||||||||||||
|
3,367 - 3,851 |
41 -55 |
|||||||||||||||||||||||||
|
3.852 - 4.336 |
56-70 |
* у |
• |
|||||||||||||||||||||||
|
71-85 |
||||||||||||||||||||||||||
|
5 О со ° 1 Е н £ |
Глубина проникновения корней травянистых растений, см |
Н |
Ценотические группы |
|||||||||||||||||||||||
|
10 25 |
26 40 |
41 55 |
56 70 |
71 - 85 |
МСМ |
ЕПл |
ЛиШЛл |
Мл |
Шл |
Сл |
||||||||||||||||
|
5-9 |
• |
+ |
э |
• |
• |
* |
• |
|||||||||||||||||||
|
9,1 - 12 |
+ / |
• |
Тэ |
|||||||||||||||||||||||
|
12,1 - 16 |
• |
- |
Та |
+ |
• |
• |
||||||||||||||||||||
|
16.1 - 18 |
+ / |
Х + / |
А |
|||||||||||||||||||||||
|
Saq |
||||||||||||||||||||||||||
|
8 5 1 8 > |
Ценотические группы |
е |
И 1 ° ч CD |
Ценотические группы |
||||||||||||||||||||||
|
МСМ |
ЕПл |
ЛиШЛл |
Мл |
Шл |
СЛ |
МСМ |
ЕПл |
ЛиШЛл |
Мл |
Шл |
Сл |
|||||||||||||||
|
1 |
• |
1 |
X |
|||||||||||||||||||||||
|
2 |
+ |
2 |
+ X |
|||||||||||||||||||||||
|
3 |
/ • |
3 |
"х4 |
|||||||||||||||||||||||
|
4 |
\ + у |
х^ |
4 |
+ X |
||||||||||||||||||||||
|
5 |
||||||||||||||||||||||||||
Рис. 4. Матрицы распределения бинарных отношений системы «фактор-явление»
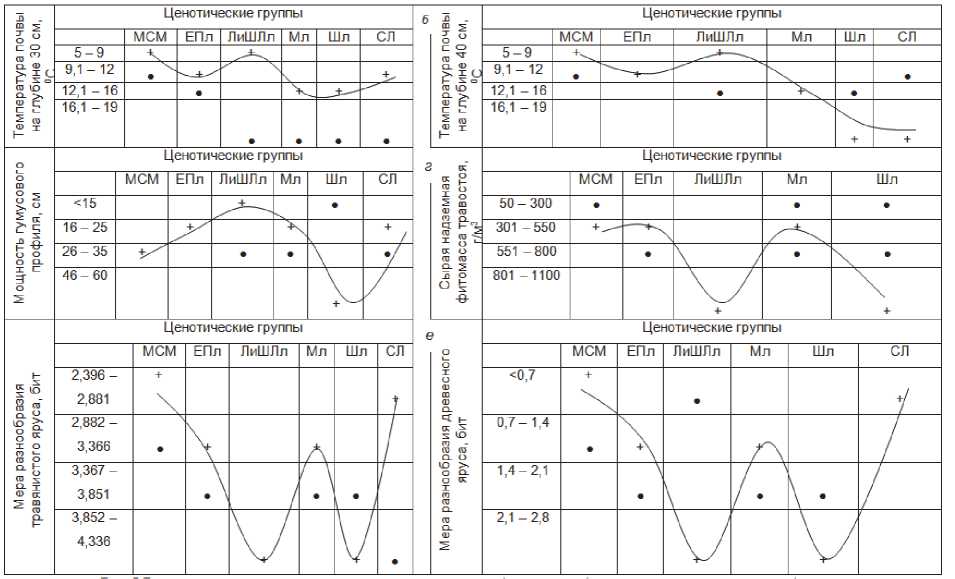
Рис. 5. Матрицы распределения бинарных отношений системы «фактор-явление»
Флористическое разнообразие травянокустарничкового яруса довольно слабо коррелирует с температурой почвы (рис. 3 е ). В целом можно отметить лишь некоторую тенденцию к увеличению богатства флоры с ростом температуры почвы. Соответственно относительно высокие и средние температуры создают больший спектр разнообразия травяно-кустарничковой растительности. Это свойственно лесам, находящимся у подножий и в нижних частях горных склонов – на абс. высотах до 220 м (рис. 3 г , д ).
Отметим далее прямо пропорциональную взаимосвязь мер разнообразия древесного и травяно-кустарничкового ярусов (рис. 4 а ). Скорее всего, здесь идет речь об их практически идентичной зависимости от определенного набора внешних факторов, главные из которых – тип местоположения, влажность почвы, экспозиция склона (если речь идет о возвышенностях). Сказывается также привязанность некоторых видов травянистых и кустарничков к особым породам деревьев, с условиями создаваемой ими затененности.
Корни травянистых растений достигают наименьшей глубины в элювиальных геотопах (рис. 2 в ), что является следствием близко залегающей здесь холодной материнской породы (рис. 2 г , д ). При смене же типа местоположения с элювиального на трансэлювиальный глубина проникновения корней травянистых начинает возрастать. Максимальных значений она достигает в мягких и наиболее теплых аллювиальных пойменных и болотных почвах супераквальных местоположений.
Чувствительность травянистого покрова к температуре почвы прослеживается также по влиянию последней на сырую надземную фитомассу травостоя. Последняя достигает максимума в условиях наиболее теплых почв (рис. 1 е ). В то же время этот максимум свойствен среднему диапазону глубины проникновения корней, а минимум массы наблюдается как при малых, так и при больших мощностях корнеобитаемого слоя почвы (рис. 4 б ). В целом же можно говорить об обратно пропорциональной связи зеленой массы травостоя с мощностью корнеобитаемого слоя почвы. Тем самым подтверждается известное положение о том, что с ухудшением внешних условий развитие надземных фракций травянистых фитоценозов ослабевает, а подземных – увеличивается.
Проследим зависимость в целом древесных ценотических группировок от геоморфологических условий (рис. 4 в, г). Осоково-сфагновая марь и мелколиственные леса, в основном березняки и осинники, тяготеют к переувлажненным супераквальным геотопам. Широколиственные леса и скалистые луга достигают своего экологического оптимума на крутых склонах, особенно солнцепечных, где они получают максимум солнечной радиации. Последнее, вместе с интенсивным стоком оказывает определенное осушающее воздействие на почву, что создает благоприятные условия для развития указанных биоценозов (рис. 4 е). Хвойные (пихтово-еловые и лиственничные) леса приурочены к трансаккумулятивному и аккумулятивному типам местоположений, где имеет место обильное увлажнение за счет стекающих сверху натечных вод, способствующих образованию подзолистого почвенного горизонта.
Интересным представляется влияние термических свойств почвы (рис. 5 а , б ) на тип фитоценоза. Своего экологического оптимума еловопихтовые и лиственничные леса, а также болотные сообщества и скалистые луга достигают при диапазоне температуры средних горизонтов почвы 5–12 0С. При этом хвойные леса толерантны и к более высоким температурным показателям – 12–19 0С, в то время как для произрастания широколиственных лесов это диапазон оптимальных температур.
Различные типы фитоценозов имеют свои оптимальные мощности гумусового профиля почвы (рис. 5 в). Максимального развития гумусовый профиль достигает на осоково-сфагновой мари, где гумус представлен по существу торфяной массой. В широколиственных лесах присутствие гумуса распространяется в почве на значительную глубину, что связано с мощным листовым опадом и оптимальным развитием сырой надземной фитомассы травостоя (рис. 5 г). Наименее развит гумусовый профиль во вторичных мелколиственных, хвойно-широколиственных и хвойных лесах. В последних образование гумуса затрудняется медленным разложением хвойного опада. Укороченный гумусовый профиль свойствен скалистым лугам, где низкая фитомасса травостоя связана с близким залеганием и выходами коренных пород.
Таблица . Качественные геокомпонентные признаки и градация их состояний в баллах
|
№ |
Геокомпонентные признаки |
Баллы |
||||
|
1 |
2 |
3 |
4 |
5 |
||
|
1 |
Тип локального местоположения (геотоп)* |
Э |
ТЭ |
ТА |
А |
Saq |
|
2 |
Механический состав почвообразующих пород** |
Га, П, С, Гл |
П, Гл |
ГлС, А, Пк |
ГД, Д, ДП |
– |
|
3 |
Вертикальный температурный градиент, ВТГ, 0 С/см |
0,03–0,06 |
0,06–0,09 |
0,09–0,12 |
0,12–0,16 |
– |
|
4 |
Состояние влажности горизонта А 1 |
мокрое |
сырое |
влажное |
свежее |
сухое |
Примечание: *Э – элювиальное, ТЭ – трансэлювиальное, ТА - трансаккумулятивное, А – аккумулятивное, Saq – супер-аквальное. **Га – галечники, П – песок, С – супесь, Гл – глина, ГлС – глинистые сланцы, А – алевролиты, Пк – песчаники, ГД – гранодиориты, Д – диориты, ДП – диоритовые порфириты.
Наибольшего флористического разнообразия травяно-кустарничко-вый ярус достигает в широколиственных и хвойно-широколиственных лесах, почвы которых обеспечены гумусом. В еловопихтовых и вторичных мелколиственных лесах разнообразие видов трав резко снижается, достигая своего минимума на болотах и скалистых лугах (рис. 5 д ). Для хвойных лесов главной причиной этого является обеднение почвы гумусом из-за опада хвои, а для мелколиственных – песчаные отложения. Минимальное разнообразие травостоя на болотах и лугах объясняется узким набором видов растений, способных произрастать в подобных специфических условиях. Для открытых для скалистых лугов – это повышенная солнечная радиация, достигающая поверхности почвы, и частые выходы коренных пород, для болот – избыточное увлажнение и повышенная кислотность почвы.
Видовое разнообразие деревьев наиболее значительно не только в широколиственных лесах, но в елово-пихтовых, где в массе фоновых древесных видов встречаются экземпляры черемухи Маака, сосны корейской, рябины амурской, некоторые виды дальневосточных берез и др. (рис. 5 е). Подобное явление подтверждает экотонный характер территории Комсомольского заповедника: к доминирующим представителям бореальной охотско-камчатской флоры здесь примыкают южные суббореальные элементы флоры маньчжурской. Минимальное флористическое разнообразие деревьев, вплоть до его полного отсутствия, свойственно болотам и скалистым лугам.
ЗАКЛЮЧЕНИЕ
Рассмотренные признаки почвенного и фито-ценотического геокомпонентных блоков бореальных и суббореальных экосистем Нижнего Приамурья достаточно восприимчивы к типу местоположения и гидротермическим условиям почвы. При этом критические показатели последних обусловливают те или иные состояния почвенно-фитоценоти-ческих признаков. Так, значения мощности гумусового профиля и глубины проникновения корней травянистых растений максимальны в супераквальных местоположениях, для которых характерна сравнительно быстрое падение температуры почвы с глубиной, и минимальны – на элювиальных геотопах, с более низкими температурными градиентами. Как продуктивность, так и разнообразие травостоя напрямую зависят от температуры почвы.
Мера разнообразия древесного яруса понижается с возрастанием влажности и снижением температуры почвы. Данный показатель довольно однозначно определяется также механическим составом пород: на аллювиальных отложениях низменных равнин древостой гораздо менее разнообразен, нежели на гранодиоритах, песчаниках и брекчиях в низкогорной части заповедника.
Аналогичные связи прослеживаются и в факторной ординации ценотических группировок лесов и болот. Переувлажненные супераквальные геотопы на галечниках, песках и глинах, а также отвечающие им низкие температуры почвы обусловливают развитие болотных экосистем и мелколиственны лесов. К трансэлювиальным местоположениям на гранодиоритах и диоритах при пониженной температуре почвы тяготеют пихтово-еловые леса, а при более высокой температуре на склонах южной экспозицией – скалистые луга и широколиственные леса. Наконец, на аккумулятивных геотопах с глубокосезонномерзлой почвой распространены лиственничники, которые толерантны к низкой температуре почвы.
Таким образом, можно предположить, что при значительных естественных (климатических) или антропогенных изменениях гидротермических показателей почвы последуют изменения в почвообразовательных процессах, которые, в конечном счете, скажутся на видовом составе растительности и ее продуктивности, а следовательно и на состоянии экосистем в целом. Выявленные вполне определенные межкомпонентные связи гео(эко)систем подтверждают повышенную чувствительность лесных сообществ экотонной территории Комсомольского заповедника к внешним воздействиям, что предопределяет неизбежность возникновение многочисленных проблем охраны ландшафтов и разработки научных подходов к их разрешению.
Список литературы Факторная ординация лесных экосистем на зональной границе (на примере Комсомольского-на-Амуре заповедника)
- Беручашвили Н.Л., Жучкова В.К. Методы комплексных физико-географических исследований. М.: Изд-во МГУ, 1997. 319 с.
- Герасимов И.П. Экологические проблемы в прошлой, настоящей и будущей географии Мира. М.: Наука, 1985. 247 с.
- Глазовская М.А. Геохимические основы типологии и методики исследований природных ландшафтов. М.: Изд-во МГУ, 1964. 230 с.
- Дьяконов, К.Н., Касимов Н.С., Тикунов В.С. Современные методы географических исследований. М.: Просвещение:, 1996. 207 с.
- Залетаев А.С. Экотонные экосистемы как географическое явление и проблемы экотонизации биосферы//Современные проблемы географии экосистем (Тезисы докл. Всесоюз. совещ.). М.: Ин-т географии АН СССР, 1984. С. 53-55.
- Исаченко А.Г. Ландшафты СССР. Л.: Изд-во ЛГУ, 1985. 320 с.
- Кастлер Г. Азбука теории информации//Теория информации в биологии. М.: Изд-во иностр. лит., 1960. С. 9-53.
- Коломыц Э.Г. Ландшафтные исследования в переходных зонах (методологический аспект). М.: Наука, 1987. 117 с.
- Коломыц Э.Г. Бореальный экотон и географическая зональность: Атлас-монография. М.: Наука, 2005. 390 с.
- Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
- Крауклис А.А. Особенности географических градаций топического порядка//Топологические аспекты учения о геосистемах. Новосибирск: Наука, 1974. С. 87-137.
- Мильков Ф.Н. Физическая география. Учение о ландшафте и географическая зональность. Воронеж: Изд-во Воронеж. ун-та, 1986. 327 с.
- Программа и методика биогеоценологических исследований/Отв. ред. Н.В. Дылис. М.: Наука, 1974. 403 с.
- Пузаченко Ю.Г., Скулкин В.С. Структура растительности лесной зоны СССР. М.: Наука, 1981. 275 с.
- Раменский Л.Г. Избранные работы. Проблемы и методы изучения растительного покрова. Л.: Наука, 1971. 334 с.
- Сочава В.Б. Растительный покров на тематических картах. Новосибирск: Наука, 1979. 189 с.
- Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
- Южная часть Дальнего Востока. Природные условия и естественные ресурсы СССР/Отв. ред. В.В. Никольская, А.С. Хоментовский. М.: Наука, 1969. 422 с.
