Фауна и эффекты гомогенизации видового состава зоопланктона устьевой области малого притока равнинного водохранилища в аномально жаркий период

Автор: Болотов Сергей Эдуардович, Айрапетян Армине Овсеповна, Крылов Александр Витальевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 5 т.16, 2014 года.
Бесплатный доступ
Приведены результаты изучения фауны зоопланктона водной системы малого притока, его устьевой области и водохранилища, а также дана оценка изменений видового состава зоопланктона устьевой области притока в условиях погодно-климатических аномалий. Показано, что в устьевой области притока, по сравнению со смежными участками реки и водохранилища, происходит увеличение таксономического богатства, систематического разнообразия и таксономической сложности зоопланктона. Под влиянием погодных термических аномалий нарушается фоновая структура сходства видового состава зоопланктона гидроэкологических зон устьевой области, снижается их фаунистическое своеобразие и наблюдается биотическая гомогенизации фауны зоопланктона.
Фауна, зоопланктон, река, водохранилище, устьевая область, температура воды, аномальные климатические условия, биотическая гомогенизация
Короткий адрес: https://sciup.org/148203307
IDR: 148203307 | УДК: 574.
Fauna and homogenization effects of zooplankton species composition mouth area of small tributary of a flatland reservoir in abnormally hot period
The research results of zooplankton fauna of the water system of small tributary, his mouth area and reservoirs, as well as an assessment of changes in the species composition of zooplankton in the mouth area inflow in the conditions of climatic anomalies. It is shown that in the mouth area of the inflow, compared with adjacent portions of rivers and reservoirs, an increase in taxonomic richness, systematic diversity and taxonomic complexity of zooplankton. Under the influence of thermal anomalies weather disrupted background structure similarity of zooplankton species composition of hydroecological zones of mouth area, disrupted them faunal specificity and observed biotic homogenization of zooplankton fauna.
Текст научной статьи Фауна и эффекты гомогенизации видового состава зоопланктона устьевой области малого притока равнинного водохранилища в аномально жаркий период
Устьевые области малых и средних притоков равнинных водохранилищ – протяженные (от 2 до > 50 км [7]) участки, которые по продольному профилю характеризуются неоднородностью водных масс [3,6].
Сведения о гидроэкологическом режиме устьевых областей притоков равнинных водохранилищ ограничены, а имеющиеся данные о развитии зоопланктона этих биотопов противоречивы.
Изучение физико-химических параметров водных масс устьевой области р. Ильдь позволило провести ее районирование и, по аналогии с районированием устьев притоков морей и крупных озер [5], выделить 3 основные гидроэкологи-ческие зоны: IIA – переходная притока с преобладанием речных вод, IIB – фронтальная, характеризующаяся значительными вертикальными градиентами с отчетливым расслоением более минерализованных речных и опресненных водохрани-лищных вод, и IIC – переходная приемника с преобладанием водных масс водохранилища.
В рамках изучения биологического режима устьевой области р. Ильдь получены первые сведения о структуре и количественной динамике сообществ зоопланктона [2], описаны экологические корреляции с приоритетными факторами среды, в том числе в период термических аномалий жарких лет [1].
Однако специальных работ, обобщающих ретроспективные и современные данные о фауни-
стическом разнообразии зоопланктона устьевой области притока в сравнении с граничными экосистемами реки и водохранилища, особенно, в условиях долговременных погодных аномалий ранее не проводилось.
Цель работы – охарактеризовать таксономическое разнообразие фауны зоопланктона устьевой области малого притока Рыбинского водохранилища и оценить ее изменения в условиях погодноклиматических аномалий жарких лет.
МАТЕРИАЛЫ И МЕТОДЫ
Сборы зоопланктона проводили 1–2 раза в месяц с мая по октябрь 2009-2011 гг. в зоне свободного течения реки Ильдь (ст. 1-2), ее устьевой области – переходной зоне притока (ст. 2А, районирована и обследована только в 2011 г.), фронтальной (ст. 3-4,4А) и переходной зоне приемника (ст. 5-7) и Волжском плесе Рыбинского водохранилища (ст. 8).
Зоопланктон собирали на медиали: на мелководных участках ведром, на глубоководных — планктобатометром объемом 5 л в столбе воды от поверхности до дна. Через газ с размером ячеи 64 мкм процеживали 20–60 л воды, пробы фиксировали 4%-ным раствором формальдегида. Камеральную обработку проб проводили по стандартной методике [4].
Анализ данных таксономического состава зоопланктона выполнен с применением запатентованной компьютерной программы «FW-Zooplankton» , разработанной в Лаборатории экологии водных беспозвоночных ИБВВ РАН.
В качестве формальных показателей сложности таксономической структуры сообществ применяли индекс средней таксономической отличительности (AvTD Δ+), характеризующий таксономическое разнообразие зоопланктона, и индекс вариабельности таксономической отличительно- сти (VarTD Λ+), отражающий меру сложности иерархического древа таксоцена.
Классификация сообществ зоопланктона выполнена путем иерархического кластерного анализа методом Варда с реализацией элементов дисперсионного анализа. Надежность кластерных решений оценивали процедурой бутстрепа для 999 пермутаций: кластеры признавали значимыми при уровне бутстреп-поддержки ≥ 70. Многомерную ординацию сообществ осуществляли методами неметрического шкалирования (nMDS).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Обобщение оригинальных данных и неопубликованных архивных материалов позволило установить, что зоопланктон водной системы малого притока, – реки Ильдь, и Волжского плеса Рыбинского водохранилища сложен достаточно разнообразным составом и включает в себя 238 видов (в том числе внутривидовых форм – 258), из которых коловраток – 144 (60,5%), ветвистоусых – 65 (27,3%) и веслоногих ракообразных – 29 (12,2%) видов (табл. 1).
|
Таблица 1. Распределение числа видов в семействах локальных фаун зоопланктона водной системы р. Ильдь – малого притока равнинного водохранилища |
|||||||||||
|
Семейство |
Станция |
Всего видов |
|||||||||
|
1 |
2 |
2A |
3 |
4 |
4A |
5 |
6 |
7 |
8 |
||
|
ROTIFERA |
|||||||||||
|
Asplanchnidae |
0 |
1 |
1 |
5 |
5 |
5 |
5 |
5 |
4 |
3 |
5 |
|
Bdelloidea |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
Brachionidae |
14 |
12 |
5 |
15 |
11 |
11 |
10 |
12 |
13 |
9 |
19 |
|
Collothecidae |
1 |
1 |
0 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
Conochilidae |
2 |
2 |
1 |
4 |
3 |
2 |
3 |
4 |
4 |
3 |
5 |
|
Dicranophoridae |
3 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
Euchlanidae |
5 |
6 |
3 |
5 |
3 |
2 |
1 |
4 |
4 |
3 |
8 |
|
Flosculariidae |
0 |
0 |
0 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
|
Gastropodidae |
2 |
2 |
0 |
4 |
2 |
1 |
4 |
3 |
3 |
0 |
4 |
|
Hexarthridae |
0 |
0 |
0 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
Lecanidae |
11 |
9 |
5 |
8 |
3 |
3 |
4 |
4 |
4 |
4 |
14 |
|
Lepadellidae |
3 |
3 |
1 |
4 |
1 |
0 |
0 |
0 |
0 |
1 |
5 |
|
Lindiidae |
2 |
2 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
2 |
|
Mytilinidae |
5 |
4 |
1 |
4 |
2 |
1 |
0 |
2 |
3 |
1 |
5 |
|
Notommatidae |
5 |
7 |
1 |
4 |
1 |
1 |
1 |
1 |
2 |
1 |
12 |
|
Philodinidae |
3 |
3 |
0 |
6 |
3 |
2 |
3 |
3 |
2 |
1 |
7 |
|
Ploima gen. sp. |
1 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
|
Proalidae |
1 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
|
Scaridiidae |
0 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
|
Synchaetidae |
7 |
8 |
6 |
11 |
10 |
10 |
11 |
11 |
12 |
14 |
17 |
|
Testudinellidae |
3 |
2 |
2 |
3 |
3 |
1 |
2 |
4 |
3 |
2 |
5 |
|
Trichocercidae |
7 |
10 |
5 |
13 |
9 |
13 |
10 |
8 |
10 |
11 |
19 |
|
Trichotriidae |
2 |
2 |
2 |
2 |
1 |
2 |
2 |
2 |
1 |
2 |
3 |
|
Trochosphaeridae |
2 |
1 |
1 |
3 |
3 |
3 |
3 |
4 |
3 |
2 |
4 |
|
CLADOCERA |
|||||||||||
|
Bosminidae |
4 |
3 |
1 |
4 |
4 |
4 |
4 |
4 |
4 |
4 |
4 |
|
Cercopagididae |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
1 |
|
Chydoridae |
14 |
16 |
11 |
12 |
10 |
7 |
10 |
13 |
8 |
8 |
26 |
|
Daphniidae |
8 |
9 |
6 |
10 |
10 |
8 |
10 |
12 |
6 |
8 |
18 |
|
Eurycercidae |
1 |
1 |
0 |
2 |
1 |
0 |
1 |
1 |
0 |
0 |
2 |
|
Ilyocryptidae |
0 |
1 |
1 |
1 |
1 |
1 |
0 |
0 |
0 |
0 |
1 |
|
Leptodoridae |
0 |
0 |
0 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
Macrothricidae |
1 |
1 |
0 |
1 |
0 |
0 |
1 |
1 |
0 |
0 |
5 |
|
Moinidae |
0 |
1 |
1 |
2 |
1 |
2 |
1 |
1 |
1 |
0 |
2 |
|
Polyphemidae |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
|
Sididae |
2 |
1 |
3 |
4 |
4 |
4 |
4 |
4 |
4 |
4 |
4 |
|
COPEPODA |
|||||||||||
|
Cyclopidae |
6 |
9 |
6 |
11 |
13 |
11 |
14 |
17 |
11 |
15 |
23 |
|
Diaptomidae |
0 |
0 |
1 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
|
Temoridae |
0 |
0 |
1 |
3 |
1 |
2 |
3 |
3 |
2 |
3 |
4 |
|
ВСЕГО ВИДОВ: |
238 |
||||||||||
Среди коловраток отмечены представители 24 семейств, из которых наибольшим богатством отличаются сем. Brachionidae – 31 вид и формы, сем. Trichocercidae – 21 вид и формы, сем. Synchaetidae – 17 видов, сем. Lecanidae – 15 видов и форм и сем. Notommatidae – 13 видов и форм. Ветвистоусые ракообразные представлены 11 семействами. Таксономическое богатство Cladocera главным образом формируют представители сем. Chydoridae (26 видов) и сем. Daphniidae (18 ви- дов); в меньшей степени – представители сем. Macrothricidae (5 видов), сем. Bosminidae (4 вида) и сем. Sididae (4 вида). Фауну веслоногих ракообразных слагают представители подсемейств Cyc-lopinae (17 видов) и Eucyclopinae (6 видов), сем. Temoridae (4 вид), сем. Diaptomidae (2 вида).
250 Общий видовой список - 238 видов
Cl V) Z
□D 150 о cl s CD
О 100
о s
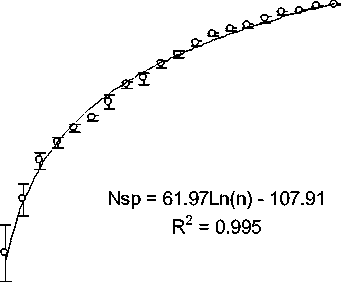
0 “I----------1-----------1----------i-----------1----------1-----------i----------i-----------i----------1-----------1----------i-----------1----------i-----------i----------1-----------i----------1-----------1----------1-----------1----------1
0 50 100 150 200
Количество проб (n)
Рис 1. Кривая зависимости числа обнаруженных видов зоопланктона в фаунистическом списке от объема наблюдений (по материалам интегральных проб за 2009-2011 гг.)
Полнота выявленного видового богатства оценивается степенью насыщения кумулятивной кривой изменения обнаруженного числа видов в зависимости от количества просмотренных проб. Полученная для изученной водной системы р. Ильдь оценка полноты выявленного видового богатства зоопланктона может быть аппроксимирована логарифмической функцией с асимптотическим насыщением при 210 видах (рис. 1). Учитывая, что общий список фауны зоопланктона водной системы притока и приемника включает 238 видов, а зона плато «кривой сборщика» устанавливается на 210 видах, можно говорить о том, что видовой состав зоопланктона выявлен достаточно полно и значительного расширения фаунистического списка в ходе последующих исследований ожидать не следует.
Минимальные значения таксономического богатства, систематического разнообразия и таксономической сложности фауны зоопланктона наблюдали в Волжском плесе Рыбинского водохранилища (табл. 2). Более высокими, по сравнению с водохранилищем, значениями параметров фауны зоопланктона характеризуется зона свободного течения притока, отличающаяся значительным разнообразием речных биотопов (наличие зарослей, различия в интенсивности водообмена, прогреваемости и др.) и видимым стоковым течением (0,3-0,7 м/с), обуславливающим обогащение фауны за счет дрифта планктона с территории водосбора.
Таблица 2. Показатели таксономического богатства, систематического разнообразия и таксономической сложности фауны зоопланктона гидроэкологических зон водной системы реки Ильдь и Волжского плеса Рыбинского водохранилища
|
Показатель |
I |
IIA |
IIB |
IIC |
III |
||||||
|
1 1 |
2 |
2A |
3 |
4 |
4A |
5 |
6 |
7 |
8 |
||
|
Число видов |
153 |
67 |
171 |
155 |
107 |
||||||
|
117 |
123 |
149 |
112 |
103 |
114 |
131 |
111 |
||||
|
Число родов |
69 |
41 |
72 |
65 |
52 |
||||||
|
58 |
58 |
66 |
57 |
50 |
53 |
61 |
54 |
||||
|
Число семейств |
32 |
24 |
32 |
30 |
27 |
||||||
|
28 |
32 |
32 |
30 |
28 |
28 |
30 |
27 |
||||
|
видов в семействе |
4.78 |
2.79 |
5.34 |
5.17 |
3.96 |
||||||
|
s У CD n CD О CL О |
4.18 |
3.84 |
4.66 |
3.73 |
3.68 |
4.07 |
4.37 |
4.11 |
|||
|
родов в семействе |
2.16 |
1.71 |
2.25 |
2.17 |
1.93 |
||||||
|
2.07 |
1.81 |
2.06 |
1.90 |
1.79 |
1.89 |
2.03 |
2.00 |
||||
|
видов |
2.22 |
1.63 |
2.38 |
2.38 |
2.06 |
||||||
|
в роде |
2.02 |
2.12 |
2.26 |
1.96 |
2.06 |
2.15 |
2.15 |
2.06 |
|||
|
AvTD |
Δ+ |
96.81 |
96.67 |
97.04 |
96.74 |
96.39 |
|||||
|
96.93 96.85 |
97.36 |
97.00 |
96.55 |
96.83 |
96.94 |
96.96 |
|||||
|
VarTD Λ+ |
175.06 |
175.48 |
159.36 |
173.07 |
193.61 |
||||||
|
169.19 175.39 |
148.07 |
160.21 |
195.11 |
171.74 |
160.60 |
165.80 |
|||||
Максимальное фаунистическое богатство, систематическое разнообразие и сложность фауны зоопланктона формируется в устьевой области притока, и, главным образом, ее фронтальной зоне, где происходит контакт разнотипных водных масс реки и водохранилища, и которую по совокупности признаков (повышенное видовое богатство, развитие краевого эффекта) мы определяем как экотон [3].
Сравнительный анализ видового состава зоопланктона показывает, что из 238 обнаруженных видов общими для районированных гидроэколо-гических зон являются только 39. Это преимущественно эврибионтные, а также некоторые зарос-левые и прибрежные виды с широким ареалом распространения. Основную их долю составляют коловратки (26 видов), в меньшей степени – ракообразные, из которых Cladocera – 10, Copepoda
– 3 вида. К видам, общим для всех гидроэкологи-ческих зон, относятся коловратки Asplanchna pri-odonta , Bdelloida spp. (non det.), Brachionus angu-laris , B. diversicornis , B. quadridentatus , Cephalo-della gibba , Conochilus hippocrepis , Euchlanis dila-tata , E. incisa , Filinia longiseta , Keratella cochlearis , K. quadrata , Lecane lunaris , Mytilina ventralis , Polyarthra dolichoptera , P. longiremis , P. luminosa , P. major , Pompholyx sulcata , Synchaeta tremula , Testudinella patina , Trichocerca (D.) similis , Trichocerca capucina , T. cylindrica , T. pusil-la , Trichotria pocillum , ветвистоусые рачки – Alona rectangula , Bosmina longirostris , Ceriodaphnia pul-chella , C. quadrangula , Chydorus sphaericus , Dia-phanosoma brachyurum , Disparalona rostrata , Polyphemus pediculus , Scapholeberis mucronata , Sida crystallina , и веслоногие ракообразные – Mesocyclops leucarti , Thermocyclops crassus , T. oithonoides .
В зоне свободного течения наиболее часто встречается Euchlanis dilatata (P=75%), реже (50– 58%) — Bdelloida sp. , Testudinella patina , Tricho-tria pocillum и копеподиты Cyclopoida . Высокой встречаемостью (P=75–100%) в переходной зоне притока характеризуются коловратки Bdelloida ssp. , E. dilatata , Lecane lunaris , Testudinella patina, Trichotria pocillum, а также ракообразные Thermocyclops oithonoides и Bosmina longirostris , Ceriodaphnia pulchella Sars, Chydorus sphaericus , являющиеся индикаторами мезоэвтрофных вод. В зоопланктоне фронтальной зоны наиболее часто (P=80–95%) встречаются Trichocerca capucina , Diaphanosoma brachyurum , Bosmina coregoni , Mesocyclops leucarti , Thermocyclops oithonoides ; высока встречаемость (>70%) видов–индикаторов мезоэвтрофных вод: Filinia longiseta , Keratella cochlearis , K. quadrata , Polyarthra euryptera , Pompholyx sulcata , Bosmina longirostris и Daphnia cucullata . К наиболее часто встречающимся (P=75–100%) видам зоопланктона переходной зоны приемника относятся Polyarthra major , Bos-mina coregoni , Mesocyclops leucarti , Thermocyclops oithonoides , а также характерные для мезоэвтроф-ных вод Brachionus angularis , Filinia longiseta , Keratella cochlearis , K. quadrata , Bosmina longiro-stris , Daphnia cucullata и Thermocyclops crassus . В зоопланктоне водохранилища высокой встречаемостью (P=70–90%) отличаются Polyarthra major , Synchaeta pectinata , S. tremula , Bosmina coregoni Baird, Mesocyclops leucarti , Thermocyclops oitho-noides и индикаторы мезоэвтрофных вод Keratella quadrata , Chydorus sphaericus , Bosmina longirostris . Таким образом, в устьевой области реки происходит увеличение встречаемости видов–индикаторов процесса эвтрофирования вод.
Исследование зоопланктона устьевой области притока в 2009 г. проходило в вегетационный период, который по метеорологическим условиям практически не отличался от среднемноголетних значений, что позволило определить его как «фоновый». Вегетационный период 2010 г. по многим показателям, например, продолжительной летней жаре, атмосферной и почвенной засухе, характеризовался как «аномально жаркий», а 2011 — как «жаркий».
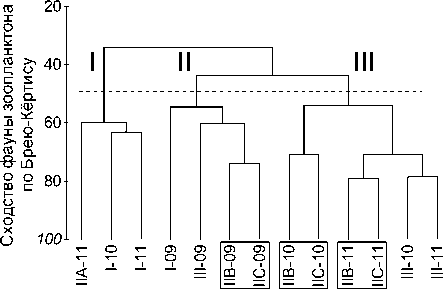
Зоопланктон водной системы притока в аномально жарких 2010 и 2011 гг. характеризовался своеобразным видовым составом, резко отличным от вегетационного периода 2009 г., соответствующего по метеорологическим условиям среднемноголетним значениям (рис. 2). При этом устьевая область притока устойчиво обособляется отдельным фаунистическим кластером.
Stress: 0.09
1-09
Фоновый год

IIC-09.
IIB-09
III
IIB-10
IIB-11 IIC-11
IIIWIIC-1O
1-10 1-11
Река в жаркие годы
IIA-11
I
Устьевая область и водохранилище в жаркие годы
Рис. 2. Дендрограмма сходства видового состава и nMDS-ординация сообществ зоопланктона гидро-экологических зон водной системы малого притока водохранилища
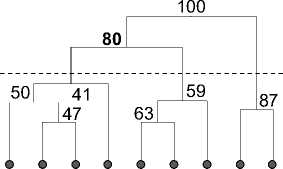
По видовому составу локальные сообщества зоопланктона фонового 2009 г. разделяются в соответствии с принадлежностью к районированным гидроэкологическим зонам (рис. 3). Так, выделяется статистически значимый (значение бут-стреп-поддержки – 100) кластер станций зоны свободного течения притока, фронтальной зоны (80) и смешанный кластер (80), объединяющий сообщества переходной зоны приемника и сходное с ними сообщество зоопланктона водохранилища.
2009 год
2010 год
2011 год
8 7 6 5 4А 4 3 2 1
61 28
43 51
ЛЛI
94 94

III+IIC
IIB I
8 7 6 5 4 4А 3 1 2
■^^^■е
III IIC+IIB IIB I
6 8 7 4 5 4А 3 2А 1 2

III+IIC+IIB IIB IIA+I
Рис. 3. Структура фаунистического сходства зоопланктона притока, его устьевой области и водохранилища в фоновом (2009) и жарких (2010, 2011) годах
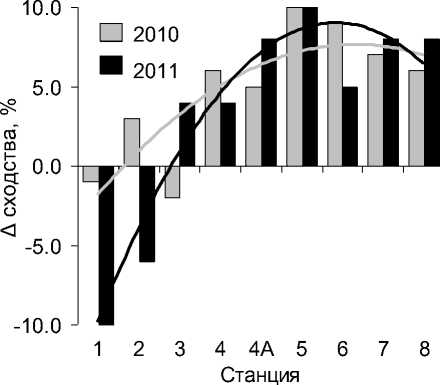
Рис. 4. Изменение средних значений сходства фауны (по Жаккару) зоопланктона реки в аномально жаркие годы по сравнению с фоновым периодом
Под влиянием сильного прогрева воды в жаркие годы по сравнению с фоновым периодом происходят изменения кластерной структуры фаунистического сходства сообществ устьевой области притока. В частности, в аномально жарком 2010 г. отдельными кластерами (значение бутстреп-поддержки кластеров – 79) обособляется сообщество зоопланктона водохранилища и смешанный кластер сообществ переходной зоны приемника и фронтальной зоны. Прогрессирующая погодная аномалия жаркого 2011 г. ведет к стиранию фаунистических различий зоопланктона водохранилища, фронтальной и переходной зоны приемника (бутстреп-поддержка кластера – 89). Значимость выделенных фаунистических кластеров для фонового и жаркого периода подтверждается также процедурой анализа группового сходства ANOSIM: R2009 = 0,712, p = 0,001; R2010 = 0,982, p = 0,001; R2011 = 0,854, p = 0,002. Таким образом, в условиях термической аномалии жарких лет нарушается фоновая структура сходства сообществ гидроэкологических зон, а их фаунистическое своеобразие стирается – проис- ходит процесс биотической гомогенизации видового состава зоопланктона [8].
Снижение водности реки и сокращение дрифта планктонных беспозвоночных с водосбора определяет развитие в зоне свободного течения притока процесса, обратного процессу гомогенизации фауны зоопланктона, а именно – ее дифференциации (рис. 4). Так, изменение среднего уровня фаунистического сходства (ΔJ) станций проточного участка реки в жаркие годы по сравнению с фоновым периодом составило в среднем +1,0% в 2010 г. и -8,0% – в 2011 г.
В устьевой области притока процесс биотической гомогенизации фауны зоопланктона наиболее выражен в переходной зоне приемника (ΔJ2010 = +8,7%, ΔJ 2011 = +7,7%), сообщества которой взаимно обогащаются видами, равномерно расселяющихся из сопредельных зон – фронтальной и водохранилища.
В целом для устьевой области процесс биотической гомогенизации фауны зоопланктона прогрессирует (ΔJ 2010 = +5,8%, ΔJ 2011 = +6,5%), а сходство между видовыми комплексами зоопланктона возрастает.
ЗАКЛЮЧЕНИЕ
На основании изложенного можно заключить, что фауна зоопланктона водной системы притока, его устьевой области и водохранилища весьма богата и включает в себя более 250 видов и форм. Наибольшее таксономическое богатство, систематическое разнообразие фауны зоопланктона и его таксономическая сложность характерны для устьевой области притока и, особенно, ее фронтальной зоны.
Погодные термические аномалии приводят к нарушению фоновой структуры сходства видового состава зоопланктона гидроэкологических зон устьевой области, снижению их фаунистического своеобразия и активизации процесса биотической гомогенизации фауны зоопланктона.
Список литературы Фауна и эффекты гомогенизации видового состава зоопланктона устьевой области малого притока равнинного водохранилища в аномально жаркий период
- Болотов С.Э., Крылов А.В., Еськов В.М., Козлова В.В., Мухортова О.В. Сравнительный анализ экологической структуры и параметров хаотической организации зоопланктона устьевой области притока равнинного водохранилища//Изв. Самар. НЦ РАН. 2014. Т. 16, № 1. С. 223-226.
- Болотов С.Э., Крылов А.В., Цветков А.И., Соколова Е.А., Поддубный С.А. Водные массы и зоопланктон зоны подпора притока Рыбинского водохранилища//Поволжский экологический журнал. 2012. № 2. С. 134-141.
- Крылов А.В., Цветков А.И., Малин М.И., Романенко А.В., Поддубный С.А., Отюкова Н.Г. Сообщества гидробионтов и физико-химические параметры устьевой области притока равнинного водохранилища//Биология внутр. вод. 2010. № 1. С. 65-75.
- Методика изучения биогеоценозов внутренних водоемов/Под ред. Ф.Д. Мордухай-Болтовского. М.: Наука, 1975. 240 с.
- Михайлов В.Н. Гидрологические процессы в устьях рек. М.: ГЕОС, 1997. 176 с.
- Рохмистров В.Л. Гидрологическая характеристика р. Солоницы в зоне подпора//Биология внутр. вод: Информ. бюл. Л.: Наука, 1973. № 23. С. 57-59.
- Рыбинское водохранилище/Отв. ред. Б. С. Кузин. Л.: Наука. Ленингр. отд-ние, 1972. 364 с.
- Rahel F.J. Homogenization of freshwater faunas//Annu. Rev. Ecol. Syst. 2002. Vol. 33. pp. 291-315.
