Фауна и сообщества донных беспозвоночных водотоков бассейна Верхней Вишеры

Автор: Паньков Н.Н., Овчанкова Н.Б.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 2, 2017 года.
Бесплатный доступ
В зообентосе бассейна Верхней Вишеры установлено 176 видов и форм. Разнообразно представлены личинки комаров-звонцов (80 видов и форм). Богаты видами ручейники (25), подёнки (23) и веснянки (20). Основу фауны (61.7%) образует группа широкоареальных видов. На долю западных и восточных палеарктов приходится 25.7 и 11.3% соответственно. Два вида (1.3%) являются эндемиками. В Верхней Вишере и её притоках - р. Хальсории, Лопье и Большой Мойве - формируются зообентоценозы с преобладанием ручейников Arctopsyche ladogensis и Apatania crymophila, веснянок Arcynopteryx compacta и подёнок Heptagenia sulphurea. Их численность составляет 1 150- 8 180 экз/м, биомасса - от 1 600 до 5 110 мг/м. В р. Ниолс, Малая Мойва и ручье Малый Лиственничный формируются зообентоценозы, ядро которых образуют ручейники Rhyacophila nubila и A. crymophila, веснянки A. compacta и подёнки H. sulphurea. Их численность составляет 800...8 010 экз/м2, биомасса - 3 980...5 600 мг/м2. В р. Муравей формируется зообентоценоз с доминированием хирономид. Его численность и биомасса достигают 34 010 экз/м2 и 30 070 мг/м2 соответственно. В ручьях формируются зообентоценозы, основу которых слагают подёнки рода Baetis, веснянки Isoperla grammatica и Amphinemura borealis, ручейники A. crymophila и Potamophylax latipennis. Численность донной фауны составляет 1 720 экз/м2 при биомассе 2 490 мг/м2.
Северный урал, водотоки, донные беспозвоночные, сообщества
Короткий адрес: https://sciup.org/147204822
IDR: 147204822 | УДК: 574.587
The fauna and benthic communities of invertebrates of watercourses of the Upper Vichera river basin
There are 176 species and forms in the zoobenthos of the Upper Vishera river basin. Larvae of chi-ronomids are the most diversely (80 species and forms). There are 25 species of caddisflies, 23 species of mayflies and 20 species of stoneflies. The basis of the fauna (61.7%) is species with broad areas. The western and the eastern Palaearctic species account for 25.7% and 11.3%. Two species (1.3%) are endemic. In the Upper Vishera and its tributaries - the Halsoria, the Lopya and the Big Moyva rivers - communities, where caddisflies Arctopsyche ladogensis and Apatania crymophila, stoneflies Arcynop-teryx compacta and mayflies Heptagenia sulphurea are dominated, are formed. Its number is varied between 1 150-8 180 ind/m2, the biomass - from 1 600 to 5 110 mg/m2. Communities, where caddisflies Rhyacophila nubila and A. crymophila, stoneflies A. compacta and mayflies H. sulphurea are dominated, are formed in the Niols river, the Small Moyva river and in the Small Listvennychnye stream. The number of zoobenthos is 800.8 010 ind/m2, the biomass - 3 980.5 600 mg/m2. Community, where larvae of chironomids are dominated, is formed in the Muravei river. The number and the biomass reach 34 010 ind/m2 and 30 070 mg/m2. Communities, where mayflies of Baetis genus, stoneflies Isoperla gram-matica, Amphinemura borealis and caddisflies A. crymophila and Potamophylax latipennis are dominated, are formed in small streams. The number of benthic fauna is 1 720 ind/itf and the biomass - 2 490 mg/m2.
Текст научной статьи Фауна и сообщества донных беспозвоночных водотоков бассейна Верхней Вишеры
Горные реки крайнего северо-востока Пермского края относятся к числу водных объектов, малоизученных в гидробиологическом отношении. Это связано, прежде всего, со слабой хозяйственной освоенно- стью этой территории, её значительной удаленностью от транспортных путей и, как следствие, недоступностью для регулярных посещений.
Первым источником, содержащим информацию о зообентоценозах водотоков бассейна Верхней Вишеры, является защищенная в 1984 г. курсовая работа студента 4-го курса биологического
факультета Пермского государственного университета А.А. Корионова, оформленная в виде научной статьи М.С. Алексевниной [Алексевнина, Корио-нов, 2006]. В ней приводятся результаты исследований донной фауны небольшого участка Вишеры и её притоков – р. Ниолс и Муравей, проведенных летом 1983 г.
В июне 1994 г., при проведении комплексных кадастровых исследований на территории Вишерского заповедника, на перекате р. Вишеры, близ устья р. Хальсории, была отобрана единственная проба зообентоса. Результаты её обработки (до уровня групп) опубликованы в монографии Е.А. Зиновьева и В.В. Семёнова [2013].
В июле и августе 2007 г. в бассейне Верхней Вишеры работали две научные экспедиции, в которых приняли участие сотрудники Вишерского заповедника и учёные Пермского государственного университета. Результатом их деятельности явились статьи А.Б. Крашенинникова с соавторами [Крашенинников и др., 2008; Крашенинников, Макарченко, 2009а, б]. В этих публикациях приводятся сведения об амфибио-тических насекомых Вишерского заповедника – комарах-звонцах, подёнках, веснянках и ручейниках, анализируется зоогеографическая структура фауны, даётся оценка степени её изученности, подвергаются ревизии ранее опубликованные списки.
Другим итогом работы научных экспедиций 2007 г. явились публикации, освещающие структуру и количественные параметры донной фауны родников в долине р. Верхней Вишеры между устьями р. Муравей и Мойва [Паньков и др., 2008; Паньков, Крашенинников, 2012].
К настоящему времени таксономический состав донных беспозвоночных бассейна Верхней Вишеры изучен сравнительно неплохо. Однако этого нельзя сказать о структуре населения и параметрах развития их сообществ. Между тем, учитывая первозданный облик природы этой заповедной территории, её ценность как эталона естественного состояния экосистем, означенные вопросы представляют исключительный интерес с точки зрения и теории, и практики.
Природные условия
Вишера – один из горных (левых) притоков р. Камы, берет начало близ северо-восточной границы Пермского края, на западном склоне хр. Поясо-вый камень (среди отрогов вершины Поримонгит-Ур) на высоте около 1000 м над ур. м., и впадает в главную реку на 958 км от ее устья. Длина Више- ры составляет 415 км, средняя высота и площадь водосбора – 317 м и 31200 км2 соответственно [Комлев, Черных, 1984].
Общепринятого членения Вишеры на участки не существует. В настоящей работе под верхним течением, или Верхней Вишерой, мы будем понимать участок реки от истока до устья р. Лыпьи.
Бассейн Верхней Вишеры, по сравнению с другими территориями Пермского края, отличается суровостью природных условий. Его территория представляет собой горный ландшафт с амплитудами высот свыше 1000 м. Отдельные вершины достигают абсолютных отметок 1331 м (Ишерим), 1351 м (Муравьиный камень) и 1469 м (Тулым-ский камень). Донья и склоны долин покрыты лесами, представленными типичной горной еловопихтовой тайгой с примесью сосны сибирской и лиственницы. Многие горные массивы поднимаются выше границы лесопроизрастания (600– 700 м), их крутые склоны покрыты обширными осыпями, а на вершинах развиты альпийские луга и горные тундры.
По сравнению с другими районами Пермского края, климат Верхней Вишеры более холодный и влажный [Шкляев, Балков, 1963]. Здесь самая длинная, суровая и многоснежная зима, короткое, холодное и дождливое лето с частыми туманами. Годовые суммы осадков в долинах составляют 850–950 мм, на хребтах и вершинах гор достигая 1300–1600 мм. Средняя годовая температура воздуха, даже в межгорных и речных долинах, не превышает –2.0…– 4.0°С, понижаясь на 0.5…0.7°С с увеличением высоты местности на каждые 100 м [там же].
Бассейн Верхней Вишеры является областью интенсивного питания водотоков. Здесь, наряду с повышенными осадками и малым испарением, имеются особо благоприятные условия для стока воды – значительные уклоны земной поверхности и водоупорность слагающих ее пород – палеозойских известняков, серицитовых и кварцитовых сланцев [Химическая география…, 1967]. В силу этого, бассейн Верхней Вишеры отличается рекордно высокими модулями стока, достигающими 23 л/сек с 1 км2 [Комлев, Черных, 1984] и располагает развитой речной сетью, густота которой превышает 0.6 м/км2 [там же]. Для водотоков характерны большие уклоны, множество водопадов, порогов и перекатов. Течение рек бурное.
Среди особенностей гидрологического режима Верхней Вишеры следует отметить затяжное половодье, связанное с постепенным стаиванием снегов в горах, и частые летне-осенние дождевые паводки [Ресурсы…, 1967].
Специфика геологического строения бассейна Верхней Вишеры, большое количество атмосферных осадков и малая испаряемость находят отра- жение в гидрохимическом режиме поверхностных вод. Они имеют низкую минерализацию (0.03–0.06 г/л) и гидрокарбонатно-кремнеземный состав, характерный для ультрапресных фаций [Химическая география…, 1967].
Материал и методы
В основу настоящей публикации положены результаты обработки 43 количественных проб зообентоса, отобранных на участке р. Верхней Вишеры протяженностью около 60 км (от устья р. Халь-сории до устья р. Лыпьи), а также в её некоторых притоках летом 1995, 2000, 2007 и 2016 гг. (рисунок).
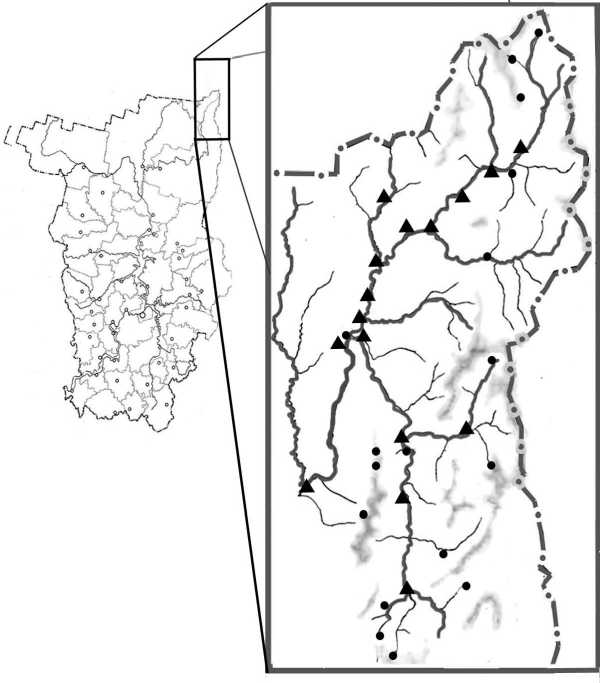
Район исследований
Треугольниками обозначены места отбора серий гидробиологических проб и имаго амфибиоти-ческих насекомых, кружками – пункты сбора имаго амфибиотических насекомых
Для наиболее полного выявления состава гидрофауны осуществлялись качественные сборы. Животные собирались вручную и пинцетом с извлеченных из воды камней, коряг, затопленных ветвей и стволов деревьев. Качественным обследо- ваниям подвергались периферийные биотопы – прибрежные мелководья, ключи, родники, старицы. Всего собрано и обработано 26 качественных проб донной фауны.
С целью уточнения таксономического состава некоторых групп амфибиотических насекомых (поденок, веснянок и ручейников) отлавливались крылатые особи. Всего обработано 54 сбора имаго, просмотрено и идентифицировано до вида 632 экземпляра.
Отбор проб, их обработка и анализ материала выполнялись по предложенной нами [Паньков, 2000] методике.
Идентификация животных выполнялась по определителям О.А. Черновой [1964], Л.А. Жильцовой [1964, 2003], О.Л. Качаловой [1987], Н.Ю. Клюге [1997], В.Д. Иванова с соавторами [2001], В.А. Тесленко и Л.А. Жильцовой [2009]. Малощетинковые черви определены доцентом Е.В. Пресновой, личинки комаров-звонцов – доцентом И.В. Поздеевым.
В полевых исследованиях приняли участие Е.Ю. Крайнев, А.Б. Крашенинников, И.И. Кропачев, Н.С. Мазура, Н.В. Панькова, В.В. Семёнов, О.С. Старова, А.В. Югов. Мы выражаем им свою искреннюю признательность.
Таксономический состав
В составе фауны донных беспозвоночных бассейна Верхней Вишеры, согласно оригинальным и литературным данным, установлено 176 видов и форм (табл. 1).
Наиболее разнообразно представлены личинки комаров-звонцов, насчитывающие здесь 80 видов и форм. Богаты видами ручейники (25), подёнки (23) и веснянки (20). Среди прочих беспозвоночных зарегистрированы нехирономидные двукрылые (10), малощетинковые черви (6), жесткокрылые (5), двустворчатые (2) и брюхоногие (1) моллюски, а также турбеллярии (1), водяные клещи (1), стрекозы (1) и вислокрылки (1).
Таблица 1
Таксономический состав и географическое распространение донных беспозвоночных бассейна Верхней Вишеры
|
Таксон |
Источник |
Географическое распространение |
|
КЛАСС TURBELLARIA |
||
|
Polycelis tenuis Ijima, 1884 |
1 |
Палеарктика |
|
КЛАСС OLIGOCHAETA |
||
|
Семейство Enchytraeidae |
||
|
Enchytraeus sp . |
1 |
|
|
Семейство Lumbriculidae |
||
|
Lumbriculus variegatus (O.F. Mueller, 1773) |
1, 2 |
Голарктика, Палеотропис |
|
Stylodrilus heringianus Claparede, 1862 |
2 |
Европа |
Продолжение табл. 1
|
Таксон |
Источник |
Географическое распространение |
|
Семейство Naididae |
||
|
Nais barbata O.F. Mueller, 1773 |
2 |
Голарктика |
|
Slavina appendiculata (d'Udekem, 1855) |
2 |
Голарктика, Палео- и Неотропис |
|
Семейство Tubificidae |
||
|
Limnodrilus sp. |
1 |
|
|
КЛАСС BIVALVIA |
||
|
Parasphaerium rectidens (Starobogatov et Streletzkaja, 1967) |
1 |
Сибирь, север Европы |
|
Pisidium amnicum (Mueller, 1774) |
2 |
Палеарктика |
|
КЛАСС GASTROPODA |
||
|
Ancylus fluviatilis Mueller, 1774 |
1, 2 |
Европа, Западная Сибирь |
|
КЛАСС ARACHNIDA |
||
|
Hydracarina indet. |
1 |
|
|
КЛАСС INSECTA |
||
|
ОТРЯД EPHEMEROPTERA |
||
|
Семейство Siphlonuridae |
||
|
Parameletus minor Bengtsson, 1909 |
1, 3 |
Север Европы |
|
Семейство Ameletidae |
||
|
Ameletus inopinatus Eaton, 1885 |
1, 3 |
Сибирь, север Европы |
|
Семейство Baetidae |
||
|
Baetis fuscatus (Linnaeus, 1761) |
1, 3 |
Палеарктика |
|
Baetis inexpectatus (Tshernova, 1928) |
1, 3 |
Европа |
|
Baetis lapponica (Bengtsson, 1912) |
1, 3 |
Север Европы |
|
Baetis muticus (Linnaeus, 1758) |
1, 2, 3 |
Палеарктика |
|
Baetis rhodani (Pictet, 1845). |
1, 3 |
Европа |
|
Baetis scambus Eaton, 1870. |
1, 3 |
Европа |
|
Baetis vernus Curtis, 1830 |
1, 3 |
Палеарктика |
|
Cloeon bifidum Bengtsson, 1912 |
1, 3 |
Палеарктика |
|
Cloeon luteolum (Mueller, 1776) |
1, 2, 3 |
Палеарктика |
|
Cloeon sp . |
6 |
|
|
Семейство Heptageniidae |
||
|
Cinygma lyriformis (McDunnough, 1924) |
1, 3 |
Сибирь, север Европы, Аляска |
|
Ecdyonurus aurantiacus (Burmeister, 1839) |
1, 3 |
Европа |
|
Ecdyonurus joernensis Bengtsson, 1909 |
1, 3 |
Палеарктика |
|
Heptagenia coerulans Rostock, 1878 |
1, 3 |
Европа |
|
Heptagenia sulphurea (Mueller, 1776) |
1, 2, 3 |
Палеарктика |
|
Heptagenia sp . |
6 |
|
|
Семейство Ephemeridae |
||
|
Ephemera lineata Eaton, 1870 |
1, 3 |
Европа |
|
* Ephemera danica Muller, 1764 |
1 |
Европа |
|
Ephemera sp . |
6 |
|
|
Семейство Caenidae |
||
|
Caenis rivulorum Eaton, 1884 |
1, 3 |
Палеарктика |
|
Семейство Leptophlebiidae |
||
|
Habrophlebia lauta McLachlan, 1884 |
1, 3 |
Европа |
|
Leptophlebia submarginata (Stephens, 1835) |
1 |
Европа, Западная Сибирь |
|
Семейство Ephemerellidae |
||
|
Ephemerella aurivillii Bengtsson, 1908 |
1, 3 |
Сибирь, север Европы, Аляска |
|
Ephemerella mucronata (Bengtsson, 1909) |
1, 3 |
Голарктика |
|
ОТРЯД ODONATA |
||
|
Семейство Libellulidae |
||
|
Sympetrum vulgatum (Linnaeus, 1758) |
1 |
Палеарктика |
|
ОТРЯД PLECOPTERA |
||
|
Семейство Perlodidae |
||
|
Arcynopteryx compacta (McLachlan, 1872) |
1, 3 |
Голарктика |
|
Diura bicaudata (Linnaeus, 1758) |
1, 3 |
Голарктика |
|
Diura nanseni (Kempny, 1900) |
1, 2, 3 |
Голарктика |
Продолжение табл. 1
|
Таксон |
Источник |
Географическое распространение |
|
Isoperla grammatica (Poda, 1761) |
1, 3 |
Европа |
|
Isoperla obscura (Zetterstedt, 1840) |
1, 2, 3 |
Палеарктика |
|
Семейство Chloroperlidae |
||
|
Isoptena serricornis (Pictet, 1841) |
1, 3 |
Север Европы |
|
Siphonoperla burmeisteri (Pictet, 1839) |
1, 3 |
Европа |
|
Chloroperlidae indet. |
2, 6 |
|
|
Семейство Nemouridae |
||
|
Amphinemura borealis (Morton, 1894) |
1, 3 |
Палеарктика |
|
Amphinemura standfussi (Ris, 1894) |
1, 3 |
Палеарктика |
|
Nemoura arctica Esben-Petersen, 1910 |
1, 3 |
Голарктика |
|
Nemoura avicularis Morton, 1894 |
1, 2 |
Палеарктика |
|
Nemoura cinerea (Retzius, 1783) |
1, 2, 3 |
Палеарктика |
|
Nemoura flexuosa Aubert, 1949 |
1, 3 |
Европа |
|
Nemurella pictetii Klapalek, 1898 |
1, 3 |
Палеарктика |
|
Protonemura intricata (Ris, 1902) |
1, 3 |
Европа |
|
Семейство Capniidae |
||
|
* Mesocapnia variabilis (Klapalek, 1920) |
1 |
Сибирь, север Европы, Аляска |
|
Capnopsis schilleri (Rostock, 1892) |
1 |
Европа |
|
Семейство Leuctridae |
||
|
Leuctra digitata Kempny, 1899 |
1, 3 |
Европа |
|
Leuctra fusca (Linnaeus, 1758) |
1, 2, 3 |
Палеарктика |
|
Leuctra hippopus Kempny, 1899 |
1, 3 |
Европа |
|
Leuctra sp . |
6 |
|
|
ОТРЯД TRICHOPTERA |
||
|
Семейство Rhyacophilidae |
||
|
Rhyacophila nubila Zetterstedt, 1840 |
1, 2, 3 |
Европа |
|
Семейство Glossosomatidae |
||
|
Agapetus ochripes Curtis, 1834 |
1, 3 |
Европа |
|
Glossosoma intermedia (Klapalek, 1892) |
1, 3 |
Палеарктика |
|
Семейство Polycentropodidae |
||
|
Plectrocnemia conspersa (Curtis, 1834) |
1, 3 |
Палеарктика |
|
Polycentropus flavomaculatus Pictet, 1834 |
1, 2, 3 |
Палеарктика |
|
Семейство Arctopsychidae |
||
|
Arctopsyche ladogensis (Kolenati, 1859) |
1, 2, 3, 6 |
Сибирь, север Европы |
|
Семейство Hydropsychidae |
||
|
Hydropsyche nevae (Kolenati, 1858) |
1, 3 |
Сибирь, север Европы |
|
Семейство Phryganeidae |
||
|
Agrypnia obsoleta (Hagen, 1859) |
1, 3 |
Палеарктика |
|
Semblis phalaenoides (Linnaeus, 1758) |
1, 3 |
Палеарктика |
|
Семейство Sericostomatidae |
||
|
Notidobia ciliaris (Linnaeus, 1761) |
1 |
Европа |
|
Sericostoma personatum (Kirby et Spence, 1826) |
1, 3 |
Европа |
|
Sericostomatidae indet . |
6 |
|
|
Семейство Leptoceridae |
||
|
Athripsodes albifrons (Linnaeus, 1758) |
1, 3 |
Европа |
|
Семейство Brachycentridae |
||
|
Brachycentrus subnubilus Curtis, 1834 |
1, 2 |
Палеарктика |
|
Micrasema gelidum McLachlan, 1876 |
1, 3 |
Сибирь, север Европы |
|
Семейство Lepidostomatidae |
||
|
Lepidostoma hirtum (Fabricius, 1775) |
1, 2 |
Палеарктика |
|
Семейство Apataniidae |
||
|
Apatania stigmatella (Zetterstedt, 1840) |
1, 3 |
Сибирь, север Европы |
|
Apatania crymophila McLachlan, 1880 |
1 |
Сибирь, север Европы |
|
Apatania sp . |
6 |
|
|
Семейство Limnephilidae |
||
|
Anabolia brevipennis Curtis, 1834 |
1, 3 |
Палеарктика |
Продолжение табл. 1
|
Таксон |
Источник |
Географическое распространение |
|
Anabolia furcata Brauer, 1857 |
1, 3 |
Палеарктика |
|
Asynarchus lapponicus Zetterstedt, 1840 |
1, 3 |
Сибирь, север Европы |
|
Halesus digitatus Schrank, 1781 |
1, 3 |
Европа, Западная Сибирь, Алтай |
|
Halesus radiatus (Curtis, 1834) |
1, 3 |
Палеарктика |
|
Halesus tesselatus Rambur, 1842 |
1, 3 |
Палеарктика |
|
Limnephilus rhombicus (Linnaeus, 1758) |
1, 3 |
Голарктика |
|
Potamophylax latipennis (Curtis, 1834) |
1, 6 |
Палеарктика |
|
ОТРЯД MEGALOPTERA |
||
|
Sialis fuliginosa Pictet, 1836 |
1 |
Палеарктика |
|
ОТРЯД DIPTERA |
||
|
Семейство Athericidae |
||
|
Atherix ibis (Fabricius, 1798) |
1, 2 |
Палеарктика |
|
Семейство Chironomidae |
||
|
Ablabesmyia monilis (Linne, 1758) |
2 |
Палеарктика |
|
Boreochlus thienemanni Edwards, 1938 |
4 |
Палеарктика |
|
Brillia flavifrons (Johannsen, 1905) |
4 |
Голарктика |
|
Bryophaenocladius ictericus (Meigen, 1830) |
4 |
Европа |
|
Chaetocladius binotatus (Lindstrom, 1915) |
4 |
Палеарктика |
|
Chaetocladius grandilobus Brundin, 1956 |
4 |
Голарктика |
|
Chaetocladius laminatus Brundin, 1947 |
4 |
Палеарктика |
|
Chaetocladius suecicus (Kieffer, 1916) |
4 |
Европа |
|
Corynoneura arctica Kieffer, 1923 |
4 |
Палеарктика |
|
Corynoneura fittkaui Shlee, 1968 |
4 |
Палеарктика |
|
Corynoneura lobata Edwards, 1924 |
4 |
Голарктика |
|
Corynoneura minuscula Brundin, 1949 |
4 |
Палеарктика |
|
Corynoneura prima Makarchenko et Makarchenko, 2006 |
4 |
Сибирь |
|
Corynoneura sp . |
2 |
|
|
Cricotopus gr . algarum (Kieffer, 1911) |
6 |
Палеарктика |
|
Cricotopus annulator Goetghebuer, 1927 |
4 |
Голарктика |
|
Cricotopus fuscus (Kieffer, 1909) |
6 |
Палеарктика |
|
Cricotopus septentrionalis Hirvenoja, 1973 |
4 |
Палеарктика |
|
Cricotopus gr . silvestris (Fabricius, 1794) |
2 |
Голарктика |
|
Cricotopus similis Goetghebuer, 1921 |
4 |
Европа |
|
Cricotopus tibialis (Meigen, 1804) |
4 |
Голарктика |
|
Cricotopus tremulus (Linnaeus, 1758) |
4 |
Голарктика |
|
Diamesa tonsa (Haliday, 1856) |
4 |
Европа |
|
Eukiefferiella boevrensis Brundin, 1956 |
4 |
Палеарктика |
|
Eukiefferiella brevicalcar (Kieffer, 1911) |
4 |
Палеарктика |
|
Eukiefferiella claripennis (Lundbeck, 1898) |
4 |
Голарктика |
|
Eukiefferiella gr . devonica (Edwards, 1929) |
4 |
Голарктика |
|
Eukiefferiella gracei (Edwards, 1929) |
6 |
Палеарктика |
|
Eukiefferiella minor (Edwards, 1929) |
4 |
Европа |
|
Hydrobaenus fusistilus (Goetghebuer, 1933) |
4 |
Сибирь, север Америки |
|
Krenosmittia novokshonovi Krasheninnikov et Makarchenko, 2009 |
4, 5 |
Бассейн Верхней Вишеры |
|
Limnophyes aagaardi Sether, 1990 |
4 |
Палеарктика |
|
Limnophyes asquamatus Sogaard et Andersen, 1937 |
4 |
Голарктика |
|
Limnophyes bidumus Sether, 1990 |
4 |
Европа |
|
Limnophyes brachytomus (Kieffer, 1922) |
4 |
Голарктика |
|
Limnophyes edwardsi Sether, 1990 |
4 |
Палеарктика |
|
Limnophyes eltoni (Edwards, 1922) |
4 |
Голарктика |
|
Limnophyes margaretae Sether, 1975 |
4 |
Сибирь, север Америки |
|
Limnophyes minimus (Meigen, 1818) |
4 |
Голарктика |
|
Limnophyes natalensis (Kieffer, 1914) |
4 |
Голарктика |
|
Limnophyes schnelli Sether, 1990 |
4 |
Палеарктика |
|
Limnophyes visheraensis Krasheninnikov et Makarchenko, 2009 |
4, 5 |
Бассейн Верхней Вишеры |
|
Metriocnemus fuscipes (Meigen, 1818) |
4 |
Голарктика |
Продолжение табл. 1
|
Таксон |
Источник |
Географическое распространение |
|
Micropsectra gr . junci (Meigen, 1818) |
1 |
Палеарктика |
|
Microtendipes gr . pedellus (De Geer, 1776) |
6 |
Палеарктика |
|
Orthocladius sp . |
4, 6 |
|
|
Paracladius quadrinodosus Hirvenoja, 1973 |
4 |
Голарктика |
|
Paracladopelma camptolabis (Kieffer, 1913) |
2 |
Европа |
|
Paracricotopus sp . |
4 |
|
|
Parakiefferiella bathophila (Kieffer, 1912) |
2, 4 |
Голарктика |
|
Paratanytarsus austriacus (Kieffer, 1924) |
2 |
Европа |
|
Paratrichocladius skirwithensis (Edwards, 1929) |
4 |
Голарктика |
|
Polypedilum exectum Kieffer, 1915 |
2 |
Палеарктика |
|
Polypedilum sordens (Van der Wulp, 1874) |
2 |
Голарктика |
|
Potthastia longimana (Kieffer, 1922) |
2 |
Голарктика |
|
Prodiamesa olivacea Meigen, 1818 |
1 |
Палеарктика |
|
Psectrocladius obvius (Walker, 1856) |
4 |
Голарктика |
|
Pseudodiamesa branickii (Novicki, 1873) |
4 |
Голарктика |
|
Pseudodiamesa gr . nivosa (Goetghebuer, 1928) |
1 |
Палеарктика |
|
Pseudorthocladius curvistilus (Goetghebuer, 1921) |
4 |
Голарктика |
|
Pseudosmittia angusta (Edwards, 1929) |
4 |
Палеарктика |
|
Pseudosmittia mathildae Albu, 1968 |
4 |
Палеарктика |
|
Pseudosmittia obtusa Strenzke, 1960 |
4 |
Европа |
|
Pseudosmittia oxoniana (Edwards, 1922) |
4 |
Голарктика |
|
Rheocricotopus gr . atripes (Kieffer, 1913) |
2 |
Палеарктика |
|
Rheocricotopus effusus (Walker, 1856) |
4 |
Голарктика |
|
Smittia akanduodecima Sasa et Kamimura, 1987 |
4 |
Сибирь |
|
Smittia extrema (Holmgren, 1869) |
4 |
Голарктика |
|
Smittia nudipennis (Goethgebuer, 1913) |
4 |
Европа |
|
Smittia pratorum (Goethgebuer, 1927) |
4 |
Голарктика |
|
Smittia seiryuwexea Sasa, Suzuki et Sakai, 1998 |
4, 5 |
Сибирь |
|
Smittia sp . |
2 |
|
|
Stilocladius intermedius Wang, 1998 |
4 |
Сибирь |
|
Tanytarsus sp . |
2 |
|
|
Thienemanniella gr . clavicornis Kieffer, 1911 |
2 |
Палеарктика |
|
Thienemanniella chuzeduodecimus Sasa, 1984 |
4 |
Сибирь |
|
Thienemanniella majuscula (Edwards, 1924) |
4 |
Голарктика |
|
Thienemannimyia lentiginosa (Fries, 1823) |
2, 6 |
Палеарктика |
|
Tokunagaia rectangularis (Goethgebuer, 1927) |
4 |
Палеарктика |
|
Tokunagaia scutellata (Brundin, 1956) |
4 |
Север Европы |
|
Twetenia sp . |
4 |
|
|
Zalutschia tatrica (Pagast, 1935) |
4 |
Голарктика |
|
Семейство Simuliidae |
||
|
Simuliidae indet . |
1, 2 |
|
|
Limoniidae |
||
|
Antocha vitripennis (Meigen, 1830) |
1 |
Север Европы |
|
Dicranota bimaculata (Schummel, 1829) |
1 |
Европа |
|
Eloeophila maculata (Meigen, 1804) |
1 |
Европа, Малая Азия, Казахстан |
|
Hexatoma sp . |
1 |
Европа |
|
Tipulidae |
||
|
Tipulidae indet . |
1 |
|
|
Tabanidae |
||
|
Tabanidae indet . |
1 |
|
|
Ceratopogonidae |
||
|
Ceratopogonidae indet . |
1 |
|
|
Psychodidae |
||
|
Psychodidae indet . |
1 |
Окончание табл. 1
|
Таксон |
Источник |
Географическое распространение |
|
ОТРЯД COLEOPTERA |
||
|
Семейство Elmidae |
||
|
Elmis sp . |
1 |
|
|
Limnius sp . |
1 |
|
|
Elmidae indet . |
2 |
|
|
Семейство Dityscidae |
||
|
Platambus maculatus Linnaeus, 1758 |
1 |
Палеарктика |
|
Dytiscidae indet . |
1, 6 |
|
|
Семейство Haliplidae |
||
|
Haliplidae indet . |
6 |
Примечания: 1 – оригинальные данные; 2 – Алексевнина, Корионов, 2006; 3 – Крашенинников и др., 2008; 4 – Крашенинников, Макарченко, 2009а; 5 – Крашенинников, Макарченко, 2009б; 6 – Зиновьев, Семёнов, 2013. Звёздочкой (*) отмечены виды, указываемые впервые для Пермского края.
Два вида, подёнка Ephemera danica Muller, 1764 и веснянка Mesocapnia variabilis (Klapalek, 1920), указываются впервые для Пермского края.
Часть таксонов, указанных для бассейна Верхней Вишеры в литературе [Алексевнина, Корио-нов, 2006; Зиновьев, Семёнов, 2013], нами не обнаружены. Таковы поденки Siphlonurus lacustris Eaton, 1870, Heptagenia fuscogrisea (Retzius, 1783), Palingenia sp. и Ephemerella ignita (Poda, 1761), веснянки Perla sp. , Xanthoperla apicalis (Newman, 1836), Taeniopteryx nebulosa (Linnaeus, 1758), Capnia atra Morton, 1896 и Capnia bifrons Newman, 1839, ручейники Chaetopterygopsis maclachlani Stein, 1874, Chaetopteryx sp. , Drusus sp ., Dicosmoecus palatus McLachlan, 1872, Hydropsyche pellucidula (Curtis, 1834) и Goera pilosa (Fabricius, 1775).
Мы считаем, что эти виды были идентифицированы либо ошибочно, либо их определения ненадёжны и требуют подтверждения.
Так, явно ошибочной следует признать идентификацию веснянок Perla sp. , обнаруженных в составе рациона вишерского хариуса Е.А. Зиновьевым и В.В. Семёновым [2013], и ручейников Drusus sp ., указанных для р. Ниолс М.С. Алексев-ниной и А.А. Корионовым [2006]. Дело в том, что, несмотря на многолетнюю работу квалифицированных специалистов, на Урале и сопредельных территориях эти таксоны ни разу не отмечались. Ближайшие находки представителей семейства Perlidae на западе относятся к Прибалтике и Карпатам, а на востоке – к Алтаю и Восточной Сибири [Тесленко, Жильцова, 2009]. По-видимому, как Perla были определены габитуально похожие на них веснянки семейства Perlodidae, массовые в водотоках бассейна Вишеры. Примечательно, что представители семейства Perlodidae в работе Е.А. Зиновьева и В.В. Семёнова [2013], как компонент питания вишерского хариуса, не упоминаются.
Что касается рода Drusus, то эти ручейники на территории России до сих пор не были известны за пределами Кавказа [Ivanov, 2011]. Опыт проведе- ния лабораторных занятий в рамках «Большого практикума по гидробиологии» показывает, что студенты, пользуясь ключами «Определителя…» [1977], применявшегося М.С. Алексевниной и А.А. Корионовым [2006], часто идентифицируют как Drusus разных представителей семейства Lim-nephilidae. Это объясняется выбором в данном источнике крайне неудачного признака для опознавания рода Drusus – резкого поперечного рубца на пронотуме, в менее выраженном виде присутствующем и у других лимнефилид.
Определения ряда видов из вышеприведённого списка недостоверны по той причине, что они могут быть надёжно идентифицированы только по признакам имаго самцов.
Так, поденки S. lacustris принадлежат к группе видов Siphlonurus s.str., в личиночной стадии не отличимых друг от друга. В Пермском крае, наряду с S. lacustris , встречается еще один представитель этой группы – Siphlonurus aestivalis Eaton, 1903 [Паньков, 2007].
Более того, вызывает серьезные сомнения правильность идентификации этих личинок даже до семейства. Нимфы Siphlonuridae обитают в стоячих и слабопроточных водоемах; в руслах водотоков они встречаются крайне редко и только в прибрежье, среди зарослей высшей водной растительности. Отмеченный М.С. Алексевниной и А.А. Корионовым [2006] факт обитания сифлонурид на перекатах горных рек, да еще и в массовом количестве, находится в резком противоречии с их экологическими предпочтениями.
Возможно, как S. lacustris были определены обычные в Вишере и ее притоках личинки Baetis s.str., по окраске и форме тела отдаленно похожие на сифлонурид; в этой связи отсутствие Baetis s.str. в опубликованном М.С. Алексевниной и А.А. Ко-рионовым списке представляется симптоматичным.
Личинки поденок H. fuscogrisea могут быть идентифицированы достаточно надежно, однако некоторые обстоятельства все же заставляют усом- ниться в правильности их определения. Эти насекомые характеризуются как фитофильные формы, избегающие быстрого течения, поэтому присутствие их в составе доминантного комплекса зообен-тоценозов плесов и перекатов горной р. Ниолс [см.: Алексевнина, Корионов, 2006] представляется маловероятным. Возможно, как H. fuscogrisea были идентифицированы широкожаберные личинки E. joernensis, достаточно обычные в реках Уральской горной страны [Паньков, 2007].
Такая ошибка представляется тем более вероятной, что описание сибирского вида E. joernensis в Определителе… [1977], ориентированном на европейскую фауну, отсутствует.
Несмотря на наличие чётких диагностических признаков, большие сомнения вызывает правильность идентификации подёнок рода Palingenia , отмеченных в питании хариуса Верхней Вишеры [Зиновьев, Семёнов, 2013]. Эти насекомые с роющей личинкой обитают в крупных равнинных реках с мягкими грунтами, и их присутствие в горных водотоках представляется маловероятным.
Анализ опубликованных данных заставляет критически отнестись к указанию для бассейна Верхней Вишеры и подёнок E. ignita . Специалист, пользующийся Определителем… [1977], фактически обречён на ошибку – опознать как E. ignita других представителей рода, имеющих основную часть ареала в Сибири – молодых личинок E. mucronata и E. aurivillii , не описанных в данном источнике. В этом убеждает и тот факт, что М.С. Алексевнина и А.А. Корионов [2006] указывают E. ignita как вид, массовый во всех обследованных водотоках, в то время как действительно массовые на Северном Урале E. aurivillii и E. mucronata, в списке отсутствуют.
Среди веснянок из Capniidae и Chloroperlidae по личинкам невозможно различить даже некоторые широко распространенные роды (например, Capnia и Mesocapnia , Xanthoperla и Siphonoperla ), не говоря уже о составляющих их видах [Жильцова, 2000]. Поэтому мы сочли необходимым исключить из списков энтомофауны р. Верхней Вишеры редкий для Прикамья вид X. apicalis , достоверные находки которого до сих пор ограничены одним экземпляром имаго из бассейна Верхней Камы [Паньков, 2004], и оба вида рода Capnia – C. atra и C. bifrons .
Что касается веснянок рода Capnia, то это весьма обычные для Прикамья формы (особенно C. atra). Однако некоторые обстоятельства заставляют усомниться в правильности идентификации этих насекомых даже до рода. Дело в том, что их нимфы развиваются преимущественно в холодное время года, а вылет имаго происходит весной и в начале лета [Тесленко, Жильцова, 2009]. Таким образом, указание веснянок рода Capnia как форм, массово представленных в бентосе в середине лета [Алексевнина, Корионов, 2006], находится в про- тиворечии с их фенологией. По нашим данным, в бассейне Верхней Вишеры водится другой представитель семейства, Mesocapnia variabilis (Kla-palek, 1920), вылетающий в конце лета – начале осени [Тесленко, Жильцова, 2009].
Определённые сомнения вызывает и присутствие веснянок Taeniopteryx nebulosa (Linnaeus, 1758), отмеченных Е.А. Зиновьевым и В.В. Семёновым [2013] в питании хариуса как Taeniopteryx sp. (на Урале водится только T. nebulosa ). Вообще говоря, этот вид по личинке при наличии элементарного опыта идентифицируется достаточно надёжно, но есть обстоятельства, заставляющие отнестись к информации о присутствии его на изучаемой территории с недоверием.
Дело в том, что веснянки T. nebulosa обитают, главным образом, в крупных реках равнин и предгорий. Поэтому сведения о нахождении этих насекомых в горной Вишере вступают в противоречие с широко известными представлениями об их экологии. Возможно, как Taeniopteryx были определены более реофильные представители того же семейства, веснянки Rhabdiopteryx acuminata Kla-palek, 1905, известные из Средней Вишеры [Паньков, 2004], или, что тоже вероятно, какие-то габи-туально похожие на Taeniopterygidae представители семейства Nemouridae.
Из списков ручейников Верхней Вишеры должен быть исключен C. maclachlani , указанный для этой территории М.С. Алексевниной и А.А. Ко-рионовым [2006]. Так, ближайшие достоверные находки генетически сибирского C. maclachlani относятся к Полярному Уралу [Pan’kov, Krasheninnikov, 2016]. Это заставляет отнестись к сведениям об их присутствии в составе трихоптерофау-ны Прикамья с известной осторожностью. К тому же, эти ручейники принадлежат к семейству Lim-nephilidae – таксону, признанному систематиками одной из наиболее проблематичных групп Trichop-tera. В силу того, что преимагинальные стадии ассоциированы менее чем для половины описанных видов лимнефилид, а используемые определительные таблицы часто построены на неадекватных признаках, весьма неполны и содержат много ошибок, среди ручейников Limnephilidae по личинкам невозможно различить даже некоторые обычные роды [Иванов, Григоренко, 1997]. Таким образом, ошибочное указание C. maclachlani для изучаемого региона весьма вероятно.
По этой же причине не могут считаться достоверными определения D. palatus и Chaetopteryx sp., отмеченные в питании хариуса Верхней Вишеры [Зиновьев, Семёнов, 2013], хотя их присутствие в составе трихоптерофауны этой территории вполне возможно. Так, палеарктический D. palatus известен из республики Коми и Башкирии, а виды рода Chaetopteryx встречаются на всём протяжении Урала, исключая его приполярные и полярные районы [Pan’kov, Krasheninnikov, 2016]. Тем не менее, обсуждаемые виды могут быть включены в списки трихоптерофауны Верхней Вишеры только после подтверждения этих находок по взрослым стадиям.
Ручейники H. pellucidula отмечены М.С. Алек-севниной и А.А. Корионовым [2006] в составе зообентоса Верхней Вишеры, и Е.А. Зиновьевым и В.В. Семёновым [2013] в питании вишерского хариуса. Эти насекомые принадлежат к числу массовых элементов трихоптерофауны Пермского края [Паньков, 2000], однако в бассейне Верхней Вишеры они до сих пор не регистрировались. Здесь водится другой представитель рода – Hydropsyche nevae (Kolenati, 1858), имеющий основную часть ареала в Сибири. Описание H. nevae в Определителе… [1977] отсутствует, а принятая в нем система признаков не позволяет отличить его от других темноокрашенных гидропсихид, поэтому ошибочная идентификация этого вида как H. pellucidula вполне вероятна. В этой связи кажется симптоматичным, что действительно массовый в Вишере H. nevae в цитированных работах вообще не фигурирует.
Е.А. Зиновьевым и В.В. Семёновым в питании хариуса Верхней Вишеры отмечены ручейники Goera sp . Учитывая, что в Европе водится лишь G. pilosa , а ближайшие находки других представителей рода относятся к Прибайкалью, Центральной Сибири и Дальнему Востоку [Ivanov, 2011], речь может идти только о G. pilosa . Однако этот вид населяет преимущественно водотоки равнин и предгорий, избегая бурного течения, и его обнаружение в горной реке Северного Урала представляется сомнительным. Примечательно, что G. pilosa , несмотря на значительный объём материалов по фауне ручейников, находящихся в руках специалистов, в бассейне Вишеры никогда не отмечался. Не известен он и из р. Щугор (Северный Урал, республика Коми), в фаунистическом отношении изученной весьма основательно [Шубина, 2006].
Из списка амфибиотических насекомых бассейна Верхней Вишеры исключены несколько видов хирономид. Из них три вида, Eukiefferiella quadridentata Tshernovskij, 1949, Limnophyes sep-tentrionalis Tshernovskij, 1949 и Limnophyes tran-scaucasicus Tshernovskij, 1949, отмеченные М.С. Алексевниной и А.А. Корионовым [2006], в официальных списках зоологической номенклатуры не значатся. Биномены, которыми обозначены эти виды, представляют собой «сомнительные названия» ( nomina nuda ), и что за ними скрывается на самом деле, в настоящее время понять невозможно [Ashe, O’Connor, 2012]. Так, под биноменом E. quadridentata в своё время фигурировали представители родов Eukiefferiella , Twetenia и Tokunagaia [там же]. Под биноменами L. septentrionalis и L. transcaucasicus могут быть означены не только представители рода Limnophyes , но и некоторые другие ортокладиины.
Такие виды, как Corynoneura celeripes Winner, 1852 и Smittia nana Sychova, 1955, в стадии личин- ки неотличимы от других представителей соответствующих родов и могут быть идентифицированы только по признакам имаго самцов. Поэтому в списке хирономид Верхней Вишеры мы оставляем Corynoneura sp. и Smittia sp.
М.С. Алексевнина и А.А. Корионов [2006] указывают для Верхней Вишеры и её притоков хиро-номид Tanytarsus gr . gregarius Kieffer, 1909 и Tanytarsus usmaensis Pagast, 1931. Поскольку личинки известны далеко не для всех видов, а некоторые из них, к тому же, в этой стадии не различаются [Макарченко, 1999], мы исключаем их из списка, оставляя Tanytarsus sp.
Некоторые таксоны, указанные в опубликованных источниках, к настоящему времени фигурируют под другими названиями.
Так, Е.А. Зиновьевым и В.В. Семёновым [2013] в питании вишерского хариуса отмечены подёнки рода Centroptilum , ручейники Stenophylax stellatus Curtis, 1834 и хирономиды Cricotopus biformis Edwards, 1929. Подёнки Centroptilum теперь фигурируют в составе рода Cloeon в ранге подрода [Клюге, 1997]. Биномен S. stellatus признан синонимом Potamophylax latipennis (Curtis, 1834) [Ivanov, 2011], а C. biformis теперь фигурирует под научным названием Cricotopus fuscus (Kieffer, 1909) [Ashe, O’Connor, 2012].
Этими же авторами для рациона хариуса указаны Microtendipes gr . chloris (Meigen, 1818) и Eukiefferiella longicalcar (Kieffer, 1911). В настоящее время первый рассматривается в составе группы видов Microtendipes gr . pedellus (De Geer, 1776), в стадии личинки не различимых [Макар-ченко, 1999], а второй биномен признан младшим синонимом Eukiefferiella gracei (Edwards, 1929) [Ashe, O’Connor, 2012].
Зоогеографическая структура
Фауна донных беспозвоночных бассейна Верхней Вишеры складывается из элементов с различным географическим распространением (см. табл. 1).
Её основу (61.7% совокупного объёма) образует группа широкоареальных видов. Из них наибольшее значение имеют элементы с палеарктическим типом ареалов (37.8%). На долю видов с голарктическим распространением приходится 22.6%. Два вида (1.3%) являются полирегиональными; это малощетинковые черви Lumbriculus variegatus (O.F. Mueller, 1773) и Slavina appendiculata (d'Udekem, 1855).
Некоторые из видов с голарктическим типом ареала приурочены к высоким широтам. Таковы циркумполярные Diura nanseni (Kempny, 1900) и Nemoura arctica Esben-Petersen, 1910.
Существенный вклад в сложение фауны вносят западные палеаркты (25.7%). Большая часть их (23.2%) в своём распространении ограничена Европой; для них Урал является восточной окраиной ареалов. Некоторые из этих видов приурочены к северным областям Европы. Таковы подёнки Pa-rameletus minor Bengtsson, 1909 и Baetis lapponica (Bengtsson, 1912), веснянки Isoptena serricornis (Pictet, 1841), хирономиды Tokunagaia scutellata (Brundin, 1956) и лимонииды Antocha vitripennis (Meigen, 1830). Для одного из этих видов, подёнок B. lapponica, Верхняя Вишера является юговосточным форпостом области сплошного распространения.
Некоторые представители западно-палеарктического комплекса, имея основную часть ареала в Европе, заходят на восток дальше Уральской горной страны, достигая при этом разных рубежей. Так, брюхоногие моллюски Ancylus fluviatilis Mueller, 1774 и подёнки Leptophlebia submarginata (Stephens, 1835) населяют водотоки Западной Сибири, двукрылые Eloeophila maculata (Meigen, 1804) встречаются в Малой Азии и Казахстане, ручейники Halesus digitatus Schrank, 1781 проникают до Алтая.
Определённую роль в зоогеографической структуре фауны играют элементы восточно-палеарктического комплекса, имеющие основную часть ареала в Сибири. На их долю приходится 11.3% её объёма. Для некоторых из них Урал служит западной границей ареалов. Таковы хирономиды Cory-noneura prima Makarchenko et Makarchenko, 2006, Hydrobaenus fusistilus (Goetghebuer, 1933), Limno-phyes margaretae Sether, 1975, Smittia akanduode-cima Sasa et Kamimura, 1987, Smittia seiryuwexea Sasa, Suzuki et Sakai, 1998, Stilocladius intermedius Wang, 1998 и Thienemanniella chuzeduodecimus Sasa, 1984.
Некоторые восточные палеаркты, перевалив Урал, проникают более или менее далеко в Европу по её северным территориям. Часть из них, такие, как веснянка M. variabilis , в своём движении на запад останавливаются на рубеже Тимана. Другие – подёнка Cinygma lyriformis (McDunnough, 1924) и ручейник Apatania crymophila McLachlan, 1880 – достигают Кольского полуострова. Третьи – двустворчатый моллюск Parasphaerium rectidens (Sta-robogatov et Streletzkaja, 1967), подёнка E. aurivillii , ручейник A.ladogensis, H. nevae , Mi-crasema gelidum McLachlan, 1876 и Apatania stig-matella (Zetterstedt, 1840) – идут на запад вплоть до Скандинавии.
Из числа восточных палеарктов наиболее обширную европейскую часть ареала имеют подёнки Ameletus inopinatus Eaton, 1885 и Asynarchus lap-ponicus Zetterstedt, 1840. Эти насекомые, помимо северной полосы, встречаются также в горах Центральной Европы, демонстрируя в пределах Западной Палеарктики ареал арктоальпийского типа.
Для ряда представителей циркумполярного голарктического и восточно-палеарктического зоогеографических комплексов, подёнки A. inopinatus , веснянок N. arctica и M. variabilis и ручейника A.
lapponicus , бассейн Верхней Вишеры служит южным форпостом их сплошного распространения на севере Европы
Из числа элементов фауны донных беспозвоночных Верхней Вишеры исключительно узкими ареалами характеризуются два вида хирономид (1.3%), Krenosmittia novokshonovi Krasheninnikov et Makarchenko, 2009 и Limnophyes visheraensis Krasheninnikov et Makarchenko, 2009, до сих пор не известные за пределами этой территории.
Таким образом, для фауны донных беспозвоночных бассейна Верхней Вишеры характерна значительная доля широкоареальных форм, сравнительно небольшое участие элементов западнопалеарктического зоогеографического комплекса и весьма заметная роль восточных палеарктов. Эти особенности в той или иной мере характерны для гидрофауны Урала в целом и отражают историю её становления.
Судя по гетерогенному и малоспецифическому зоогеографическому составу и преобладанию широкоареальных форм, гидрофауна Урала очень молода и имеет типичный аллохтонный характер [Паньков, 2000]. Причём в бассейне Верхней Вишеры эти черты выражены в существенно большей степени, чем на остальной территории Прикамья. Учитывая особенности географического положения и климата бассейна Верхней Вишеры, это представляется вполне закономерным. Так, удалённость от европейских генетических центров и суровость климата обусловливает относительную бедность гидрофауны Верхней Вишеры сравнительно теплолюбивыми элементами западно-палеарктического зоогеографического комплекса, а близость к североевропейскому фаунистическому коридору, служившему миграционным путём для генетически сибирских элементов, объясняет существенную роль последних в её составе.
В то же время среди элементов гидрофауны Верхней Вишеры присутствуют два эндемичных вида хирономид, что может интерпретироваться как необычное для гидрофауны Урала явление, придающее ей черты автохтонности. Это представляется тем более вероятным, что именно для хирономид, в отличие от других групп амфибиоти-ческих насекомых, отмечается наибольшее таксономическое разнообразие в высоких широтах [Крашенинников, 2011], что наводит на мысли об их склонности к интенсивному видообразованию в послеледниковое время. С другой стороны, учитывая слабую изученность фаун хирономид на обширных территориях Палеарктики, нельзя исключить и вероятность обнаружения этих видов далеко за пределами Урала.
По своему таксономическому составу фауна донных беспозвоночных бассейна Верхней Вишеры занимает положение, промежуточное между фаунами бассейна Печоры и остальной части бассейна р. Камы (несколько ближе к бассейну р. Пе- чоры). Так, из 68 видов подёнок, веснянок и ручейников, выявленных в бассейне Верхней Вишеры, 63 вида являются общими с видами, обнаруженными в бассейну Печоры, и 58 – с видами бассейна Камы (без Верхней Вишеры).
Гидрофауны бассейна Печоры и Верхней Вишеры сближает наличие в составе последней подёнок A. inopinatus , B. lapponica и E. danica , веснянок N. arctica и M. variabilis , ручейников Seri-costoma personatum (Kirby et Spence, 1826), A. stig-matella , A. brevipennis , Anabolia furcata Brauer, 1857, A. lapponicus , неизвестных из других частей бассейна Камы.
С остальной частью гидрофауны бассейна р. Камы общими являются подёнки Ecdyonurus au-rantiacus (Burmeister, 1839) и Heptagenia coerulans Rostock, 1878, а также ручейники Agapetus ochripes Curtis, 1834, Athripsodes albifrons (Linnaeus, 1758) и Halesus digitatus Schrank, 1781.
Учитывая, что из десяти видов первой группы семь (кроме E. danica , S. personatum и A. furcata ) принадлежат к элементам циркумполярного голарктического и восточно-палеарктического зоогеографических комплексов, а все пять представителей второй группы – это западные палеаркты, отмеченные сходства и различия представляются закономерными.
июле) водой и валунно-галечными грунтами. Здесь формируется три основных типа биотопов, из которых наибольшее значение имеют плёсы. Второе место по своей значимости занимают перекаты. И, наконец, в качестве самостоятельного биотопа фигурируют ямы в подвалье перекатов. Скорость течения повсюду значительна – от 0.4 до 1.0 м/с на плёсах и до 3.0 м/с на перекатах, и даже в ямах она достигает 0.4–0.6 м/с.
На плёсах в 2016 г. формировался зообентоце-ноз, в составе которого преобладали личинки комаров-звонцов и ручейники A. ladogensis (табл. 2). Им сопутствовали подёнки B. vernus , личинки мух A. ibis , личинки и взрослые жуки Elmis sp. , веснянки A. compacta . Постоянным компонентом донной фауны являлись подёнки H. sulphurea , ручейники A. crymophila и Glossosoma intermedia (Klapalek, 1892). Численность сообщества достигала довольно значительной величины (6 860 экз./м2) при умеренной биомассе (3 460 мг/м2).
На перекатах Верхней Вишеры формировался монодоминантный зообентоценоз с резким преобладанием ручейников A. ladogensis . Подёнки B. vernus , личинки мух A. ibis , личинки и взрослые жуки Elmis sp . , веснянки A. compacta продолжали присутствовать в составе сообщества на правах субдоминантов. Особенность донной фауны пере-
Сообщества зообентоса
В пределах изученного участка Верхняя Вишера представляет собой быструю горную реку шириной 30–70 м с чистой прохладной (до 14–18ºС в катов составляло участие в его составе личинок мошек Simuliidae. Численность зообентоценозов перекатов достигала 5 350 экз./м2 при биомассе 3 470 мг/м2 (табл. 2).
Таблица 2
Структура населения, численность (N, экз./м2) и биомасса (B, мг/м2) зообентоценозов Верхней
Вишеры
|
Вид, группа |
Плёсы |
Перекаты |
Ямы |
Плёс в районе устья Большой Мойвы |
||||
|
Июль 2016 г. |
Август 2007 г. |
|||||||
|
N |
B |
N |
B |
N |
B |
N |
B |
|
|
A. ladogensis |
290 |
990 |
370 |
2 090 |
310 |
930 |
170 |
1 740 |
|
A. compacta |
40 |
120 |
20 |
70 |
50 |
240 |
10 |
60 |
|
A. crymophila |
10 |
10 |
10 |
20 |
10 |
20 |
310 |
600 |
|
A. ibis |
100 |
270 |
30 |
60 |
10 |
10 |
0 |
0 |
|
A. ochripes |
0 |
0 |
0 |
0 |
0 |
0 |
80 |
100 |
|
B. vernus |
180 |
510 |
290 |
300 |
310 |
180 |
160 |
70 |
|
Chironomidae |
4 880 |
1 210 |
4 220 |
590 |
1 450 |
110 |
10 |
30 |
|
Elmis sp . |
1 300 |
260 |
260 |
150 |
160 |
30 |
10 |
10 |
|
G. intermedia |
10 |
10 |
30 |
50 |
40 |
50 |
30 |
40 |
|
H. sulphurea |
10 |
50 |
10 |
30 |
0 |
0 |
30 |
180 |
|
Simuliidae |
0 |
0 |
90 |
60 |
0 |
0 |
0 |
0 |
|
Прочие |
40 |
30 |
20 |
30 |
40 |
30 |
340 |
1 870 |
|
Всего |
6 860 |
3 460 |
5 350 |
3 470 |
2 380 |
1 600 |
1 150 |
4 700 |
В подвальях перекатов (ямах) складывалось качественно и количественно обедненное сообщество, в структуре которого ручейники A. ladogensis сохраняли своё безусловное лидерство. На фоне снижения показателей развития других беспозвоночных возрастала роль веснянок A. compacta. Численность донной фауны в данных биотопах составляла 2 380 экз./м2 при биомассе 1 600 мг/м2 (табл. 2).
Эти результаты близки к нашим данным, полученным ранее. Так, в середине августа 2007 г. численность зообентоса на плёсе ниже устья р. Мойвы составила 1 150 экз./м2 при биомассе 4 700 мг/м2. В составе сообщества доминировали ручейники A. ladogensis . Им сопутствовали ручейники A. crymophila и A. ochripes , а также подёнки H. sulphurea и B. vernus .
Сходные параметры развития донной фауны приводятся и в литературных источниках.
Так, по данным М.С. Алексевниной и А.А. Ко-рионова [2006], на одном из перекатов р. Верхней Вишеры в середине лета 1984 г. численность составляла 481 экз./м2 при биомассе 2.1 г/м2. Основу сообщества (свыше половины биомассы) образовывали ручейники. К числу субдоминантов относились личинки двукрылых. Наряду с ними, были зарегистрированы подёнки, веснянки, малощетинковые черви и двустворчатые моллюски.
В июне 1994 г. на перекате Верхней Вишеры близ устья Хальсории численность зообентоса достигала 639 экз./м2 при биомассе 6.4 г/м2. В составе сообщества доминировали ручейники и подёнки, которым сопутствовали моллюски, личинки двукрылых, веснянок и жуков, а также малощетинковые черви [Зиновьев, Семёнов, 2013].
Большая численность, но меньшая биомасса донной фауны в июле 2016 г., по сравнению с июнем 1994 и августом 2007 гг., объясняется характером сезонной динамики речных зообентоцено-
Таблица 3
Структура населения, численность (N, экз./м2) и биомасса (B, мг/м2) зообентоценозов некоторых притоков Верхней Вишеры
|
Вид, группа |
Хальсория |
Ниолс |
Лопья |
Малая Мойва |
Малый Лиственничный |
Большая Мойва |
||||||
|
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
|
|
A. ladogensis |
770 |
1 900 |
0 |
0 |
50 |
890 |
0 |
0 |
0 |
0 |
50 |
1 510 |
|
A. crymophila |
10 |
10 |
10 |
20 |
80 |
230 |
40 |
100 |
80 |
420 |
180 |
320 |
|
R. nubila |
10 |
30 |
500 |
850 |
40 |
770 |
280 |
3 240 |
60 |
1 640 |
10 |
170 |
|
E. joernensis |
10 |
30 |
20 |
30 |
10 |
40 |
10 |
30 |
100 |
320 |
10 |
30 |
|
P. conspersa |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
20 |
440 |
0 |
0 |
|
A. compacta |
10 |
30 |
80 |
400 |
60 |
600 |
60 |
320 |
160 |
980 |
20 |
130 |
|
H. sulphurea |
10 |
40 |
10 |
30 |
80 |
720 |
20 |
150 |
100 |
700 |
20 |
110 |
|
A. fluviatilis |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
10 |
220 |
|
A. ochripes |
10 |
10 |
10 |
20 |
90 |
260 |
1 120 |
1 440 |
40 |
100 |
140 |
360 |
|
Elmis sp . |
10 |
10 |
450 |
280 |
20 |
20 |
10 |
10 |
60 |
40 |
20 |
10 |
|
Baetis spp . |
20 |
10 |
650 |
1 650 |
10 |
20 |
80 |
100 |
10 |
10 |
60 |
40 |
|
Chironomidae |
2 350 |
110 |
6 230 |
470 |
30 |
10 |
640 |
170 |
130 |
30 |
730 |
250 |
|
Simuliidae |
4 960 |
2 920 |
20 |
10 |
10 |
10 |
30 |
20 |
20 |
10 |
30 |
20 |
|
Прочие |
10 |
10 |
30 |
620 |
10 |
10 |
30 |
20 |
20 |
10 |
20 |
10 |
|
Всего |
8 180 |
5 110 |
8 010 |
3 980 |
1 300 |
3 580 |
2 320 |
5 600 |
800 |
4 700 |
1 300 |
3 180 |
В то же время, эти реки обладают и некоторыми индивидуальными особенностями.
Так, донная фауна р. Хальсории отличается массовым развитием личинок мошек Simuliidae, на долю которых приходится больше половины зов: в середине лета происходит интенсивное размножение амфибиотических насекомых, не скомпенсированное вылетом имаго, поэтому численность донной фауны в данный период велика. В то же время, донные беспозвоночные в середине лета представлены, в основном, самыми молодыми формами, из-за чего биомасса зообентоса достигает минимальных значений в её годичном цикле [Паньков, 1997].
Изученные притоки Верхней Вишеры по структуре донных сообществ подразделяются на четыре группы.
К первой группе относятся р. Хальсория, Лопья в её среднем течении и Большая Мойва. Это небольшие горные речки длиной от 17 (Хальсория) до 51 км (Большая Мойва) и шириной в районе устья 15–25 м, сходные по своему гидрологическому режиму с главной рекой.
В этих водотоках формируются зообентоцено-зы, по составу и уровню развития близкие к сообществам Верхней Вишеры. Их численность изменяется в пределах 1 300–8 180 экз./м2, биомасса – от 3 180 до 5 110 мг/м2. В состав доминантного комплекса зообентоценозов, как и в Верхней Вишере, входят ручейники A. ladogensis , A. crymophila и A. ochripes , веснянки A. compacta и подёнки H. sulphurea . Спецификой этих сообществ, придающей им характерные черты, является существенная роль ручейников R. nubila , довольно редких в русле Верхней Вишеры (табл. 3).
массы донной фауны.
Река Лопья в приустьевом её участке характеризуется значительно меньшим уровнем развития зообентоса, биомасса которого составляет здесь всего 460 мг/м2 при численности 1 580 экз./м2. Ве- роятно, это связано с особенностями её грунтов, представленных здесь мелкой галькой и гравием, покрытыми перемываемыми песчаными дюнами. В составе зообентоса доминируют ручейники P. latipennis и личинки хирономид.
Ко второй группе притоков Верхней Вишеры относятся р. Ниолс, Малая Мойва и ручей Малый Лиственничный. Протяжённость этих водотоков изменяется от 11 км (Малый Лиственничный) до 26 км (Ниолс). Как и для главной реки, для них характерны галечно-валунные и галечные грунты и высокая скорость течения. Их особенностью, по сравнению с Верхней Вишерой и её более крупными притоками, является сравнительно низкая температура воды (12–14ºС).
Численность зообентоса в этих реках изменяется в пределах 800–8 010 экз./м2, биомасса – от 3 980 до 5 600 мг/м2, что близко к аналогичным показателям, известным для Верхней Вишеры, Хальсории, Лопьи и Большой Мойвы.
Опубликованные данные по зообентосу водотоков рассматриваемой группы ограничены р. Ниолс [Алексевнина, Корионов, 2006]. Авторы цитированного источника указывают численность донной фауны – 2 642 экз./м2 на перекате, и 484 экз./м2 на плёсе, при биомассе 6.06 и 6.86 г/м2 соответственно.
Характерными чертами р. Ниолса, Малой Мойвы и ручья Малый Лиственничный является сравнительно большое значение ручейников R. nubila и полное отсутствие A. ladogensis в составе донных сообществ. В то же время, здесь сохраняется роль ручейников A. crymophila и A. ochripes , веснянки A. compacta и подёнки H. sulphurea , что сближает эти водотоки с таковыми Верхней Вишеры.
К третьей группе притоков Верхней Вишеры относится р. Муравей. Длина этой речки составляет 19 км при ширине в её нижнем течении около 10 м. Особенностями р. Муравей в пределах изученного участка являются грунты, представленные крупными валунами и галькой, покрытыми мощными обрастаниями из водных мхов, низкая температура воды, не превышающая 10ºС, умеренная скорость течения, достигающая 0.8 м/с.
В приустьевом участке этого водотока формируется своеобразный зообентоценоз, характеризующийся весьма высокими показателями численности и биомассы, которые достигают здесь 34 010 экз./м2 и 30 070 мг/м2 соответственно. Ядро сообщества образуют личинки комаров-звонцов, на долю которых приходится 93% численности и 76% общей массы донной фауны. Наряду с ними сравнительно велико значение ручейника P. latipennis и подёнки A. inopinatus . К числу второстепенных видов относятся подёнки Baetis rhodani (Pictet, 1845), Cinygma lyriformis (McDunnough, 1924), и Ephemerella mucronata (Bengtsson, 1909), веснянки Isoperla grammatica (Poda, 1761), Amphinemura borealis (Morton, 1894) и Nemoura sp., ручейник R. nubila .
Структурные характеристики зообентоса р. Муравей и высокие показатели его развития, по-видимому, обусловлены мощными обрастаниями из водных мхов, в которых личинки хирономид находят убежище и обильную пищу.
Приустьевой участок р. Муравей вряд ли может считаться показательным для всего водотока. Об этом позволяют судить и опубликованные материалы, в которых приводятся гораздо более скромные показатели численности и биомассы донной фауны переката этой реки, достигающих 965 экз./м2 и 3.33 г/м2 соответственно [Алексевни-на, Корионов, 2006], что вполне сопоставимо с показателями других рек бассейна Верхней Вишеры. Но и литературные данные говорят о своеобразии структуры зообентоценозов р. Муравей, в составе которых велико значение личинок комаров-звонцов, подёнок родов Baetis , Ephemerella и семейства Heptageniidae, веснянок родов Isoperla , Nemoura и семейства Capniidae, ручейников Lim-nephilidae.
Четвёртую группу притоков Верхней Вишеры составляют небольшие безымянные ручьи. Особенностями этих объектов являются грунты, представленные слабо окатанными глыбами и валунами, а также очень низкая температура воды, не превышающая 6–8ºС.
В приустьевых участках ручьёв формируются зообентоценозы, основу которых слагают подёнки рода Baetis , веснянки I. grammatica и A. borealis , ручейники A. crymophila и P. latipennis . Им сопутствуют личинки комаров-звонцов Chironomidae и мошек Simuliidae. Численность донной фауны составляет 1 720 экз./м2 при биомассе 2 490 мг/м2.
Выводы
-
1. В составе зообентоса бассейна Верхней Вишеры установлено 176 видов и форм. Наиболее разнообразно представлены личинки комаров-звонцов (80 видов и форм). Богаты видами ручейники (25), подёнки (23) и веснянки (20). Среди прочих беспозвоночных зарегистрированы нехи-рономидные двукрылые (10), малощетинковые черви (6), жесткокрылые (5), двустворчатые (2) и брюхоногие (1) моллюски, а также турбеллярии (1), водяные клещи (1), стрекозы (1) и вислокрыл-ки (1). Два вида, подёнка E. danica и веснянка M. variabilis , указываются впервые для Пермского края.
-
2. Фауна донных беспозвоночных бассейна Верхней Вишеры складывается из элементов с различным географическим распространением. Её основу (61.7%) образует группа широкоареальных видов. На долю элементов с палеарктическим типом ареалов приходится 37.8% объёма фауны, на голарктов – 22.6%. Два вида (1.3%) являются по-лирегиональными. Существенный вклад в сложение фауны вносят западные (25.7%) и восточные
-
3. В р. Верхней Вишере и её притоках – Халь-сории, Лопье и Большой Мойве – формируются зообентоценозы, в составе которых доминируют ручейники A. ladogensis и A. crymophila , веснянка A. compacta и подёнка H. sulphurea . Их численность изменяется в пределах 1 150–8 180 экз./м2, биомасса – от 1 600 до 5 110 мг/м2.
-
4. В р. Ниолс, Малая Мойва и ручье Малый Лиственничный формируются зообентоценозы, характеризующиеся доминированием ручейников R. nubila и A. crymophila , веснянки A. compacta и подёнки H. sulphurea . Численность донной фауны изменяется в пределах 800–8 010 экз./м2, биомасса – от 3 980 до 5 600 мг/м2.
-
5. В приустьевом участке р. Муравей формируется зообентоценоз, характеризующийся высокими показателями численности и биомассы (34 010 экз./м2 и 30 070 мг/м2 соответственно). Ядро сообщества образуют личинки комаров-звонцов. Сравнительно велико значение ручейника P. lati-pennis , подёнок A. inopinatus , B. rhodani , C. lyri-formis и E. mucronata , веснянок I. grammatica , A. borealis и Nemoura sp ., ручейника R. nubila .
-
6. В приустьевых участках небольших безымянных ручьёв формируются зообентоценозы, основу которых слагают подёнки рода Baetis , веснянки I. grammatica и A. borealis , ручейники A. crymophila и P. latipennis . Им сопутствуют личинки комаров-звонцов Chironomidae и мошек Simu-liidae. Численность донной фауны составляет 1 720 экз./м2 при биомассе 2 490 мг/м2.
(11.3%) палеаркты. Исключительно узкими ареалами характеризуются два вида хирономид, K. no-vokshonovi и L. visheraensis , не известные за пределами этой территории.
Список литературы Фауна и сообщества донных беспозвоночных водотоков бассейна Верхней Вишеры
- Алексевнина М.С., Корионов А.А. Донная фауна Верхней Вишеры и ее притоков в районе Вишерского заповедника//Заповедник Вишерский: итоги и перспективы исследований (15 лет с основания). Пермь, 2006. С. 76-79
- Жильцова Л.А. Отряд Plecoptera -Веснянки//Определитель насекомых Европейской части СССР. Т. 1. М., Л.: Наука, 1964. С. 177-200
- Жильцова Л.А. Обзор веснянок сем. Leuctridae (Plecoptera) фауны России и сопредельных стран//Фауна, проблемы экологии, этологии и физиологии амфибиотических и водных насекомых России: материалы VI Всерос. трихоптеролог. симпозиума. Воронеж, 2000. С. 11-15
- Жильцова Л.А. Веснянки (Plecoptera). Группа Eu-holognatha. СПб.: Наука, 2003. 538 с
- Зиновьев Е.А., Семёнов В.В. Рыбы Вишерского заповедника. Пермь, 2013. 96 с
- Иванов В.Д., Григоренко В.Н. Исследования личинок ручейников фауны России//Проблемы происхождения, систематики и экологии ручейников России и сопредельных территорий: материалы V Всерос. трихоптеролог. симпозиума. Воронеж, 1997. С. 7-12
- Иванов В.Д., Григоренко В.Н., Арефина Т.И. Trichoptera ручейники//Определитель пресноводных беспозвоночных России и сопредельных территорий. СПб., 2001. Т. 5. Высшие насекомые. С. 7-72
- Качалова О.Л. Отряд Trichoptera -Ручейники//Определитель насекомых Европейской части СССР. Л.: Наука, 1987. Т. IV. С. 107-193
- Клюге Н.Ю. Поденки (Ephemeroptera)//Определитель пресноводных беспозвоночных России и сопредельных территорий. СПб., 1997. Т. 3. Паукообразные. Низшие насекомые. С. 175-220
- Комлев А., Черных Е. Реки Пермской области. Пермь, 1984. 214 c
- Крашенинников А.Б. Фауна и систематика хирономид (Diptera, Chironomidae) Урала и Приуралья: дис.... канд. биол. наук. Пермь, 2011. 227 с
- Крашенинников А.Б., Макарченко Е.А. К фауне хирономид подсемейств Podonominae, Diamesi-nae и Orthocladiinae (Diptera, Chironomidae) заповедника Вишерский и прилегающих территорий (Северный Урал)//Евразиатский энтомол. журнал, 2009а. Т. 8(3). С. 335-340
- Крашенинников А.Б., Макарченко Е.А. Новые и малоизвестный виды хирономид (Diptera, Chi-ronomidae, Orthocladiinae) из Вишерского государственного природного заповедника (Северный Урал)//Евразиатский энтомол. журнал, 2009б. Т. 8. С. 97-101
- Крашенинников А.Б. и др. Амфибиотические насекомые северо-востока Пермского края и сопредельных территорий (поденки, веснянки, ручейники)//Вестник Пермского университета. 2008. Вып. 9(25). Биология. С. 15-26
- Макарченко Е.А. Chironomidae комары-звонцы//Определитель пресноводных беспозвоночных России и сопредельных территорий. СПб., 1999. Т. 4. Высшие насекомые. Двукрылые. С. 210-295
- Определитель пресноводных беспозвоночных Европейской части СССР (планктон и бентос)/под ред. Л.А. Кутиковой и Я.И. Старобогатова. Л.: Гидрометеоиздат, 1977. 510 с
- Паньков Н.Н. Структурные и функциональные характеристики сообществ зообентоса реки Сыл-вы (бассейн Камы): дис.. канд. биол. наук. Пермь, 1997. 230 с.
- Паньков Н.Н. Зообентос текучих вод Прикамья. Пермь: Гармония, 2000. 192 с
- Паньков Н.Н. Фауна веснянок (Plecoptera) Прикамья//Фауна, вопросы экологии, морфологии и эволюции амфибиотических и водных насекомых России: материалы II Всерос. симпозиума по амфибиотич. и водн. насекомым. Воронеж, 2004. С. 121-126
- Паньков Н.Н. Поденки (Ephemeroptera) Пермского Прикамья//Проблемы водной энтомологии России и сопредельных стран: материалы III Всерос. симпозиума по амфибиотич. и водн. насекомым. Воронеж, 2007. С. 222-228.
- Паньков Н.Н. и др. Фауна родников Урала и Предуралья (Пермское Прикамье)//Рыбные ресурсы Камско-Уральского региона и их рациональное использование: материалы науч.-практ. конф. Пермь, 2008. С. 146-151
- Паньков Н.Н., Крашенинников А.Б. Зообентос родников Урала и Предуралья (Пермское Прикамье)//Вестник Пермского университета. Сер. Биология. 2012. Вып. 1. С. 18-24
- Ресурсы поверхностных вод СССР. Основные гидрологические характеристики. Л.: Гидрометео-издат, 1967. Т. 11, вып. 1. Кама. 536 с
- Тесленко В.А., Жильцова Л.А. Определитель веснянок Insecta, Plecoptera России и сопредельных стран. Имаго и личинки. Владивосток: Дальнаука, 2009. 382 с
- Химическая география и гидрохимия Пермской области. Пермь, 1967. 182 с
- Чернова О.А. Отряд Ephemeroptera -Поденки//Определитель насекомых Европейской части СССР. М.; Л.: Наука, 1964. Т. 1. С. 110-136
- Шкляев А.С., Балков В.А. Климат Пермской области. Пермь, 1963. 191 с
- Шубина В.Н. Бентос лососевых рек Урала и Тимана. СПб.: Наука, 2006. 401 с
- Ashe P, O 'Connor J.P. A World Catalogue of Chironomidae (Diptera). Dublin: Irish Biogeographical Society and National Museum of Ireland, 2009. P. 1. 445 p
- Ashe P, O 'Connor J.P. A World Catalogue of Chironomidae (Diptera). Dublin: Irish Biogeographical Society and National Museum of Ireland, 2012. P. 2. 968 p
- Ivanov V.D. Caddisflies of Russia: Fauna and biodiversity//Zoosymposia. 2011. P. 171-209
- Pan'kov N.N., Krasheninnikov A.B. Current state of knowledge of a faunal inventory of Trichoptera (Hexapoda, Trichoptera) from the Ural Mountains and neighboring regions//Zoosymposia. 2016. P. 331-339
