Фауна, экология и зоогеография млекопитающих Валдайского поозерья: ретроспектива и современное состояние

Автор: Хляп Людмила Айзиковна, Шварц Евгений Аркадьевич, Баскевич Марина Исаковна, Леонтьева Ольга Александровна, Черепанова Елена Владимировна, Глазов Петр Михайлович, Николаев Валерий Иванович, Тишков Аркадий Александрович
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Зоология
Статья в выпуске: 1, 2017 года.
Бесплатный доступ
Проведена ревизия современной фауны млекопитающих Валдайского поозерья, дан ее полный список и проанализированы изменения териофауны и зоогеографии края от 1930-х гг. до наших дней. Представлены результаты экологических исследований последних десятилетий, в том числе учетов крупных млекопитающих. С помощью современных генетических методов получены новые сведения о видах-двойниках и внутривидовых формах представителей отрядов Rodentia и Eulipotyphla, которые подтверждают большую роль ледниковья в формировании фауны и в видообразовании. Современный этап формирования териофауны характеризуется появлением новой зоогеографической группы чужеродных видов. Чужеродные виды не только увеличили разнообразие фауны млекопитающих края, но и изменили облик и функционирование околоводных экосистем.
Млекопитающие, фауна, виды-двойники, кариоформы, зоогеография, валдай, динамика численности, биологические инвазии
Короткий адрес: https://sciup.org/146116688
IDR: 146116688 | УДК: 599.3/591.8
The fauna, ecology and zoogeography of mammals in Valdai lakeland: retrospective and current status
A revision of modern mammal fauna of the Valdai Lakeland is carried out. Its full list is provided. Transformations of theriofauna and zoogeography from 1930s to the present day are analyzed. The results of the ecological research of the last decades, including winter monitoring data on large mammals, are presented. New information about sibling species and intraspecific forms of Rodentia and Eulipotyphla was obtained by the modern genetic techniques. These data confirm the important role of the last glaciation in the fauna and species formation. The current stage of theriofauna formation is characterized by the appearance of the new zoogeographical group of alien species. Alien species did not only increase the diversity of mammal fauna of the Valdai Lakeland, but also changed functioning of riparian ecosystems.
Текст научной статьи Фауна, экология и зоогеография млекопитающих Валдайского поозерья: ретроспектива и современное состояние
Введение. В 2016 г. исполнилось 80 лет после публикации в Зоологическом журнале работ С.У. Строганова (1936а, б), сохранивших свою актуальность. Этот своеобразный юбилей одного из классических исследований фауны. С.У.Строганов (1904-1960) многие годы изучал млекопитающих Валдайской возвышенности, будучи учеником С.И. Огнева (Жизнь..., 1962). В двух статьях 1936 г. впервые дан полный список фауны млекопитающих Валдая. Приведены краткие очерки о 55 видах млекопитающих, в которых содержатся сведения по систематике (часто это результаты собственных исследований автора) и некоторым морфологическим признакам, охарактеризовано распределение видов по территории края, численность и биотопическая приуроченность. Завершаются статьи зоогеографическим анализом фауны. Столь глубокое фаунистическое исследование С.У. Строганова стало вехой для изучения млекопитающих региона. Без ссылок на эти публикации не обходится ни одна из последующих териологических работ, хоть в малой степени касающихся Валдая.
За 80-летний период представления о видовом составе и структуре некоторых групп млекопитающих претерпели изменения: были обнаружены виды-двойники и (или) генетически дискретные внутривидовые формы. Изменилось состояние популяций животных, возросла степень их изученности.
К числу регионов Валдая, хорошо изученных в териологическом отношении и привлекающих зоологов биологическим разнообразием, относится Валдайской поозерье (Кузнецов, Тишков, 2012). Во время последнего оледенения Русской равнины этот регион лежал в краевой ледниковой зоне, что отразилось как на рельефе (конечно-моренные гряды и холмы, камы, озовые гряды, зандровые поля и др.), так и на своеобразии формировании биоты (возможность сохранения рефугиумов, смешение фаун и др.). В современную эпоху он располагается на границе южной тайги и хвойно-широколиственных лесов. Показана близость этого региона к белорусским и польским лесам, включающим неморальный комплекс. Немаловажна древняя многовековая антропогенная трансформация этого региона, которая привела к обезлесиванию больших пространств, распашке земель, фрагментации местообитаний, формированию уникального лесо-лугово-полевого ландшафта (Белоновская и др., 2014; Matuszkevicz et al., 2014). Однако в последние годы в связи со снижением экономического развития, депрессивным сельским хозяйством и усиливающейся депопуляцией местного населения наметилась противоположная тенденция: увеличение лесопокрытой площади, зарастание пашен и лугов при росте уровня рекреационного воздействия на побережьях крупных озер и в окрестностях г. Валдай. Имеющиеся данные позволяют проанализировать изменения фауны и населения млекопитающих, произошедшие под воздействием этих разнонаправленных природных и антропогенных процессов на фоне изменения климата.
Относительно недавно проанализирована тысячелетняя история изменения фауны млекопитающих Валдайского поозерья (Тишков, 2013).
Цель настоящей сообщения — оценить какие изменения произошли в систематике, фауне и населении млекопитающих Валдайского поозерья за прошедшие 80 лет. При этом ставилась задача сравнить новые материалы с изложенным ранее С.У. Строгановым, оставляя углубленный анализ причинно-следственных зависимостей в изменении фауны региона для будущих публикаций.
Материал и методика. К Валдайскому поозерью мы относим широкую озерную полосу, протянувшуюся от озера Селигер (на юге) до озера Боровно (на севере), включая озера Велье, Валдайское, Ужин и др. Ландшафтное, биологическое и историческое своеобразие региона определило создание в этом регионе Национального парка «Валдайский» (НПВ) площадью 158 тыс. га (рис. 1). В наши дни большая его часть (83,9%) занята лесами, преимущественно еловошироколиственными и сосновыми. Луга и кустарники занимают 15,2% территории, озера и водотоки - 5,6%. Характерна мозаичность, переплетение и соседство биотопов разных типов. Массивы сплошных еловых лесов занимают в среднем 2,7 га, сосновых - 2,3 га.
В сообщении использованы материалы, собранные авторами в разные годы (с 1973 г. по настоящее время) в основном на территории НПВ, преимущественно в районе Валдайского стационара Института географии РАН (ИГРАН) (рис. 1), где проводили многолетние комплексные исследования разных компонентов биоты (Глазов, 2004; Кузнецов, Тишков, 2012 и др.). Использованы также литературные источники.
В 1974-1987 гг. ловушками Геро было поймано 2888 зверьков, а ловчими канавками - 1620. Подробней объемы работ и размещение ловушек в этот период даны в сводке (Шварц и др., 1992а). В 2011 и 2013 гг. зверьков ловили трапиковыми живоловками и конусами (отловлено 163 зверька 12 видов), основные места отлова совпадали с районами предшествующих многолетних исследований: стационар ИГРАН, д. Новотроицы оз. Кренье, ур. Соколово (рис. 1). Видовую принадлежность и внутривидовую структуру насекомоядных и грызунов уточняли методом хромосомного анализа с приготовлением воздушно-высушенных препаратов и их последующей обработкой с использованием различных методов окрашивания хромосом (рутинная, G- и С-окрашивание) (Seabright, 1971; Summer, 1972). Кариотипировано 6 обыкновенных бурозубок Sorex araneus Pall., 5 лесных мышовок Sicista betulina Pall., 3 подземные полевки Terricola subterraneus Selys-Longchamps, 8 полевок-экономок Alexandromys oeconomus Pall., 1 полевка надвидового комплекса M. arvalis s.L, 2
малые лесные Sylvaemus uralensis Pall, и 4 желтогорлые Sylvaemus flavicollis Melch. мыши.
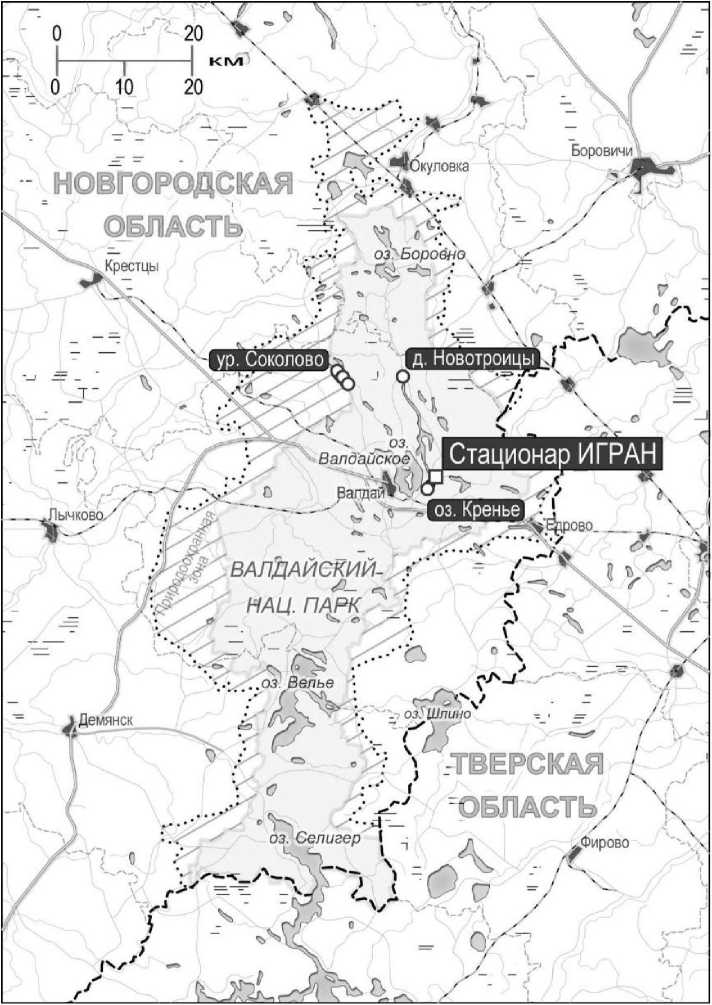
Рис. 1 . Валдайское поозерье: показаны: территория Национального парка «Валдайский» и его охранная зона, места основных многолетних работ, в т.ч. исследований 2011 и 2013 гг., гидросеть, дороги и граница между областями.
Для крупных незимоспящих млекопитающих использованы материалы зимних маршрутных учетов (ЗМУ), которые проводятся ежегодно с участием одного из авторов настоящего сообщения (В.И. Николаева) согласно Методическим рекомендациям по организации, проведению и обработке данных ЗМУ охотничьих животных в России (научно-технический совет Минсельхоза России, протокол №15 от 28 мая 2009 г.). Использованные здесь результаты получены на 21 постоянном маршруте суммарной длиной 211 км (198 км по лесным угодьям и 13 км по полям), что характеризует 122,6 тыс. га территории НПВ. Медведя учитывали по следам и встречам в летне-осенний период, а также (совместно с кабаном) на подкормочных площадках и на кормовых полях в августе-сентябре. Численность других зимоспящих млекопитающих оценивали глазомерно во время многолетней полевой работы и пеших экскурсий в разных частях НПВ.
Систематика млекопитающих дана по: Млекопитающие России ... (2012). Все латинские названия приведены в таблицах 1-4 и не дублируются в тексте. Столбец «Современные данные» в этих таблицах обобщает результаты наших наблюдений и дополняющие их литературные сведения последних десятилетий. В таблицах использованы следующие сокращения оценки численности: Оч. ред. -очень редкий, Ред. - редкий, Об. - обычный, Мн.- многочисленный, н/д - нет данных. Для материалов С.У. Строганова эти градации даны в авторской интерпретации. Мы относили к многочисленным те виды, численность которых была выше других, превышая по оценкам как минимум вдвое таковую других видов. К редким отнесены встречающиеся единично или спорадически, к очень редким -исчезающие. Остальных считали обычными. Включение вида в Красную книгу Тверской области (Викторов, Истомин, 2002) обозначено: ККТо, в Красную книгу Новгородской области (Дойникова и др., 2015) - ККНо.
Результаты и обсуждение
Изменения фауны и населения млекопитающих
Насекомоядные (Eulipotyphla). С.У.Строганов в своей статье (1936а) обсуждает 10 видов отряда насекомоядных (табл. 1). Из них, в настоящее время сведения об обитании в Валдайском поозерье тундряной бурозубки не нашли подтверждения. В Европе ее достоверные находки известны не западнее средней Мезени (Бобрецов и др., 2008). Выхухоль была отмечена только в Тверской области, находки единичны, приурочены к бассейну верховий Волги. Последняя достоверная встреча - в 1955 г. (Викторов, Истомин, 2002). Видимо, в настоящее время её в Валдайском поозерье нет. В итоге современная фауна насекомоядных Валдайского поозерья насчитывает 8 видов. Все они представлены в НПВ.
Генотипирование по микросателлитам образца ткани ежа, отловленного в 2011 г. на окраине д. Новотроицы, подтвердило обитание здесь Е. europaeus (Матвеева и др., 2016).
За 80 лет произошли существенные изменения в понимании внутривидовой структуры обыкновенных бурозубок. К настоящему времени известно 74 валидные хромосомные расы этого вида (Щипанов, Павлова, 2016), различающиеся порядком соединения диагностических акроцентриков (g-r) (Searle et al., 1991; Searle, Wojcik, 1998). На территории России описано 26 хромосомных рас (Щипанов, Павлова, 2016), многие из которых сосредоточены в области поздневалдайского оледенения или вблизи южной границы ледника (Орлов, Козловский, 2002; Орлов и др., 2007). В Валдайском поозерье и ближайших окрестностях известны расы Селигер, Санкт-Петербург, Москва (Щипанов, Павлова, 2016). Так, раса Селигер впервые найдена на южной окраине поозерья между оз. Селигер и Стерж (Bulatova et al., 2000). Позже в этой местности выявлена гибридная зона между расами Селигер и Москва (Bulatova et al., 2007), а в 50 км юго-западнее (близ оз. Лучанское, Тверская обл.) - зона контакта трех хромосомных рас (Селигер, Западная Двина и Москва) (Orlov, Borisov, 2007; Орлов и др., 2010; Bulatova et al., 2011). В окрестностях оз. Валдайское могут быть встречены расы Селигер и Санкт-Петербург (Bulatova et al., 2007; Орлов и др., 2008). К западу от Валдайского поозерья распространена раса Псков, а к северо-востоку - раса Молога, проникновение которых в Валдайском поозерье также нельзя исключать.
Таблица 1 Насекомоядные (Eulipotyphla) Валдайского поозерья
|
№ |
Русское и латинское названия |
По Строганову, 1936a |
Конец XX - начало XXI века |
Наличие вНПВ |
|
1. |
Обыкновенный ёж Erinaceus europaeus L. |
Об.* |
Подтверждено генотипированием. Ред. |
+ |
|
2. |
Европейский крот Talpa europaea L. |
Всюду |
Об. |
+ |
|
3. |
Выхухоль Desmana moschata L. |
Ред. |
Исчезла, ККТо |
- |
|
4. |
Малая бурозубка Sorex minutus L. |
Об. |
Об. |
+ |
|
5. |
Средняя бурозубка Sorex caecutiens Laxmann |
Об. |
Об. |
+ |
|
6. |
Равнозубая бурозубка Sorex is о don Turov |
Ред. |
Ред. |
+ |
|
7. |
Обыкновенная бурозубка Sorex araneus L. |
Мн. |
Мн. |
+ |
|
8. |
Крошечная бурозубка Sorex m in utissim us Zimmermann |
Оч. ред. |
Ред., ККТо |
+ |
|
9. |
Обыкновенная кутора Neomys fodiens Pennant |
Об. |
Об. |
+ |
|
- |
Тундряная бурозубка Sorex tundrensis Merriam |
Единичны |
Обитание не подтверждено |
- |
Примечание. * - здесь и далее в таблицах расшифровку сокращений см. во Введении
Нами проведены кариологические исследования обыкновенных бурозубок, отловленных в 2013 г. близ оз. Кренье (57,98° с.ш., 33,37° в.д., см. рис. 1). Было кариотипировано (G-окраска хромосом) 6 особей (все самки). Показана их принадлежность к хромосомной расе Селигер. Хромосомная формула этой расы: of, be, g, hn, ik, jl, m/q, pr, o, tu, XX/XY1Y2; стандартная номенклатура с указанием диагностических хромосом: g/o hn, ik, jl, m/q, pr (Bulatova et al., 2000). Для расы Селигер известен хромосомный полиморфизм по трем соединениям: 1) go / g, о; 2)mq / т, q; 3) pr / р, г (Bulatova et al., 2007; Павлова и др., 2007). В изученной нами выборке выявлен полиморфизм по робертсоновским транслокациям типа рг / р, г и jl / j, I. Три из исследованных особей оказались гетерозиготами только по хромосомной мутации типа рг /р, г. Их хромосомная формула (g, hn, ik, jl, mq, о, p/r), а число хромосом в диплоидном наборе 2п=23. В кариотипе четвертой самки была обнаружена гетерозиготность по двум перестройкам типа рг / р, г и jl / j, I; хромосомная формула этой особи (g, hn ,ik, j/l, mq, о, p/r,); 2n=24. Еще две особи имели 22-хромосомный кариотип и хромосомную формулу (g, hn, ik, jl, mq, о, pr).
В период 1974-1987 гг. на стационаре ИГРАН и в его окрестностях в ловчие канавки было поймано 1620 зверьков из них 1082 бурозубки и 103 куторы. Обыкновенная бурозубка занимала 1 место по численности, как среди всех отловленных этим способом зверьков (39%), так и среди бурозубок (58,4%). Средняя бурозубка была обычна: ее доля среди бурозубок - 23,8%. Для остальных бурозубок этот показатель составлял: 13.9% - малая, 3,9% -равнозубая, 0,5% - крошечная (Шварц и др., 1992а).
Крот, которого в начале века промышляли и был заметен его перепромысел (Строганов, 1936а), сейчас обычен. В разнотравнокисличных и неморальных ельниках его плотность в среднем составляла 3-4 особи на 1га, а в других типах ельников в 3-4 раза меньше. Численность резко снижается в годы летних засух, на ее восстановление уходит 2-3 года (Глазов, 2004).
Полученные материалы и комплексный анализ биотопического размещения землероек Валдайской возвышенности, морфометрии их ротового аппарата, особенностей питания, распределения кормовой базы позволили показать, что пространственно совместно обитающие виды бурозубок существенно различаются по размерам ротового аппарата, что позволяет объяснить процессы формирования многовидовых сообществ землероек Валдая, в т.ч. при смешении фаун (см. ниже). Например, восточный палеаркт - средняя бурозубка на Валдае обитает совместно с западными палеарктами - обыкновенной и малой бурозубками, но обыкновенная крупнее, а малая - мельче средней бурозубки (подробней см.: Шварц, Демин, 1986; Шварц и др., 1992а; 19926; Шварц, 2004; Shvarts, Demin, 1994). Эволюционное, зоогеографическое и биоценотическое значение различий размерности совместносуществующих бурозубок было показано и для других регионов (Шефтель, 1990; Kirkland, 1991; Sheftel, 1994; Fox, Kirkland, 1992; Hanski, 1994; Whitaker, French, 1994, Churchfield et ah, 1997; 1999 и др.).
На основе результатов исследований многих компонентов биоценозов лесов Валдая (см. выше) рассчитано, что здесь в разных типах леса бурозубки за год потребляют 24 - 245 кг/га пищи, что составляет 32 - 240% от определенного летом общего запаса почвенных и напочвенных беспозвоночных без учета их воспроизводства (Shvarts et al., 1997; Шварц, 2004). Круглогодично изымая значительную долю беспозвоночных, бурозубки стимулируют их воспроизводство, что наряду с другими факторами (Щипанов и др., 2006) позволяет считать этих насекомоядных мощными биокатализаторами процессов разложения лесной подстилки в бореальных лесах.
Рукокрылые (Chiroptera). С.У.Строганов (1936а) указывает на 9 видов (табл. 2). За последующие 80 лет этот список практически не изменился. Добавлена ночница Наттерера, которая была найдена недавно вблизи г. Боровичи и внесена в Красную книгу Новгородской области (Дойникова и др., 2015). Ночницу Брандта С.У.Строганов (1936а) рассматривал на уровне подвида усатой ночницы, обсуждая при этом, что он затруднился в определении подвидовой принадлежности исследованных ночниц, но размеры предплечья “приближают их к Myotis mystacinus brandti”. В настоящее время усатую ночницу и ночницу Брандта рассматривают как 2 самостоятельных неродственных вида (Стрелков, Бунтова, 1982; Стрелков, 1983; Крускоп, 2012), однако диагностика их затруднена. Ночница Брандта с территории Новгородской области известна по одной находке недалеко от Новгорода (Дойникова и др., 2015). Достоверных подтверждений о наличии “собственно” усатой ночницы М. mystacinus в Валдайском поозерье до сих пор нет, но судя по данным из сопредельных областей, возможно обитание обоих видов. На территории НПВ недавно обнаружены северный кожанок, водяная, прудовая и, предположительно, усатая ночницы (Попов, 2012; Шауро, 2012). Возможно обитание летучих мышей других видов. Рукокрылые остаются наименее изученным отрядом млекопитающих в фауне Валдайского поозерья (Попов, 2012).
Зайцеобразные (Lagomorpha). В перечне С.У. Строганова (19366) упомянуто 2 вида зайцев (табл. 2), что справедливо и для настоящего времени с той лишь разницей, что в те годы зайцеобразных рассматривали в составе отряда грызуны. Валдайский и Демянский районы издавна славились промыслом русаков, которых отлавливали сетями (Строганов, 19366). В наши дни русак потерял свое былое охотничье значение.
Таблица 2 Рукокрылые (Chiroptera) и зайцеобразные (Lagomorpha) Валдайского поозерья
|
№ |
Русское и латинское названия |
Ho Строганову, 1936a; 19366 |
Конец XX -начало XXI века |
Наличие в НПВ |
|
Рукокрылые (Chiroptera) |
||||
|
10. |
Ночница Наттерера Myotis nattereri Kuhl |
He отмечена |
Ред., ККНо |
- |
|
И. |
Усатая ночница Myotis mystacinus Kuhl |
Об. |
Требуется подтверждение |
+ |
|
12. |
Ночница Брандта Myotis brandtii Eversmann |
Ред., ККНо |
||
|
13. |
Водяная ночница Myotis daubentonii Kuhl |
Без оценки численности |
Без оценки численности |
+ |
|
14. |
Прудовая ночница Myotis dasycneme Boie |
Без оценки численности |
Ред., ККНо |
+ |
|
15. |
Бурый ушан Plecotus auritus L. |
Об. |
Без оценки численности |
- |
|
16. |
Рыжая вечерница Nyctalus noctula Schreber 1774 |
Об. |
Ред., ККНо |
- |
|
17. |
Лесной нетопырь Pipistrellus nathusii Keyserling et Blasius |
Об. |
Без оценки численности |
- |
|
18. |
Нетопырь карлик Pipistrellus pipistrellus Schreber |
Без оценки численности |
Требуется подтверждение |
- |
|
19. |
Северный кожанок Eptesicus nilssonii Keyserling et Blasius |
Без оценки численности |
Ред., ККНо |
+ |
|
20. |
Двухцветный кожан Vespertilio murinus L. |
Об. |
Требуется подтверждение |
- |
|
Зайцеобразные (Lagomorpha) |
||||
|
21. |
Заяц-русак Lepus europaeus Pallas |
Об. |
Об. |
+ |
|
22. |
Заяц-беляк Lepus timidus L. |
Об. |
Об. |
+ |
Заяц-беляк - обычен. При зимних учетах с 1990 по 2015 гг. в НПВ его численность варьировала с тенденцией к снижению. До 2007 г. ежегодно насчитывали более 2 тыс. зайцев. Подъемы (более 4000 ос., максимально 4580) наблюдали в 1992, 1996, 2001, 2002гг. Начиная с 2007 г., численность зайца-беляка не превышала 1703 особей. Минимум (669) пришелся на 2011 г.
Грызуны (Rodentia). С.У.Строганов (19366) упоминает 20 видов грызунов, из них серую крысу, как приблизившуюся к Валдайскому поозерью (Ржевский район), но еще не заселившую его. За 80 лет этот список изменился (табл. 3). Лесную соню, которую отмечали очень редко только в Тверской области, относят к вероятно исчезнувшей (Викторов, Истомин, 2002). Садовую соню в Тверской области также относят к вероятно исчезнувшему виду (Викторов, Истомин, 2002); в Новгородской области она еще встречается, но редко (Дойникова и др., 2015); на территории НИВ не отмечена (Николаев, 2010). Очень редка летяга. Последние достоверные находки в 2004 г. в хвойном лесу близ д. Осташево Мошенского р-на Новгородской области (Дойникова и др., 2015) и в 2012-2013 гг. в Селижаровском и Андреапольском р-нах Тверской области (две поимки в ловушки для куниц) (Комарова и др., 2015). Резко сократилась численность черной крысы. Ее еще ловили в 1979 г., но нет данных о находках этого вида в последние годы. Интересно обнаружение С.У.Строгановым обыкновенного хомяка, начавшего проникать на Валдайскую возвышенность с начала 1930-х гг. с юга и юго-востока вслед за сведением лесов. В августе 1932 г. на ячменном поле близ д. Шиблино Луковницкого района Тверской области было обнаружено около 10 особей обыкновенного хомяка (Строганов, 19366). Судя по всему, в Новгородскую область он не проникал. Современные находки обыкновенного хомяка отсутствуют, он внесен в Красную книгу Тверской области (Викторов, Истомин, 2002).
За 80 лет список грызунов Валдая пополнился пятью видами. Стали обычны интродуцированная ондатра и реинтродуцированный речной бобр. Численность бобра в НИВ, по-видимому, достигла насыщения. В последние 20 лет насчитывали 500-600 ос. Информация об обитании на территории НИВ канадского бобра (Бобров и др., 2008) не нашла достоверного подтверждения.
С.У.Строганов не упоминал лесного лемминга, хотя он был обнаружен в Новгородской области еще в 1931 г., позже (2004 г.) найден и в лесах НИВ (Дойникова, 2005; Дойникова и др., 2015). Не подозревал С.У.Строганов о существовании видов-двойников М. arvalis s. 1.: обыкновенной и восточноевропейской полевок. В 1930-е годы эти полевки (не известно: один или оба вида) были, наряду с европейской рыжей полевкой, самыми распространенными грызунами Валдая, обитая на полях, а в период уборки хлебов и зимой на гумнах, в ометах соломы и в постройках (Строганов, 19366). На севере Валдайского поозерья, где преобладают леса, за 1974-1987 гг. отловили всего 12 полевок надвидового комплекса М. arvalis s. 1. (0,27% от всех отловленных за этот период зверьков). Пять из них на сельскохозяйственных угодьях, а остальные в ельниках (кисличнике, зеленомошном и майниковом) и сосняке-брусничнике (Шварц и др., 1992а). В 2011 г. в окрестностях д. Соколово была отловлена 1 полевка этого надвидового комплекса (0,67% от всех отловленных зверьков) на поляне среди зарастающей вырубки на месте дубравы. Хромосомное типирование этой находки показало ее принадлежность к 54-хромосомной М. rossiaemeridionalis. Не исключено также обитание в Валдайском поозерье 46-хромосомной M.arvalis формы arvalis, обнаруженной юго-восточнее в соседнем ландшафтном регионе (Старицкий р-н Тверской обл.) (Булатова и др., 2010; Щипанов и др., 2010). '
Таблица 3 Грызуны (Rodentia) Валдайского поозерья
|
№ |
Русское и латинское названия |
По Строганову, 19366 |
Конец XX — начало XXI века |
Наличие в НПВ |
|
23. |
Обыкновенная летяга Pteromys volans L. |
Без оценки численности |
Ред., ККНо; ККТо |
+ |
|
24. |
Обыкновенная белка Sciurus vulgaris L. |
Об. |
Об. |
+ |
|
25. |
Лесная соня Dryomys nitedula Pallas |
Оч. ред. |
Исчезла, ККТо, в Новгродской обл. не отмечали |
- |
|
26. |
Садовая соня Eliomys quercinus L. |
Ред. |
Ред., ККНо; ККТо |
- |
|
27. |
Обыкновенный (речной) бобр Castor fiber L. |
Не было |
Об., интродукция с с 1949 г. |
+ |
|
28. |
Лесная мышовка Sicista betulina Pallas |
Без оценки численности |
Об., кариотипирована |
+ |
|
29. |
Обыкновенный хомяк Cricetus cricetus L. |
Ред., проник недавно |
Исчез, ККТо, в Новгродской обл. не отмечали |
- |
|
30. |
Лесной лемминг Myopus schisticolor Lilljeborg |
Не отмечен |
Ред., ККНо, ККТо |
+ |
|
31. |
Европейская рыжая полёвка Myodes glareolus Schreber |
Мн. |
Мн. |
+ |
|
32. |
Красная полёвка Myodes rutilus Pallas |
Ред. |
Ред. |
+ |
|
33. |
Ондатра Ondatra zibethicus L. |
Не было |
Об., интродукция с 1931 г. |
+ |
|
34. |
Водяная полёвка Arvicola amphibius L. |
Об., повсеместно |
Об. |
+ |
|
35. |
Подземная полёвка Terricola subterraneus Selys-Longchamps |
Не упомянут |
Ред., ККНо, ККТо, кариотипирована |
+ |
|
36. |
Полёвка-экономка Alexandromys oeconomus Pallas |
Об., спорадична |
Об., кариотипирована |
+ |
|
37. |
Тёмная полёвка Microtus agrestis L. |
Ред. |
Ред. |
+ |
|
38. |
Обыкновенная полёвка Microtus arvalis Pallas |
Мп. |
Об.?, требуется подтверждение |
7 |
|
39. |
Восточноевропейская полевка Microtus rossiaemeridionalis Ognev |
Ред.?, кариотипирована |
+ |
|
|
40. |
Мышь-малютка Micromys minutus Pallas 1771 |
Ред. |
Ред. |
+ |
|
41. |
Полевая мышь Apodemus agrarius Pallas |
Оч. ред. |
Ред. |
+ |
|
42. |
Малая лесная мышь Sylvaemus uralensis Pallas |
Оч. ред. |
Об. |
+ |
|
43. |
Желтогорлая мышь Sylvaemus flavicollis Melchior |
Об. |
Ред./Об. |
+ |
|
44. |
Домовая мышь Mus musculus L. |
Об. |
Об. |
+ |
|
45. |
Серая крыса Rattus norvegicus Berkenhout |
Только Ржевский р-н |
Об. |
+ |
|
46. |
Чёрная крыса Rattus rattus L. |
Об. |
Ред.?, нет современных подтверждений |
7 |
Большое значение для понимания формирования териофауны края имел прогноз, основанный на концепции «комбинативной системы экологических ниш мелких млекопитающих», а затем и доказательство обитания подземной полевки на Валдае, т.е. на сотни километров северо-восточней известного в те годы ареала этого вида (Шварц, 1985; 2004; Шварц, Замолодчиков, 1991; Шварц и др., 1992а). Наличие неизвестной ранее части ареала подземной полевки подтвердилось находками из Эстонии, Ленинградской, Псковской, Новгородской, Тверской и Брянской областей (Шварц, 1985; Загороднюк, 1992; Шварц и др., 1997; Баскевич и др., 2007 и др.). По мнению И.В. Загороднюка (1992) проникновение подземной полевки в эти районы Восточной Европы могло произойти из Балканского рефугиума, при этом многие черты ее экологии приобрели особый характер. На Валдае этот вид - типичный обитатель травянистокустарниковых сообществ неморального типа, произрастающих под пологом бореальных комплексов (Шварц, 1985). Показано, что подземная полевка из северо-западной части ареала вида, в т. ч. с Валдая, характеризуется 54-хромосомным кариотипом и относится к подвиду transvolgensis. Популяции, распространенные южнее, характеризуются 52-хромосомным кариотипом. До проведения специальных исследований было предложено рассматривать 54- и 52-хромосомные географически замещающие кариоморфы как расы, под названиями subterraneus и dacius соответственно (Загороднюк, 1989). Позднее И.В. Загороднюк (1992) предложил повысить ранг 54- и 52-хромосомных форм подземной полевки до самостоятельных географически замещающих видов. Однако исследования М.И. Баскевич с соавторами (2015) показали, что различия по хромосомам между двумя кариоморфами сводятся к одной хромосомной перестройке: робертсоновской транслокации, которой недостаточно для изоляции 54- и 52-хромосомных форм.
Интересные данные получены по генетике полевок-экономок Валдая (Баскевич и др., 2014; 2016). На территории НПВ был обнаружен хромосомный полиморфизм по робертсоновской диссоциации хромосом: две из 8 кариотипированных полевок-экономок имели 31-хромосомный кариотип, а б - стандартный 30-хромосомный, выявленный практически везде, где изучали цитогенетику этого вида (см. Воронцов и др., 1986; Баскевич и др., 2016). С помощью G-окраски хромосом установлено, что полевки-экономки из НПВ полиморфны по робертсоновской диссоциации хромосомы № 8 (РТ), обнаруженной только в гетерозиготном состоянии. Ранее существование робертсоновского полиморфизма (2п=32, 31, реже 30) было установлено только для изолятов в Фенноскандии (Fredga et al., 1980), но там найдены как гетерозиготные, так и гомозиготные носители этой перестройки. Полученные результаты могут служить аргументом в пользу существования рефугиума в ледниковой области Скандинавского щита и участия потомков сохранившихся там полевок-экономок в формирования современного населения полевок Валдая (Васкевич и др., 2014; 2016). Результаты молекулярно-генетического анализа полевок-экономок Валдая пока не получены. Предположительно их можно отнести к североевропейской филогруппе, выделенной по материалам из других регионов (Brunhoff et al., 2003; Abramson, Tikhonova, 2005).
Проведены цитогенетические исследования Sicista группы betulina (5 особей), добытых близ оз. Кренье. Использование хромосомных маркеров подтвердило принадлежность этих мышовок к 32-хромосомному виду-двойнику 5. betulina (Васкевич и др., 2012). По характеру С-окраски хромосом лесные мыщовки из НПВ незначительно отличаются от мышовок из Московской обл. и Восточных Карпат, что свидетельствует о выявленной на хромосомном уровне структурированности вида в европейской части его ареала (Васкевич и др., 2012).
В Валдайском поозерье лесная мышовка приурочена к сообществам неморального типа с богатым травяным ярусом и березнякам. Во второй половине августа молодые зверьки ловятся и в сосняках (Шварц и др., 1992а). Из валдайских ельников лесная мышовка предпочитает разнотравные, в зеленомошных и майниковых ельниках практически не встречается (Шварц и др., 1992а; Глазов, 2004). Весной, в неморальных ельниках и пойменных лесолуговых сообществах самцы лесной мышовки были очень активны, и их доля в отловах канавками была значительной (Шварц и др., 1992а). С 1974 по 1987 г. в окрестностях Валдайского стационара ИГРАН было отловлено 37 лесных мышовок ловушками Геро (1.3% от всех мелких млекопитающих отловленных ловушками) и ПО - канавками (9.3% от всех, отловленных канавками). В ельнике-кисличнике неморального типа лесная мышовка ловилась относительно регулярно, ее численность варьировала от 0,2 до 2 зверьков на 100 л-с (Шварц и др., 1992а). В отловах 2011-2013 гг. доля мышовок среди отловленных мелких млекопитающих составляла около 2%.
Лесные мыши представлены в Валдайском поозерье желтогорлой 5. flavicollis и малой лесной Sylvaemus uralensis. Последнюю ранее относили к обыкновенной лесной мыши номинативного подвида Apodemus sylvaticus sylvaticus L., 1758 (Строганов, 19366; Шварц и др., 1992а), что не соответствует современным взглядам. У кариотипированных нами 2 экз. малых лесных мышей, добытых близ оз. Кренье, выявлены хромосомные характеристики (С-окраска хромосом) (Баскевич и др., 2012), соответствующие таковым у 5. uralensis mosquensis (Орлов и др., 1996, Богданов, 2001). Численность малой лесной мыши возросла по сравнению с периодом, описанным С.У.Строгановым (19366). Если Строганов (19366) исследовал всего 1 экз. этого вида, то в отловах ловушками 1974-1987 гг. она составляла 4,4% от всех пойманных зверьков, достигая максимального обилия в неморальных ельниках (до 7 зв. на 100 л.-с. в августе 1985 г.). В наши дни она также обычна (в 2011 г. - 8% от всех отловленных грызунов).
Исследованы хромосомные характеристики 4 желтогорлых мышей, добытых в окрестностях Соколово (п=3) и оз. Кренье (п=1) (рис. 1). У всех изученных особей выявлен стандартный 48-хромосомный кариотип (Баскевич и др., 2012), характерный для большинства изученных популяций вида (Картавцева, 2002). Желтогорлая мышь из Солецкого района Новгородской области (п=1) принадлежала к северной группировке внутривидовой структуры этого вида, выделенной по митохондриальному гену первой субъединицы цитохромоксидазы ^СОГ) (Богданов и др., 2014). И, хотя ареал этой группировки пока намечен в самых общих чертах (Богданов и др., 2014), можно предположить, что мыши Валдайского поозерья относятся к ней. Желтогорлые мыши в НПВ в 1970-е годы встречались единично (1,3% от отловленных грызунов), т.е. реже, чем у С.У. Строганова. В 2011 г. этот вид был обычен (19,4% среди грызунов).
Остается многочисленной европейская рыжая полевка. В период 1974-1987 гг. она составляла 27,5% среди всех отловленных зверьков и 67,5% от отловленных ловушками грызунов. Наибольшая численность зарегистрирована в дубраве (34 ос. на 100 л.-с. в августе 1981 г.). В ельнике неморального типа ловили до 21 ос. на 100 л.-с. (август 1985), в малоблагоприятном ксеросериальном ельнике зеленомошнике - до 4 ос. на 100 л.-с. (сентябрь 1975 г.).
Обычна белка. Ее средняя численность за последнее 26-летие составляла 3650 особей, достигая максимума в 1998, 2002 и 2015 гг. (6302, 6970 и 6354 особей), а минимума в 2005-2007 гг. (700, 1260, 1655 особей).
Выросла численность серой крысы, которая в 1930-е гг. обитала только в Ржевском районе Тверской области, с 1950-х гг. стала встречаться в Новгородской области и к концу XX века широко расселилась, хотя из 3 районов, в пределах которых расположен НПВ, ее отмечали только в Валдайском (Кучерук, 1990).
Хищные (Carnivora). За 80 лет список хищных млекопитающих пополнился двумя чужеродными видами: енотовидная собака и американская норка (табл. 4). Енотовидная собака была выпущена в 1934 г. вблизи г. Вышний Волочок (Тверская область), а в 1935 г. - в
30 км юго-восточнее г. Старая Русса (Новгородская область). В первые годы после выпуска отмечали вспышки её численности и резкое снижение численности гнездящихся на земле птиц, в первую очередь тетеревиных — глухаря, тетерева, рябчика (Морозов, 1948). В настоящее время обычна: на автодорогах находят сбитых машинами особей и регулярно встречают следы.
Таблица 4
|
Хищные (Carnivora) и парнокопытные |
(Artiodactyla) Валдайского поозерья |
|||
|
№ |
Русское и латинское название |
По Строганову, 19366 |
Конец XX -начало XXI века |
Наличие вНПВ |
|
Хищные Carnivora |
||||
|
47 |
Енотовидная собака Nyctereutes procyonoides Gray |
Не было |
Об., интродукция с 1934 г. |
+ |
|
48 |
Волк Canis lupus L. |
Об. |
Ред. |
+ |
|
49 |
Обыкновенная лисица Vulpes vulpes L. |
Об. |
Об. |
+ |
|
50 |
Бурый медведь Ursus arctos L. |
Об. на севере региона |
Об. |
+ |
|
51 |
Лесная куница Martes martes L. |
Об. на севере региона |
Об. |
+ |
|
52 |
Ласка Mustela nivalis L. |
Об. |
Ред. |
+ |
|
53 |
Горностай Mustela erminea L. |
Об. |
Ред. |
+ |
|
54 |
Европейская норка Mustela lutreola L. |
Повсеместно “где больше, где меньше” |
Оч. ред., ККНо, ККТо |
+ |
|
55 |
Лесной хорь Mustela putorius L. |
Об. |
Об. |
+ |
|
56 |
Американская норка Neovison vison Schreber |
Не было |
Об., интродукция |
+ |
|
57 |
Росомаха Gulo gulo L. |
Ред. |
Заходы, нуждается в подтверждении |
7 |
|
58 |
Европейский барсук Meles meles L. |
Об., спорадичен |
Об. |
+ |
|
59 |
Речная выдра Lutra lutra L. |
Ред. |
Ред. |
+ |
|
60 |
Обыкновенная рысь Lynx lynx L. |
Об. |
Ред. |
+ |
|
Парнокопытные (Artiodactyla) |
||||
|
61 |
Кабан Sus scrofa L. |
Единичные заходы |
Об. |
+ |
|
62 |
Европейская косуля Capreolus capreolus L. |
Ред., спорадично |
Ред., ККНо |
+ |
|
63 |
Европейский лось Alces alces L. |
Оч. ред. |
Об. |
+ |
|
64 |
Благородный олень Cervus elaphus L. |
Не упомянут |
Завозят в охотхозяйства, единичные встречи |
- |
|
65 |
Пятнистый олень Cervus nippon Temminck |
Не упомянут |
В XX в. завозили в охотхозяйства |
- |
Появление американской норки в Новгородской области датируется 1970-ми гг. В Валдайский район она, по-видимому, проникла из соседнего Боровичского района, где располагались зверофермы (Туманов, 2009). В 1987 г. американская норка заселила значительную часть Валдайского района и составляла 80% среди норок обоих видов. В 2002 г. в этом районе встречали только американских норок. На территории Демянского района в 2002 г. американская норка составляла 52% среди норок, Окуловского - 10% (Туманов, 2009).
Хотя конкурентные отношения с американской норкой не служат основным фактором сокращения численности и исчезновения норки европейской (Туманов, 2009), аборигенной норки на Валдае стало заметно меньше после появления норки-пришельца. В Новгородской области еще в 1987 г. европейскую норку отмечали во всех районах, но уже в 2002 г. только в 11 из 21 (Туманов, 2009), включая Демянский и Окуловский, на территорию которых заходит НПВ. По данным И.Л. Туманова (2009) в Демянском районе предпромысловый запас европейской норки в 1987 г. оценивался в 200 особей, а к 2002 г. он сократился до 192 особей, в Окуловском районе 243 и 32 (соответственно), в Валдайском - 40 и 0 особей.
В прошлом на территорию Новгородской губернии изредка заходила росомаха: ее встречали в Боровичском уезде в конце XIX в., в конце 1930-х гг. отмечен заход росомахи в болота к востоку от оз. Селигер (Строганов, 19366). Сведения о встречах росомахи на территории области во вторую половину XX в. ограничиваются единичными устными сообщениями. Охотник В.Тиханов сообщил о заходе 2 зверей в Валдайский район (д. Байнёво) в 1965-1966 гг. От егерей и охотоведов поступала информация о заходе 2 молодых росомах в Холмский район (Дойникова, устное сообщение). Обитание в наши дни требует подтверждения.
Остается редкой выдра (30-40 особей на территории НПВ), немногочисленная практически по всему своему ареалу. Не так критически, но снизилась численность рыси (рис. 2), волка, ласки, горностая, которые были упомянуты С.У.Строгановым (19366) как обычные, а теперь редки.
К числу хищников, численность которых относительно невысока, следует отнести бурого медведя (рис. 2). Следы его жизнедеятельности зарегистрированы во всех 13 лесничествах НПВ. Южная и центральная части парка (Демянский и Валдайский районы) характеризуются оптимальным сочетанием кормовых и защитных свойств, обеспечивающих нормальное воспроизводство популяции этого вида (Дойникова, Заворовская, 2001). Поголовье бурого медведя за последние 26 лет менялось незначительно. В первой половине 1990- х гг. оно достигало 80 особей. Небольшой спад (40-41 особь) приходился на 1991, 2005 и 2006 гг. В последние годы наметился рост до 69 особей в 2015 г. (рис. 2). На медведя охотятся, но уровень изъятия небольшой (0-1 особь в год), что объясняется сложностью проведения охоты на него и стремлением многих охотников добыть крупных зверей, оставляя без внимания преобладающих в популяции особей небольшого размера.
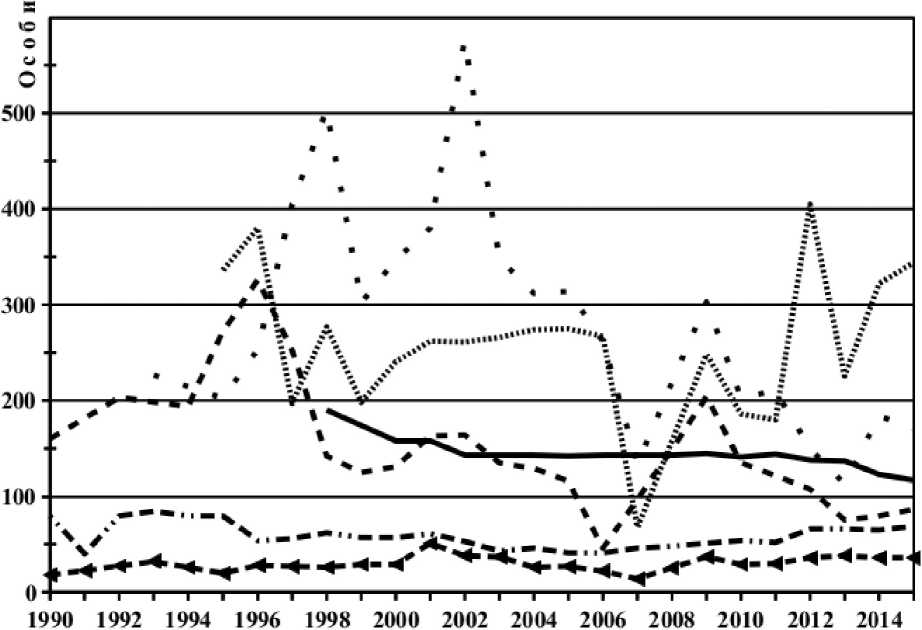
- - - - | ,. — — —. 2 ■ ■ • 3 .............4 ^^-^^- 5 ■ - ^- - 6
Рис. 2 . Многолетняя динамика численности лисицы (1), бурого медведя (2), лесной куницы (3), лесного хоря (4), европейского барсука (5) и рыси (6).
К хищникам, обычным в Валдайском поозерье, как в прошлом, так и в наши дни, относятся лисица, бурый медведь, лесная куница, лесной хорь и европейский барсук. Среди них наибольшей численности достигают лесной хорь и лесная куница (рис. 2).
Несмотря на наметившуюся новую тенденцию: появление одичавших собак и кошек, - численность хищных млекопитающих в целом сокращается.
Парнокопытные (Artiodactyla). Перечень парнокопытных за 80 лет практически не изменился (табл. 4) и включает 3 вида. В охотхозяйства периодически завозят пятнистого и благородного оленя. Благородные олени, по-видимому, убежавшие из частных охотхозяйств, изредка встречаются на территории НПВ. Их отмечали
Окуловском районе, на границе с Тверской областью, описан случай убийства браконьерами (Привалов, 2011). Однако нет данных, подтверждающих натурализацию этого вида в природе. На наш взгляд эти олени в наши дни находятся на первых фазах их интродукции, и их пока не следует включать в фауну региона.
Северного оленя (Rangifer tarandus L. 1978) С.У. Строганов (19366) считал исчезнувшим, но в 1938 г. к северу от Твери за пределами Валдайского поозерья (болото Оршинский Мох) были найдены реликтовые “острова” обитания северного оленя (Гентнер и др., 1961). Более поздних сведений об этом олене в обсуждаемом регионе нет. Несмотря на мнение, что при наблюдавшемся в XX веке отступлении северных оленей к северу, небольшие группы этих животных могли сохраниться особенно в глухих труднодоступных местах (Кузнецов, Тишков, 2012; Тишков, 2013), мы не стали включать этот вид в список фауны Валдайского поозерья XX века.
Кабан, ранее редко заходивший на Валдай, теперь стал обычен и опромышляется. Показатели среднемноголетней плотности населения кабана в Валдайском районе были многие годы одними из самых высоких в Новгородской области (30-40 особей на 100 км2), что объясняется оптимальным сочетанием лесных и полевых площадей, регулярным проведением биотехнических и охранных мероприятий (Дойникова, Чистякова, 2001). В НПВ его численность сильно колеблется в зависимости от условий зимовки, особенно от глубины снежного покрова. Она достигала максимума (427-458 особей) в 1991, 2007 и 2008 гг. и была минимальной (46-80 ос.) в 1995-1997 гг. В виду неблагоприятных условий зимовки 2010-2011 гг. (многоснежье) и эпизоотии африканской чумы свиней в сопредельных районах Тверской области, численность кабана в НПВ сократилась и в последние годы не превышает 200 голов.
Существенно, как и во многих регионах европейской части России, выросла численность лося (Кузнецов, 1983; Кузнецов, Тишков, 2012). За последние 26 лет максимальная его численность в НПВ отмечена в 1992 и 1993 гг. (576 и 625 ос.). После небольшого спада поголовья на рубеже веков (192 ос.) численность лося стала постепенно расти и достигла 386 ос. в 2015 г.
Европейская косуля остается редкой, ее численность на территории Новгородской области непостоянна. Включена в Красную книгу Новгородской области, как вид, находящийся в состоянии, близком к угрожаемому (NT) (Дойникова и др., 2015). Из-за многолетних колебаний глубины снежного покрова граница ее распространения непрерывно пульсирует. Существуя в Новгородской области на пределе своего ареала, косуля то исчезает, то появляется здесь вновь. Основные лимитирующие факторы динамики численности косуль на территории области - высота снегового покрова, браконьерство и хищничество волка (Дойникова и др., 2015). Если в I960 - начале 1970-х гг. косуль встречали единично (Кузнецов, Тишков, 2012), то с конца 1990-х гг. наметилась выраженная тенденция к росту их поголовья. Это обусловлено совокупным действием систематического контроля над численностью волка, запретом на охоту, регулярным проведением биотехнических мероприятий, а также малоснежьем зим первого десятилетия XXI века. Общая численность в Валдайском районе оценена в 10 особей. В НПВ при зимних учетах 2006-2010 гг. была отмечена только в 2008 г. Последний раз косулю в Валдайском районе видели поздней осенью 2015 г. (Кузнецов, устное сообщение). Очевидно, без постоянного внимания со стороны человека к состоянию популяции косули (подкормка, запреты на охоту, ограничение численности волка и пр.) наметившаяся тенденция роста популяции может прекратиться (Максимюк, Дойникова, 2007).
Зоогеографический анализ фауны
С.У.Строганов (19366), анализируя фауну млекопитающих, отмечает, что в ее формировании участвуют 3 группы видов: транспалеарктическая (17 видов), западные (8 видов) и восточные (4 вида) палеаркты. Летучих мышей С.У.Строганов не рассматривал. Для сравнимости данных мы тоже не включили представителей этого отряда в свой анализ. Исключена также тундряная бурозубка, обитание которой в Валдайском поозерье не подтверждено. Виды современной фауны Валдайского поозерья распределяются по этим группам так: 16, 21 и 5 видов (табл. 5). Причем группы транспалеарктов и восточных палеарктов - палеарктические условно, т.к. включают по 1 виду, распространенному и в Северной Америке (циркумбореальные): полевка-экономка и красная полевка соответственно. Еще 7 видов современной фауны - чужеродные виды, расселившиеся далеко от нативного ареала при участии человека (Бобров и др., 2008). Принципиально иной фактор формирования их современного ареала, в том числе появления на Валдае, не позволяет отнести эти виды ни к одной из перечисленных групп, и мы выделили их в новую зоогеографическую группу чужеродных видов. Из 7 видов этой группы домовая мышь и черная крыса - древние вселенцы (инвайдеры), а остальные 5 - современные. Имеется еще один вид -обыкновенный бобр, который был реинтродуцирован в Валдайском поозерье во второй половине XX века после более чем 100-летнего отсутствия на этой территории. При зоогеографической классификации видов для него принято особое решение. По
“неожиданности” воздействия на экосистемы и характеру роста численности после вселения (Завьялов, 2015; Петросян и др., 2012; 2016) бобр первые 20-40 лет после реинтродукции сравним с другими видами-интродуцентами. Однако по расположению прежнего и восстановившегося ареала - типичный западный палеаркт, к которым мы его и причислили (табл. 5).
Таблица 5 Значение западных и восточных палеарктов в фауне млекопитающих Валдая (без летучих мышей)
|
Фаунистические группы |
Виды (по Строганов, 19366) |
Современные представления |
|
Транспале аркты |
Малая и обыкновенная и бурозубки, кутора, заяц-беляк, летяга, белка, полевки темная и экономка, медведь, волк, лисица, росомаха, рысь, выдра, ласка, горностай, лось (17 видов) |
Кутора, заяц-беляк, летяга, белка, полевка-экономка*, мыши малютка и полевая, волк, лисица, медведь,ласка, горностай, росомаха, выдра, рысь, лось (16 видов) |
|
Западные палеарк-ты |
Еж, выхухоль, заяц-русак, сони лесная и садовая, рыжая полевка, желтогорлая и малая лесная мыши, лесная куница, европейская норка, лесной хорь (11 видов) |
Обыкновенный еж, европейский крот, выхухоль-, малая и обыкновенная бурозубки, заяц-русак, лесная-и садовая сони, обыкновенный бобр, лесная мышовка, обыкновенный хомяк-; рыжая, водяная, подземная, темная, обыкновенная и восточноевропейская полевки, малая лесная и желтогорлая мыши, лесная куница, европейская норка, лесной хорь, европейский барсук, европейская косуля (21 вид + 3 исчезнувших) |
|
Восточн ые палеарк-ты |
Бурозубки средняя, равнозубая и крошечная, красная полевка (4 вида) |
Средняя, равнозубая и крошечная бурозубки, лесной лемминг, красная полевка* (5 видов) |
|
Чужеродные |
Не рассматривал |
Ондатра, домовая мышь, серая и черная крысы, енотовидная собака, американская норка, кабан (7 видов) |
Примечание. * циркумбореальные виды; — исчезнувшие виды
Увеличение доли группы западных палеарктов, на наш взгляд, связано с выросшей изученностью ареалов млекопитающих. Лишь 7 видов, слагающих эту группу, практически не выходят за пределы
Европы: еж, выхухоль, садовая соня, подземная полевка, желтогорлая мышь, европейский барсук и европейская косуля. Остальные виды имеют разную степень проникновения в Сибирь. Некоторых из них (малую и обыкновенную бурозубок, темную полевку) С.У.Строганов (19366) включал в группу транспалеарктов. Мы же относим эти виды к западным палеарктам, т.е. в группу, к которой мы и С.У.Строганов включили, например, рыжую полевку, отдельные места встреч которой лежат к востоку от р. Енисей (Шварц и др., 1987).
Таблица 6 Зонально-экологический облик фауны млекопитающих Валдайского поозерья (без летучих мышей)
|
По Строганов, 19366 |
Современные представления |
|||
|
Группы |
Виды |
Группы |
Виды |
|
|
45 ^ S и о к о о |
широкорас простране иные лесные виды (преимуще ственно таежные отмечены звездочкой) |
европейский крот, бурозубки обыкновенная, средняя, крошечная и рав-нозубая*,заяц-беляк, белка, летяга, лесная мышовка, полевки рыжая, красная*, экономка*, медведь, лесная куница, барсук, росомаха*, рысь, лось, темная полевка (19 видов) |
Широкораспро с-траненные лесные виды |
заяц-беляк, летяга, белка, медведь, лесная куница, европейский барсук, рысь, лось (8 видов) |
|
Северои среднетаежные (бореальные) |
средняя, равнозубая** и крошечная бурозубки, лесной лемминг, красная и темная полевки, росомаха (7 видов); |
|||
|
Южнотаежные |
европейский крот, малая и обыкновенная бурозубки, лесная мышовка, рыжая полевка (5 видов) |
|||
|
Преимущественно широколиственнолесные |
лесная и садовая сони, лесная и желтогорлая мыши, европейская косуля (5 видов) |
Широколиствен нолесные (неморальные) |
лесная- и садовая сони, подземная полевка, малая лесная и желтогорлая мыши, европейская косуля (5 видов + 1 исчезнувший) |
|
|
2. Виды побережий различных бассейнов; болот и пойм |
выхухоль, кутора, водяная полевка, выдра, европейская норка (5 видов) |
Околоводные виды |
выхухоль-, кутора, бобр, ондатра, водяная полевка, полевка-экономка, европейская норка, американская норка, выдра, (8 видов + 1 исчезнувший) |
|
|
3. Виды открытых пространств: перелески, луга, поля и т.п., включая |
еж, заяц-русак, обыкновенный хомяк, М. arvalis s.L, полевая мышь, мышь-малютка, лесной |
Луговополевые, включая лесостепные |
обыкновенный еж, заяц-русак, обыкновенный хомяк-, обыкновенная и восточноевропейская полевки, мыши малютка и полевая, лесной хорь |
|
|
лесостепные формы |
хорь (7 видов) |
(7 видов + 1 исчезнувший) |
|
|
4. Виды, обитающие в различных ландшафтных зонах |
волк, лисица, ласка, горностай (4 вида) |
Виды, обитающие в различных ландшафтных зонах |
енотовидная собака, волк, лисица, ласка, горностай, кабан (6 видов) |
|
5. Виды, паразитирующие в жилищах |
домовая мышь, серая и черная крысы (3 вида) |
Синантропные виды |
домовая мышь, серая и черная крысы (3 вида) |
Примечание. * отмечены С.У. Строгановым как таежные; ** - занимает особое место в группе (Шварц, 1989); — исчезнувшие виды
Несмотря на изменение состава и объема зоогеографических групп, сохраняются общие закономерности композиции фауны Валдая: 1) наличие разных групп видов, формирующих фауну, 2) малая доля восточных палеарктов, 3) значительное участие западнопалеарктических видов. Наличие видов с разной историей формирования их ареалов, на наш взгляд, во многом обуславливается сохранением рефугиумов для животных разных групп в пределах ледникового щита последнего Валдайского оледенения. Анализ фауны млекопитающих Валдайского поозерья показывает, что для западных палеарктов было существенно больше путей проникновения сюда и возможностей сохранения в рефугиумах этого региона, чем для восточных. Заметим, что все 3 исчезнувших на Валдае вида, относятся к группе западных палеарктов (табл. 5), т.е. виды этой группы оказались наиболее чувствительны к воздействию комплекса современных факторов, в том числе антропогенных.
Антропогенное воздействие привело еще и к тому, что доля чужеродных видов, превысила в настоящее время в Валдайском поозерье долю восточных палеарктов. Это общая черта развития современной фауны млекопитающих России: присутствие чужеродных видов отмечено практически по всей ее территории (Хляп и др., 2011).
С.У.Строганов анализирует также экологический облик фауны Валдайской возвышенности, выделяя 5 крупных групп (табл. 6). В первой группе лесных форм им выделены 2 подгруппы: а) формы, широкораспространенные в лесах, включая характерные для фауны тайги (автор указывает 4 таежные вида - в табл. 6 они отмечены звездочкой, - но заканчивает этот перечень словами “и др.”, подчеркивая, что таежных видов больше); б) формы, обитающие преимущественно в широколиственных лесах.
Необходимо отметить, что вопрос о зонально-экологической классификации видов млекопитающих нельзя считать в настоящее время решенным. Обсуждение разных подходов (Кучерук, 1959;
Кулик, 1974; Матюшкин, 1972; 1982; Шварц, 1987; 1989; Сазонов, 2012; Равкин и др., 2013; 2014 и мн. др.) выходит за рамки нашей статьи. Здесь за основу (для сравнимости) взята классификация С.У.Строганова (табл. 6). В нее внесены минимально необходимые изменения. Прежде всего, списки видов дополнены новыми фаунистическими находками и чужеродными видами, натурализовавшимися на Валдае.
Кроме того, учтены результаты многолетних исследований мелких млекопитающих на Валдайском стационаре ИГРАН. Целенаправленный анализ этих результатов показал, что по биотопическим предпочтениям выделяются 3 группы видов: а) лесов таежного облика: средняя и крошечная бурозубки, красная и темная полевки; б) широколиственных и хвойно-широколиственных лесов: подземная и рыжая полевки, лесная и желтогорлая мыши, а также имеющая особое размещение, но тяготеющая к таким местообитаниям лесная мышовка; в) богатых влажных травянистых местообитаний, таких как пойменные луга и низинные болота (близка к предыдущей группе, особенно по распределению землероек): равнозубая бурозубка, кутора, полевка-экономка и водяная полевка (Шварц и др., 1992а).
Сказанное выше, а также обобщающая работа по анализу фауны млекопитающих лесов Северной Евразии (Кулик, 1974) позволили уточнить состав некоторых групп. Имеющиеся данные позволяют также считать целесообразным отделить виды, предпочитающие южнотаежные леса от видов северо- и среднетаежных (бореальных) лесов с одной стороны, и от видов широколиственных (неморальных) лесов с другой. Это нововведение отчасти согласуется с мнением С.В. Сазонова (2012), который, изучая зоогеографию птиц, не включал южнотаежные и подтаежные виды в северо-среднетаежную фаунистическую группу. К южнотаежным видам мы отнесли те, оптимум ареала которых лежит в южной тайге. Эти виды И.Л. Кулик (1974) включала в таежные, но на Валдае они группируются с видами широколиственных и хвойношироколиственных лесов (Шварц и др., 1992а - см. выше). Полученные нами результаты представлены в табл. 6.
Наш анализ подтверждает, что ядро фауны Валдая составляют лесные виды. В сумме на них приходится около половины от общего видового разнообразия млекопитающих Валдая: 25 из 49 анализируемых видов (исключены летучие мыши и исчезнувшие виды). Среди лесных видов преобладают широкораспространенные (8 видов). На втором месте - бореальные (7 видов). Количество южнотаежных и неморальных видов сравнялось. При этом наибольшее значение в населении млекопитающих имеют доминирующие по численности южнотаежные виды: рыжая полевка и обыкновенная бурозубка. Другие 3 южнотаежных вида: малая бурозубка, европейский крот и лесная мышовка, - обычны. Среди бореальных видов численно преобладает средняя бурозубка, занимающая вторые позиции в населении бурозубок. Роль неморальных видов невелика: лесная соня исчезла, садовая соня очень редка и встречается локально. По-видимому, это относится и к подземной полевке, хотя изученность ее распространения на Валдае крайне мала. Косуля, желтогорлая и лесная мыши более значимы, но не существенно. Это позволяет предполагать, что современные процессы на Валдае направлены в сторону повышения значения южнотаёжных млекопитающих.
При высокой обводненности Валдайского поозерья большое значение имеют околоводные виды. Их роль существенно возросла после вселения бобра, ондатры и американской норки, которые достигают более высокой численности, чем аборигенные виды этой группы.
В связи с сокращением площадей пахотных земель, покосов и зарастанием этих пространств лесом значение лугово-полевых видов в настоящее время сокращается. Повсеместное снижение численности крупных млекопитающих ведет к падению роли видов, обитающих в различных ландшафтных зонах. Из этой группы в последние годы растет лишь численность кабана, особенно там, где его специально подкармливают для охоты.
Забрасывание деревень и сокращение скота в деревнях сказывается на численности синантропных видов. Домовая мышь из маленьких деревень исчезает (Истомин, 1994, 2008), судьба черной крысы в наши дни не известна. Из новых тенденций изменения фауны - появление бродячих собак и кошек.
Заключение. В XX в. на территории Валдайского поозерья было зарегистрировано 65 видов млекопитающих. С.У.Строганов (1936а, б) в 1930-е годы дал список из 55 видов, включающий тундряную бурозубку, определение которой, возможно, было ошибочным и серую крысу, которая тогда еще не проникла в Валдайское поозерье. Отсюда фауна млекопитающих края в те годы оценивалась 53 видами млекопитающих. За последующие 80 лет были обнаружены, видимо, обитавшие здесь и ранее ночница Наттерера, лесной лемминг и подземная полевка. Усатую ночницу и ночницу Брандта, которых С.У.Строганов (1936а) считал подвидами, стали рассматривать как 2 самостоятельных вида. Изменились представления о систематике: Microtus arvalis s.L На сегодня достоверно доказано обитание в Валдайском поозерье М. rossiaemeridionalis. Наличие другого вида-двойника (М. arvalis s. str.) ожидается, но требует подтверждения. Был реинтродуцирован бобр, интродуцированы ондатра, енотовидная собака и американская норка.
Проникла и стала обычной серая крыса. Кабан, который в ЗО-е годы заходил лишь изредка, стал также обычен. Завозили европейского и пятнистого оленей, но они не натурализовались. Исчезли выхухоль и лесная соня — эти виды и раньше были редки, а также обыкновенный хомяк, начавший проникать на территорию края вслед за ростом сельскохозяйственного освоения, но несохранившийся здесь из-за современного спада земелепашества. Требуется подтверждение современного наличия на Валдае росомахи и черной крысы. Недостаточно изучены летучие мыши. Нет достоверных данных о наличии усатой ночницы, нетопыря карлика и двуцветного кожана.
Таким образом, в фауне млекопитающих рассматриваемой территории за последние десятилетия произошли следующие изменения: исчезло 3 вида, обнаружено 5 новых видов (дифференциация видов-двойников и неопознанные ранее виды), натурализовались и стали обычными 6 видов-вселенцев. Внутривидовая структура ряда видов грызунов и насекомоядных была уточнена на основе использования современных генетических подходов. В итоге - современная фауна млекопитающих Валдайского поозерья оценивается 60 видами, но как минимум для 6 видов из них требуется подтверждение обитания в наши дни.
Как и в начале века, ядро фауны млекопитающих Валдая составляют лесные виды. В сумме на них приходится около половины от общего видового разнообразия млекопитающих Валдайского поозерья. Лесные виды (без рукокрылых) представлены бореальными (7 видов), южнотаежными (5), неморальными (5) и свойственными лесам разного типа (8). Из них наибольшее значение имеют доминирующие по численности южнотаежные виды, а роль неморальных невелика. Это позволяет предполагать, что современные процессы на Валдае направлены в сторону повышения значения южнотаёжных млекопитающих.
При высокой обводненности Валдайского поозерья большое значение имеют околоводные виды. Их роль существенно возросла после появления видов-вселенцев, численность которых выше, чем аборигенных обитателей этих биотопов. Чужеродные виды не только увеличили разнообразие фауны млекопитающих края, составив от нее около 13%, но и изменили облик и функционирование околоводных экосистем.
Изменения хозяйствования в последние десятилетия ведет к сокращению значения лугово-полевых и синантропных видов. Из группы видов, обитающих в различных ландшафтных зонах (см. табл. 6), в последние годы растет лишь численность кабана, особенно там, где его специально подкармливают для охоты. Из новых тенденций изменения фауны - появление бродячих собак и кошек.
Современные генетические исследования (многообразие рас и полиморфизм обыкновенной бурозубки, полиморфизм полевки-экономки) подтверждают большую роль ледниковья в формировании фауны края. Контрастные смены ландшафтов в доисторические времена в ответ на процессы ледникового и послеледникового периодов, а в исторические еще и существенные антропогенные преобразования (Тишков, 2013), обусловили смешение и переплетение разнообразных элементов фауны, сохраняющееся и в наши дни. Современные процессы увеличения облесенности края поддерживают главенство лесного ядра фауны и, прежде всего, южнотаежных элементов. За счет видов-вселенцев существенно возросло значение околоводных млекопитающих.
Авторы искренне благодарят всех, принимавших участие в полевых работах на территории Валдайского поозерья, а также рецензента, предложения которого позволили улучшить содержание статьи.
THE FAUNA, ECOLOGY AND ZOOGEOGRAPHY OF MAMMALS IN VALDAI LAKELAND: RETROSPECTIVE AND CURRENT
STATUS
L.A. Khlyap1, E.A. Shvarts2, M.I. Baskevich1, V.I. Nikolaev3, A.A. Tishkov4, O.A.Leonteva5, E.V. Cherepanova1, P.M. Glazov4
^evertsov Institute of Ecology and Evolution RAS, Moscow
-
2 Russia World Wide Fund, Moscow
-
3 National Park «Valdaysky», Valday Geographical Institute RAS, Moscow "Lomonosov Moscow State University, Moscow
A revision of modern mammal fauna of the Valdai Lakeland is carried out. Its full list is provided. Transformations of theriofauna and zoogeography from 1930s to the present day are analyzed. The results of the ecological research of the last decades, including winter monitoring data on large mammals, are presented. New information about sibling species and intraspecific forms of Rodentia and Eulipotyphla was obtained by the modern genetic techniques. These data confirm the important role of the last glaciation in the fauna and species formation. The current stage of theriofauna formation is characterized by the appearance of the new zoogeographical group of alien species. Alien species did not only increase the diversity of mammal fauna of the Valdai Lakeland, but also changed functioning of riparian ecosystems.
Об авторах:
ХЛЯП Людмила Айзиковна - кандидат биологических наук, старший научный сотрудник лаборатории сохранения биоразнообразия и использования биоресурсов, ФГБУН Институт проблем экологии и
Хляп Л.А. Фауна, экология и зоогеография млекопитающих Валдайского поозерья: ретроспектива и современное состояние / Л.А. Хляп, Е.А. Шварц, М.И. Васкевич, В.И. Николаев, А.А. Тишков, О.А. Леонтьева, Е.В. Черепанова, П.М. Глазов // Вести. ТвГУ. Сер.: Биология и экология. 2017. № 1. С. 125-159.
Список литературы Фауна, экология и зоогеография млекопитающих Валдайского поозерья: ретроспектива и современное состояние
- Баскевич М.И., Крысанов Е.Ю., Малыгин В.М., Сапельников С.Ф. 2007. Новые данные по хромосомной изменчивости подземной полевки Microtus (Terricola) subterraneus (Rodentia, Arvicolidae) на территории России и Украины//Зоол. журн. Т. 86. Вып. 3. С. 369-376.
- Баскевич М.И., Потапов С.Г., Миронова Т.А., Хляп Л.А., Окулова Н.М. 2015. Структура вида у полевок подрода Terricola (Rodentia, Arvicolinae, Microtus) фауны России в свете кариологических, молекулярных и краниологических данных//Структура вида у млекопитающих. Матер. Конф. (Москва 21-23 окт. 2015). М.: Товарищество научных изданий КМК. С. 14.
- Баскевич М.И., Хляп Л.А., Шварц Е.А. 2012. Хромосомные подходы в изучении таксономического и генетического разнообразия грызунов Валдайской возвышенности//Матер. межд. конф. "Экология, эволюция и систематика животных" (Рязань, 13-16 ноября 2012). Рязань: "Голос губернии". С. 191-193.
- Баскевич М.И., Хляп Л.А., Шварц Е.А. 2014. Хромосомный полиморфизм в периферической популяции полевки-экономки Microtus oeconomus (Rodentia, Arvicolinae) из Валдайского национального парка // Доклады Академии наук. Т. 454, № 2. С. 227-231. (перевод: Baskevich M.I., Khlyap L.A., Shvartz E.A., 2014. Chromosomal polymorphism in the peripheral population of the root vole Mirotus oeconomus (Rodentia, Arvicolinae) of the Valdai National Park // Doklady Biological Sciences, V. 454. P. 29-33.) DOI: 10.1134/S0012496614010062
- Белоновская Е.А., Кренке-мл. А.Н., Тишков А.А., Царевская Н.Г. 2014. Природная и антропогенная фрагментация растительного покрова Валдайского поозерья//Изв. РАН. Сер. геогр., № 5. С. 67-82.
- Бобрецов А.В., Купрянова И.Ф., Петров А.Н. Демидова Т.Б., Щипанов Н.А. 2008. Европейская лесная форма тундряной бурозубки (Sorex tundrensis)//Зоологический журнал. Т. 87. Вып. 7. С. 841-849.
- Бобров В.В., Варшавский А.А., Хляп Л.А. 2008. Чужеродные виды млекопитающие в экосистемах России. Ред. Дгебуадзе Ю.Ю., Неронов В.М. М.: Товарищество научных изданий КМК. 232 с.
- Богданов А.С. 2001. Хромосомная дифференциация популяций малой лесной мыши, Sylvaemus uralensis в восточной части ареала вида//Зоологический журнал. Т. 80. Вып. 3. C. 331-342.
- Богданов А.С. Стахеев В.В. Зыков А.Е. Окулова Н.М. Миронова Т.А. 2014. Внутривидовая дифференциация желтогорлой мыши Sylvaemus flavicollis в восточной части ареала: результаты изменчивости фрагмента митохондриального гена первой субъединицы цитохромоксидазы//Зоологические исследования регионов России и сопредельных территорий. Н. Новгород: Изд-во гос. Ун-та им. К. Минина. С. 258-264.
- Булатова Н.Ш., Голенищев Ф.Н., Ковальская Ю.М., Емельянова Л.Г., Быстракова Н.В. 2010. Цитогенетическое изучение парапатрической зоны контакта двух 46-хромосомных форм обыкновенной полевки в Европейской России//Генетика. Т. 46. № 4. С. 502-508.
- Викторов Л.В., Истомин А.В. 2002. Млекопитающие Mammalia//Красная книга Тверской области. Ред. А.С. Сорокин. Тверь: ООО "Вече Твери", ООО "Издательство АНТЭК". C. 153-161.
- Воронцов Н.Н., Ляпунова Е.А., Боескоров Г.Г., Ревин Ю.В. 1986. Стабильность кариотипа полевки-экономки (Microtus oeconomus) в центральной части ареала и история становления современного ареала вида//Зоологический журнал. Т. 65. № 11. С. 1705-1715.
- Гептнер В.Г., Насимович А.А., Банников А.Г. 1961. Млекопитающие Советского Союза. Т. 1. Парнокопытные и непарнокопытные. Москва. Изд-во «Высшая школа». 776 с.
- Глазов М.В. 2004. Роль животных в экосистемах еловых лесов. М.: Пасьва. 239 с.
- Дойникова О.Ю. 2005. Новый вид родентофауны национального парка "Валдайский"//Уч. зап. ин-та сельского хоз-ва и природ. ресурсов НовГУ. Великий Новгород, Т. 13. Вып. 2. С. 134-137.
- Дойникова О.Ю., Заворовская Е.В. 2001. Биотопическая приуроченность и половозрастная структура популяции медведей Ursusarctos в Новгородской области//Северо-Запад России: взаимодействие общества и природы. Ч. 1. Псков. С. 171-172.
- Дойникова О.Ю., Завьялов Н.А., Попов И.Ю., Хляп Л.А. 2015. Млекопитающие//Красная книга Новгородской области/Ред. Веткин Ю.Е., Гельтман Д.В., Литвинова Е.М., Конечная Г.Ю., Мищенко А.Л. С.-Пб.: Дитон. С. 145-158.
- Дойникова О.Ю., Чистякова А.Н. 2000. География и численность ресурсов кабана Sus scrofa в Новгородской области//Социальные и экологические проблемы Балтийского региона. Мат-лы общественно-научной конф. (Псков, 2-4 ноября 2000 г.). Псков. С. 167-169.
- Жизнь и деятельность Сергея Ульяновича Строганова. 1962.//Строганов С.У. Звери Сибири. Хищные. Отв. ред. В.Г. Гептнер. М.: "Изд-во академии наук СССР". С. 5-8.
- Завьялов Н.А., 2015. Средообразующая деятельность бобра (Castor fiber L.) в европейской части России/Труды гос. природного заповедника "Рдейский". Вып. 3. Великий Новгород. 320 с.
- Загороднюк И.В. 1989. Таксономия, распространение и морфологическая изменчивость полевок рода Terricola Восточной Европы//Вестник зоологии. № 5. С. 3-14.
- Загороднюк И.В. 1992. Географическое распространение и уровни численности Terricola subterraneus на территории СССР//Зоологический журнал. Т. 71. Вып. 2. С. 86-97.
- Истомин А.В. 1994. Грызуны населенных пунктов малоосвоенной территории водораздела Волги и Западной Двины//Синантропные грызуны. Матер. 2 совещ., Иваново, 1993. Характер поведения. Особенности обитания в постройках человека. Методы учетов. Пути ограничения численности. М.: без издательства. С. 124-129.
- Истомин А.В. 2008. Мелкие млекопитающие в региональном экологическом мониторинге (на примере Каспийско-Балтийского водораздела). Псков: без издательства. 278 с.
- Картавцева И.В. 2002. Кариосистематика лесных и полевых мышей. Rodentia: Muridae. Владивосток: «Дальнаука». 141 с.
- Комарова В.Н., Палкова Т.С., Шмитов А.Ю. 2015. Новые флористические и фаунистические находки редких и охраняемых видов в Тверской области//Изучение и охрана природного и исторического наследия Валдайской возвышенности и сопредельных регионов: Мат-лы межрегион. научно-практ. конф., посвященной 25-летию национального парка "Валдайский", г. Валдай, 24-25 апреля 2015 г. Вышний Волочек. С. 191-197.
- Крускоп С.С. 2012. Отряд/Order Chiroptera Blummenbach, 1779//Млекопитающие России: систематико-географический справочник. Ред. И.Я. Павлинов, А.А. Лисовский М.: Т-во науч. изд. КМК. C. 73-126.
- Кузнецов Г.В. 1983. Влияние лося на лесную растительность южной тайги//Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 88. Вып. 1. С. 28-35.
- Кузнецов Г.В., Тишков А.А. 2012. Зоогеографические исследования на территории Валдайской возвышенности (к 40-летию Валдайского стационара Института географии РАН)//Известия РАН. Серия географическая. № 2. С. 82-95.
- Кулик И.Л. 1974. Сравнительный анализ фаунистических комплексов млекопитающих (Mammalia) лесной части Северной Евразии//Териология. Т. 2. Новосибирск. С. 151-161.
- Кучерук В.В. 1959. Степной фаунистический комплекс млекопитающих и его место в фауне Палеарктики//География населения наземных животных и методы его изучения. М.: Изд-во АН СССР. С. 45-87.
- Кучерук В.В. 1990. Ареал//Серая крыса: Систематика, экология, регуляция численности. Ред. В.Е. Соколов, Е.В. Карасева. М.: Наука. С. 34-84.
- Максимюк Н.Н., Дойникова О.Ю. 2007. Косуля европейская как редкий вид ресурсных копытных Новгородской области//Известия Санкт-Петербургского гос. аграрного университета. № 6. С. 90-93.
- Матвеева Е.И., Белоконь М.М., Белоконь Ю.С., Лебедев В.С., Рутовская М.В. 2016. Популяционно-генетическое разнообразие и гибридизация обыкновенных ежей европейской части России и Западной Сибири//Териофауна России и сопредельных территорий. Междунар. совещание (X съезд Териол. общества при РАН). М.: Товарищество научн. изданий КМК. С. 255.
- Матюшкин Е.Н. 1972. Смешанность териофауны Уссурийского края: ее общие черты, исторические корни и современные проявления в сообществах Среднего Сихотэ-Алиня//Исследования по фауне Советского Союза (млекопитающие). Сб. тр. Зоологического музея МГУ. Т. 13. М.: МГУ. С. 86-144.
- Матюшкин Е.Н. 1982. Региональная дифференциация лесной фауны Палеарктики в прошлом и настоящем. В кн.: Теоретические и прикладные аспекты биогеографии. М.: Наука. С. 59-80.
- Млекопитающие России: систематико-географический справочник. 2012. Ред. И.Я. Павлинов, А.А. Лисовский. М.: Т-во научн. изд. КМК. 636 с.
- Морозов В.Ф. 1948. Результаты акклиматизации енотовидной собаки (Nyctereutes procyonoides) и в Ленинградской и Новгородской областях//Труды Всесоюз. НИИ охотничьего промысла Министерства заготовок СССР. Л., Т. 8. С. 111-124.
- Николаев В.И. 2010. Предварительный список видов позвоночных животных национального парка "Валдайский" (по состоянию на 2010 год)//Труды нац. парка "Валдайский": юбил. сб. к 20-летию Валдайского национального парка. Вып. 1. Ред. Е.М. Литвинова. СПб.: без издательства. С. 413-422.
- Орлов В.Н., Борисов Ю.М., Ирхин С.Ю., Ковалева А.А. 2010. Особенности зоны контакта трех хромосомных рас обыкновенной бурозубки Sorex araneus L. (Mammalia) показатель конкуренции популяций//Экология. № 6. С. 459-463.
- Орлов В.Н., Козловский А.И. 2002. О роли ледниковых эпох в формировании хромосомного полиморфизма обыкновенной бурозубки Sorex araneus p. (Insectivora, Mammalia)//Доклады РАН. Т. 386. С. 423-426.
- Орлов В.Н., Козловский А.И., Балакирев А.Е., Борисов Ю.М. 2007. Эндемизм хромосомных рас обыкновенной бурозубки Sorex araneus L. (Insectivora, Mammlia) и возможность сохранения рефугиумов в области покровного оледенения поздневалдайской эпохи//Доклады Академии наук. Т. 416. № 6. С. 1-4.
- Орлов В.Н., Козловский А.И., Балакирев А.Е., Борисов Ю.М. 2008. Процессы фиксации метацентрических хромосом в популяциях обыкновенной бурозубки Восточной Европы//Генетика. Т. 44. № 6. С. 581-593.
- Орлов В.Н., Козловский А.И., Наджафова Р.С., Булатова Н.Ш. 1996. Хромосомные диагнозы и место генетических таксонов в эволюционной классификации лесных мышей Sylvaemus Европы (Apodemus, Muridae, Rodentia)//Зоологический журнал Т. 75. Вып. 1. С. 88-102.
- Павлова C.B., Булатова H.Ш. Щипанов H.A. 2007. Цитогенетический контроль гибридной зоны двух хромосомных рас Sorex araneus перед сезоном размножения//Генетика. T. 43. № 12. C. 1619-1626.
- Попов И.Ю. 2012. К фауне летучих мышей Новгородской области//Полевой сезон -2011: Исследования и природоохранные действия на особо охраняемых территориях. Ред. Е.М. Литвинова. Валдай: Печатный двор "Великий Новгород". С. 65-67.
- Петросян В.Г., Голубков В.В., Горяйнова З.И., Завьялов Н.А. Альбов С.А. 2012. Опыт моделирования динамики численности речного бобра (Castor fiberL.) в бассейне малой реки Таденки притока Оки (Приокско-террасный заповедник) // Российский журнал биологических инвазий. № 3. С. 44-59. (перевод: PetrosyanV.G., GolubkovV.V., GoryainovaZ.I., Zav’yalovN.A., Al’bovS.A. et al., 2013. Modeling of the Eurasian beaver (Castor fiberL.) population dynamics in the basin of a small Oka River tributary, the Tadenka river (Prioksko-Terrasny nature reserve) // Russian Journal of Biological Invasions, Vol. 4, No. 1. P. 45-53.)
- Петросян В.Г., Голубков В.В., Завьялов Н.А., Горяйнова З.И., Дергунова Н.Н., Омельченко А.В., Бессонов С.А., Альбов С.А., Марченко Н.Ф., Хляп Л.А. 2016. Закономерности динамики численности речного бобра (Castor fiberL.) после его вселения в особо охраняемые природные территории Европейской части России // Российский журнал биологических инвазий. № 3. С. 66-89. (Перевод: Petrosyan V.G., Golubkov V.V., Zavyalov N.A., Goryainova Z.I., Dergunova N.N., Omelchenkо A.V., Bessonоv S.A., Albov S.A., Marchenko N.F., Khlyap L.A. Patterns of population dynamics of Eurasian beaver (Castor fiberL.) after reintroduction into nature reserves of European part of Russia // Russian Journal of Biological Invasions, 2016, Vol. 7, No. 4. P. 355-373.)
- Привалов К. 2011. Убийство оленя браконьерами на Валдае осталось безнаказанным//Ваши новости. Новгородская интернет-газета. . Режим доступа: http://vnnews.ru/proishestviai/11252-ubijstvo-putinskogo-olenya-brakonerami-na-valdae-novgorodskaya-oblast-ostal.html. Дата обпащения: 26.08.2011.
- Равкин Ю.С., Богомолова И.Н., Николаева О.Н. 2013. Териофаунистическое районирование Северной Евразии//Сиб. экол. журн. Т. 20. № 1. С. 111-121.
- Равкин Ю.С., Богомолова И.Н., Николаева О.Н. 2014. Районирование Северной Евразии по фауне наземных позвоночных и классификация их по сходству распространения//Сиб. экол. журн. Т. 21. № 2. С. 163-181.
- Сазонов С.В. 2012. Обновленная классификация типов фауны и фаунистических групп птиц для запада евразийской тайги//Труды Карельского научн. центра РАН. № 1. С. 70-85.
- Стрелков П.П. 1983. Усатая ночница (Myotis mystacinus) и ночница Брандта (Myotis brandti) в СССР и взаимоотношения этих видов. Сообщение 2//Зоологический журнал. Т. 62. Вып. 2. С. 259-271.
- Стрелков П.П., Бунтова Е.Г. 1982. Усатая ночница (Myotis mystacinus) и ночница Брандта (Myotis brandti) в СССР и взаимоотношения этих видов. Сообщение 1//Зоологический журнал. Т. 61. Вып. 8. С. 1227-1242.
- Строганов С.У. 1936а. Фауна млекопитающих Валдайской возвышенности//Зоологический журнал. Т. 15. Вып. 1. С. 128-142.
- Строганов С.У. 1936б. Фауна млекопитающих Валдайской возвышенности III//Зоологический журнал. Т. 15. Вып. 3. С. 520-559.
- Тишков А.А. 2013. Тысячелетняя история изменения фауны млекопитающих Валдайского поозерья//Вопросы географии. Вып. 136. С. 385-412.
- Туманов И.Л. 2009. Редкие хищные млекопитающие России (мелкие и средние виды). СПб: ООО "Бранко". 448 с.
- Хляп Л.А., Варшавский А.А., Бобров В.В. 2011. Разнообразие чужеродных видов млекопитающих в различных регионах России // Российский журнал биологических инвазий. № 3. С. 79-88. (перевод: Khlyap L. A., Warshavsky A. A., Bobrov V. V. 2011, Diversity of Alien Mammal Species in Different Regions of Russia // Russian Journal of Biological Invasions, Vol. 2, No. 4, pp. 293-299)
- Шауро Т.Н. 2012. Выявление рукокрылых на территории Валдайского национального парка//Полевой сезон -Исследования и природоохранные действия на особо охраняемых территориях. Ред. Е.М. Литвинова. Валдай: Печатный двор "Великий Новгород". С. 68-69.
- Шефтель Б.И. 1990. Анализ пространственного распределения землероек в средней Енисейской тайге//Экологическая ординация и сообщества. М.: Наука. С. 15-32.
- Шварц Е.А. 1985. О распространении и биологии европейской подземной полевки на севере//Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 90. Вып. 3. С. 25-31.
- Шварц Е.А. 1987. Зональный лесостепной териокомплекс и зоогеографические тенденции, связанные с антропогенным преобразованием территории европейской части СССР//Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных. Тез. Всесоюзного совещания. Ч. 1. М.: без издательства. С. 53-55.
- Шварц Е.А. 1989. Формирование фауны мелких грызунов и насекомоядных таежной Евразии//Фауна и экология грызунов. М.: МГУ. Вып. 17. С. 115-143.
- Шварц Е.А. 2004. Сохранение биоразнообразия: сообщества и экосистемы. М.: Т-во научных изданий КМК, 2004. 112 с.
- Шварц Е.А., Демин Д.В. 1986. О факторах сосуществования родственных видов в местах симпатрии ареалов (на примере Soricidae)//Доклады Академии наук. Т. 289. № 1. С. 253-256.
- Шварц Е.А., Демин Д.В., Глазов М.В., Замолодчиков Д.Г. 1992а. Организация населения землероек лесов умеренного пояса Евразии и влияние на нее структуры населения почвенной мезофауны//Доклады Академии наук. Т. 322. № 2. С. 427-431.
- Шварц Е.А. Демин Д.В., Замолодчиков Д.Г. 1992б. Экология сообществ мелких млекопитающих лесов умеренного пояса (на примере Валдайской возвышенности). М.: Наука. 127 с.
- Шварц Е.А., Замолодчиков Д.Г. 1991. Комбинативная система экологических ниш как способ отражения структуры населения мышевидных грызунов природных экосистем Валдайской возвышенности//Зоологический журнал. Т. 70. Вып. 4. С. 113-124.
- Шварц Е.А., Коршунова Е.Н., Хейфец О.А., Воеводин П.В. 1997. Фауна мелких млекопитающих заповедника "Брянский лес"//Вестник зоологии. Т. 31. Вып. 3. С. 25-32.
- Шварц Е.А., Шефтель Б.И., Жуков М.А. 1987. Закономерности распространения рыжей полевки на востоке ареала//Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 92. Вып. 2. С. 17-26.
- Щипанов Н.А., Александров Д.Ю., Александрова А.В. 2006. Распространение спор микроскопических грибов мелкими млекопитающими//Зоологический журнал. Т. 85. Вып. 1. С. 101-113.
- Щипанов Н.А., Булатова Н.Ш., Павлова С.В., Щипанов А.Н. 2009. Обыкновенная буроубка (Sorex araneus) -модельный вид эколого-эволюционных исследований//Зоологический журнал. Т. 88. Вып. 8. С. 975-989.
- Щипанов Н.А.,Купцов А.В.,Калинин А.А., Демидова Т.Б., Олейниченко В.Ю. 2010. Мелкие млекопитающие юго-востока Тверской области. Сообщение 1. Фауна и биотопическое распределение//Сибирский экологический журнал, (Новосибирск). Т.17. №5. С.799-806.
- Щипанов Н.А., Павлова С.В. 2016. Многоуровневая подразделенность в группе видов "araneus" рода Sorex. 1. Хромосомная дифференциация//Зоологический журнал. Т. 95. Вып. 2. С. 216-233.
- Abramson N.I., Tikhonova E.P. 2005. Reevaluation of taxonomic structure of the root vole (Microtus oeconomus Pallas, 1776, Rodentia, Arvicolidae) from the territory of the former USSR based on evidence from craniometric and molecular data//Russian J. Theriology. V. 4. № 1. P. 63-73.
- Brunhoff C., Gakbreath K.E., Fedorov V.B., Cook J.A., Jaarola M.J. 2003. Holoarctic phylogeography on the root vole Microtus oeconomus: implications for late Quaternary biogeograpgy of high latitudes//Molecular Ecology. V. 12, № 4. P. 957-968.
- Bulatova N., Jones R.M., White T.A., Shchipanov N.A., Pavlova S.V., Searle J.B. 2011. Natural hybridization between extremely divergent chromosomal races of the common shrew (Sorex araneus, Soricidae, Soricomorpha): hybrid zone in European Russia//Journal of Evolutionary Biology. V. 24. № 3. P. 573-586.
- Bulatova N.Sh., Searle J.B., Bystrakova N., Nadjafova R., Shchipanov N., Orlov V. 2000. The diversity of chromosome races in Sorex araneus from European Russia//Acta Theriol. V. 45. Suppl. 1. P. 33-46.
- Bulatova N., Shchipanov N., Searle J.B. 2007. The Seliger -Moscow hybrid zone between chromosome races of common shrews -an initial description//Russian J. Theriology. V. 6. № 1. P. 111-116.
- Churchfield S., Nesterenko V.A., Shvarts E.A. 1999. Food niche overlap and ecological separation amongst six species of coexisting forest shrews (Insectivora: Soricidae) in the Russian Far East//Journal of Zoology. V. 248. P. 349-359.
- Churchfield S., Sheftel B.I., Moraleva N.V., Shvarts E.A. 1997. Habitat occurrence and prey distribution of a multispecies community of shrews in the Siberian taiga//Journal of Zoology. V. 241. P. 55-71.
- Fox B.J., Kirkland G.L. Jr. 1992. An assembly rule for functional groups applied to North American soricidae communities//Journal of Mammology. V. 73. P. 491-503.
- Fredga K., Persson A., Stenseth Ch. 1980. Centric fission in Microtus oeconomus. A chromosome study of isolated populations in Fennoscandia//Hereditas. V. 92. № 2. P. 209-216.
- Hanski I. 1994. Population biological consequence of body size in Sorex//Advances in the biology of shrews. Merritt J.F., Kirkland G.L. Jr., and Rose R.K. (Eds). Carnegie Museum of Natural History Special Publication 18. P. 15-26.
- Kirkland G.L. 1991. Competition and coexistence in shrews (Insectivora, Soricidae)//The biology of Soricinae. Special Publication The Museum of Sowthwestern Biology. Findley J.S., Yates T.L. (Eds). University of New Mexico. Albuquerque. P. 15-22.
- Matuszkevicz J.M., Bielonowska E., Kowalska A., Carewska N., Baranowski J. 2014. Biaiowieїa Forest (NE-Poland) and Valday (NW-Russia) -biogeographical characteristics of eutrophic deciduous forests//Quaestiones Geographicae, № 33(3). P. 111-123.
- Orlov V.N., Borisov Yu.M. 2007. Chromosome races of the common shrew Sorex araneus L., 1758 (Mammalia: Insectivora) from the south part of Valdai Heights (Russia)//Compar. Cytogenet. V. 1. № 2. P. 101-106.
- Seabright M. 1971. A rapid banding technique for human chromosomes//Lancet. V. 11. P. 971-972.
- Searle J. B., Fedyk S., Fredga K., Hausser J., Volobouev V.T. 1991. Nomenclature for the chromosomes of the common shrew (Sorex araneus)//Mйmoires de la Sociйtй Vaudoise des Sciences naturelles. V. 19. P. 13-22.
- Searle J.B., Wуjcik J.M. 1998. Chromosomal evolution: the case of Sorex araneus. Survey of hybrid zones. In: Wуjcik J.M. and Wolsan M. (eds.), Evolution of shrews. Mammal Research Institute Polish Acad. Sci., Biaiowieїa. P. 243-253.
- Sheftel B.I. 1994. Spatial distribution of nine species of shrews in the central Siberian taiga//Advances in the biology of shrews. Merritt J.F., Kirkland G.L. Jr.,Rose R.K. (Eds). Carnegie Museum of Natural History Special Publication 18. Pittsburgh, PA. P. 45-56.
- Shvarts E.A., Demin D.V. 1994. Community organization of shrews in temperate zone forests of Northwestern Russia//Advances in the Biology of Shrews. Eds.: J.F. Merritt, G.L. Kirkland Jr., R.K. Rose. Special Publication of the Carnegie Museum of Natural History № 18. Pittsburgh. P. 57-66.
- Shvarts E.A., Chernychev N.V., Popov I.Yu. 1997. Do shrews impact on soil invertebrates in Eurasian forests?//Ecoscience. V. 4. № 2. P. 158-162.
- Sumner A.T. 1972. A simple technique for demonstrating centromeric heterochromatin//Exp. Cell Res. V. 75. P. 304-306.
- Whitaker J.O., French T.W. 1994. Foods of six species of sympatric shrews from New Brunswick//Canadian Journal of Zoology. V. 62. P. 622-626.
