Фауна, население и особенности ландшафтного распределения дождевых червей (Oligochaeta, Lumbricidae) пояса остепненных лугов Центрального Кавказа
 пояса остепненных лугов Центрального Кавказа")
Автор: Рапопорт Ирина Борисовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 4-2 т.17, 2015 года.
Бесплатный доступ
Исследованы видовой состав, структура населения и ландшафтное распределение дождевых червей пояса остепненных лугов эльбрусского варианта поясности. Показано, что в остепненных лугах превалирующим фактором, формирующим структуру населения люмбрицид, является характер распределения влаги, которая аккумулируется в отрицательных формах рельефа и на пологих теневых склонах, где отмечены максимальные показатели обилия дождевых червей. Число видов, отмеченных в поясе остепненных лугов, меньше, чем в поясах луговых степей и широколиственных лесов. Распределение видов по стациям обитания зависит от ландшафтного положения биогеоценоза и ориентации ущелья относительно основных потоков переноса влаги. Пояс остепненных лугов от ниже лежащих поясов отличается более «кавказским» типом населения дождевых червей. Доминируют в большинстве сообществ собственно почвенные люмбрициды. Преобладание собственно почвенных видов обусловлено общей ксерофитизацией ландшафтов эльбрусского варианта поясности. В горных степях на высотах от 1000 м над ур. м. отмечено замещение космополитного A. rosea автохтонным викарирующим видом D. tellermanica, в остепненных лугах - собственно почвенной формой D. schmidti.
Дождевые черви, фауна, население, экология, ландшафтное распределение, пояс остепненных лугов, центральный кавказ
Короткий адрес: https://sciup.org/148203891
IDR: 148203891 | УДК: 591.9:595.14
Fauna, population and peculiarities of landscape distribution in earthworms (Oligochaeta, Lumbricidae) of the steppe meadow belt in the Central Caucasus
The species composition, population structure and landscape distribution of earthworms in the steppe meadow belt from the elbrusskiy variant of vertical zonation, are studied. It is shown that the character of moisture distribution accumulating in negative landforms and on gentle shady slopes where maximum parameters of earthworm abundance are registered, is a prevailing factor for developing the population structure of Lumbricidae. The number of species from the steppe meadow belt is less than in the belts of meadow steppes and broad-leaved forests, but it exceeds the species abundance of the subalpine belt. The species distribution in their stations depends on landscape location of the biogeocenosis and gorge orientation relative to basic moisture transfer flows. The steppe meadow belt differs from subjacent belts by the more “caucasian” type of the earthworm population. Proper soil Lumbricidae dominate in most communities. Prevalence of proper soil species which are more adapted to survive under adverse climatic conditions is associated with total xerophytization of landscapes in the elbrusskiy variant of vertical zonation. In mountain steppes at altitude from 1000 m a.s.l. cosmopolitan A. rosea is substituted by autochthonous vicarious D. tellermanica, and by proper soil D. schmidti in steppe meadows.
Текст научной статьи Фауна, население и особенности ландшафтного распределения дождевых червей (Oligochaeta, Lumbricidae) пояса остепненных лугов Центрального Кавказа
более 40 биотопов с повторностью 5-8 проб. Диапазон высот: на Джинальском хр., бас. р. Южная Золка (900-1417 м над ур. м.); в ущ. р. Тызыл и Гунделен (850-1262 м над ур. м.); на хр. Качкор-таш, бас. Р. Чегем (1250-1260 м над ур. м.). При сборе материала использовали метод ручной разборки почвы [6] из проб 25х25 см2. Раскопки проводили на глубину встречаемости дождевых червей. Таксономический статус люмбрицид и характеристика морфо-экологических форм приведены согласно литературным сводкам [7, 8]. Обработка полученных данных выполнена в программе Statistica 10 . 0.
Упрощенная схема деления ландшафтных позиций на элювиальные, транзитные и аккумулятивные [9 и мн. др.], принятая в почвенной зоологии, на наш взгляд, в горах должна быть дополнена экспозицией склонов. Ориентация ущелий северного макросклона Центрального Кавказа такова, что преобладают склоны северозападной и юго-восточной экспозиции. В связи с этим мы сочли возможным рассматривать отдельно теневые северные, северо-восточные, северо-западные склоны и солярные южные и юго-западные. Неожиданно выклинились отличия, наблюдающиеся в структуре населения люмбрицид восточных и юго-восточных склонов, которые мы также даем отдельно. Как правило, для Центрального Кавказа характерны более влажные пологие северные склоны и очень крутые южные с выраженными для этого типа рельефа эрозионно-денудационными процессами [10]. В пределах северных и северо-западных склонов согласно схеме Исаченко [11], учтены не только трансэллювиальные, аккумулятивно-трансэлювиальные (проточные водосборные понижения и эрозионные лощины на склонах), но и супераквальные местоположения. Тогда как на южных, юго-восточных и восточных – только транзитные.
Мы также считаем необходимым рассматривать отдельно биогеоценозы хребтов Джиналь-ского и Качкорташ, представляющие типичный случай эдафо-климатических условий пояса остепненных лугов эльбрусского варианта поясности и биогеоценозы ущелья р. Тызыл. Ориентация ущелий, образованных р. Золка Южная (Джинальский хр.) и р. Чегем (хр. Качкорташ) – северо-восточная. Р. Тызыл течет с запада на восток. Как известно, среднегодовые температуры воздуха выше в долинах северо-западного и северного направлений и ниже в долинах северовосточного направления [12].
Краткое описание района исследований . Орография ущелий пояса остепненных лугов характеризуется наличием платообразных возвышенностей, балок и русел рек со значительной крутизной склонов. Продолжительность периода вегетации растений 166-174 дня [13]. Безморозный период – 177-197 дней. Гидротермический коэффициент 1,5-2,0 – увлажнение умеренное. От лежащего ниже пояса луговых степей отличается широким развитием лугов с субальпийскими элементами, обусловленного лучшей обеспеченностью влагой и большей продолжительностью зимнего периода. Джинальский хребет представляет собой западную оконечность Мелового (Пастбищного) хребта. Значительную площадь занимают горные черноземы и горные луговочерноземные почвы. Почвообразующие породы – глинистые сланцы и известняки, залегающие пластами, наклонными на северо-восток. Преобладающий тип растительности – остепненные луга и луговые степи [1]. Рельеф Тызыльского ущелья гористый, почвы горно-луговые, местами заболоченные, растительность переходная от среднегорных лугов к субальпийским, местами значительная примесь степных элементов [14]. Северо-восточные склоны ущелья на исследованных участках заняты буковым лесом.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Общая характеристика фауны дождевых червей
В пределах рассматриваемой территории отмечены 12 видов дождевых червей: Aporrectodea caliginosa trapezoides (Dugès 1828) ,
A. jassyensis (Michaelsen, 1891), A. rosea (Savigny 1826) , Dendrobaena mariupolienis mariupolienis Wyssotzky 1898, D. octaedra (Savigny 1826), D. schmidti Michaelsen 1903, D. tellermanica Perel 1966, Dendrodrilus rubidus tenuis (Eisen 1874), Lumbricus rubellus (Hoffmeister 1843), Eisenia fetida (Savigny 1826) , Eiseniella tetraedra tetraedra (Savigny 1826), Octolasion lacteum (Örley 1885) (табл. 1, 2). Из них A. caliginosa trapezoides и A. jassyensis встречены менее чем в 1% проб. Наибольшим разнообразием отличается род Dendrobaena Eisen 1973, в пределах исследованной территории объединяющий четыре вида. Три вида включает род Aporrectodea Örley 1885, у остальных пяти родов – Dendrodrilus Omodeo 1956 , Lumbricus Örley 1885 , Eisenia Malm 1877 , Eiseniella Michaelsen 1900 и Octolasion Oerley 1885 – выявлено по одному виду. Число видов, отмеченных в поясе остепненных лугов, меньше, чем в поясах луговых степей и широколиственных лесов (13 и 16 видов), но превышает видовое богатство субальпийского пояса (9) [4]. Наиболее близка фауна люмбрицид рассматриваемого пояса к фаунам луговых степей, лесостепи и степной зоны, что в значительной степени определяется частой встречаемостью видов, хорошо приспособленных к перенесению как засушливого лета, так и холодной малоснежной зимы.
Как и в большинстве районов Северного Кавказа, собственно кавказская фауна представлена крымско-кавказскими субэндемиками D. mariupolienis mariupolienis, D. schmidti и близкородственным D. schmidti видом, еще недавно имевшим статус подвида - восточноевро-азиатским D. tellermanica. Средиземноморский A. jassyensis [15] отмечен в небольшом количестве проб. Преобладают виды с космополитным типом ареала (рис. 1 А).
В поясе остепненных лугов отмечены все жизненные формы люмбрицид (рис. 1 Б).
D. schmidti , у которого сильно варьируют размеры тела, окраска покровов и глубина стратификации в почвенном профиле, представлен собственно почвенной и почвенно-подстилочной морфо-экологическими формами.
Биотопическое распределение дождевых червей
Хребты Джинальский и Качкорташ (бас. р. Мокрая Золка и Чегем) . Наибольшее видовое богатство дождевых червей отмечено на плакорах. Ядро зональной фауны составляли 3 вида – D. tellermanica (доминант), собственно почвенная форма D. schmidti и норник D. mariupolienis mariupolienis (табл. 1) . Остальные виды отмечены в почвах фитоценозов с разной степенью пастбищной дигрессии – в злаково-разнотрав-но-клеверовом ( A. c. trapezoides , A. jassyensis и O. lacteum ) и пасторальном высокотравье ( De. rubidus tenuis и E. fetida ). На теневых склонах
К, 67%
Ср, 8%
Ве-Аз, 8%
Кр-Кв, 17%

|
ПМ, 8% |
D. schmidti |
|
A. c. trazoides, |
|
|
A. rosea, |
|
|
СП, 42% ■ |
A. jassyensis, |
|
D. tellermanica, |
|
|
O. lacteum |
|
|
ПП, 17% |
E. fetida, |
|
L. rubellus |
|
|
D. octaedra, |
|
|
П, 25% -X |
De. r. tenuis, |
|
Ei. t. tetraedra, |
|
|
D. m. mariupolienis |
|
Б
А
Рис. 1. Разнообразие хорологических (А) и морфо-экологических (Б) групп дождевых червей в поясе остепненных лугов эльбрусского варианта:
К – кавказские, Ср – средиземноморские, Ве-Аз – восточноевроазиатские, Кр-Кв – крымско-кавказские, ПМ – полиморфный, СП – собственно-почвенные, ПП – почвенно-подстилочные, П – подстилочные, Н – норник характер доминирования менялся. В зависимости от водного баланса почв среди люмбрицид преобладали собственно почвенная форма D. schmidti, или A. rosea и E. fetida. На высотах от 1000 м A. rosea концентрировался по берегам ручьев. Встречаемость D. tellermanica по сравнению с плакорами сокращалась, однако, в ряде биогеоценозов этот вид отмечен в качестве субдоминанта. Только на северных и северо-западных склонах зарегистрирован D. octaedra, полнее представлены D. mariupolienis mariupolienis и не характерный для степных и луговых сообществ De. rubidus tenuis. На солярных склонах отмечено наименьшее видовое богатство дождевых червей. На южных и юго-западных склонах преобладал D. schmidti (собственно почвенная форма), на наиболее крутых восточных и юго-восточных – D. tellermanica; реже встречался D. mariupolienis mariupolienis. В ложбинах, днищах речных балок и ущелий доминировал D. schmidti (собственно почвенная и почвенно-подстилочная формы), за исключением супераквальных местообитаний, в которых наибольшая численность отмечена у D. tellermanica и A. rosea.
В Тызыльском ущелье наименьшее видовое богатство с близкой численностью D. schmidti (собственно почвенная форма) и D. tellermanica отмечено на плакорах. На остальных позициях рельефа преобладали почвенно-подстилочная форма D. schmidti, часто встречался D. octaedra, реже D. m. mariupolienis. В пойменных и заболоченных почвах были обильны E. fetida и Ei. tetraedra. Остальные виды - O. lacteum, D. tellermanica, De. rubidus tenuis и L. rubellus – относились к категории редких. D. tellermanica зарегистрирован исключительно на плакорах и солярных склонах, O. lacteum – на теневых. К склоновому расположению тяго- теет и D. m. mariupolienis. Для большинства видов ландшафтной преференции не выявлено.
Хорологическая структура населения дождевых червей
Несмотря на то, что в поясе остепненных лугов видовое богатство дождевых червей с космополитным типом ареала было больше видового богатства всех остальных хорологических групп (рис. 1, 2), по численности хребтах Джи-нальский и Качкорташ чаще всего преобладали крымско-кавказские субэндемики и кавказский по происхождению восточноевро-азиатский D. tellermanica. Исключение составляли биогеоценозы, занимающие теневые склоны.
Разница в положении Тызыльского ущелья к направлению основных потоков переноса воздушных масс, формирующих климат пояса остепненных лугов, сказывалась в изменении хорологической структуры населения дождевых червей. Космополитные виды доминировали в биогеоценозах, расположенных на днище ущелья, неморально-степной восточноевро-азиатский D. tellermanica был обилен на плакорах и редко встречается на других позициях рельефа. Лесные космополиты D. octaedra и E. fetida зарегистрированы не только на теневых, но и на солярных склонах. В Тызыльском ущелье не отмечено видов со средиземноморским типом ареала.
Экологическая структура населения дождевых червей
В почвенных пробах хребтов Джинальского и Качкорташ по видовому богатству и численности преобладали собственно почвенные дождевые черви (табл. 1, рис.3). Почвенно-подстилочные виды,
Таб. 1. Видовой состав, биотопическое распределение, численность (экз./м2) дождевых червей хребтов Джинальского и Качкорташ
|
О о хо |
хо о |
04 |
40 |
1V0 |
40 |
g i У ° £ « 6 Й а 2 о 5 §-«^ Ь 6 Й о § о U g И Я Л а и Е CL> О t- о --=S ° S ia « га о о « ^Р « н о й га Я S О д я ю си 2 д S ^ S ° я 5 й a g а а ° S С W о 0 2 2 . W си VO CL> S _ 6 2 <2 ® 2 с« га 2 и га 2 о н U С 2 О гг< 2 ” $ 5 s Ь а о S Е-1 Щ rS S — g а § * § § и г: CD со s s ” о S U 2 о Я й 4 я у =§ а о л а ЕО и 2 0 0 2 Ю со S га си t[ со 2 О ^ CL> с га vd О CL> га g о > И га й н £ я - & к ° ° ^ и щ V О СО w У У X И S Ю « Я а а га га 2 о S S га га S 2 га 2 н о с 0 2 2 2 2 6 2 га га О щ О 2 pq 2 О й 2 ЕО ш р со Ю о 2 6 2 Ш « ! S Я о щ щ s а я Вот S о S а? я С > g |
|||||||||||
|
t* ^ аг о о U Л g со Н >3 СО а о 2 у 2 СО о о ч ^ от 43 14 О К к о 5 S У а7 S к си R си ч о 2 с и cti & о о и о 2 2 о о 2 ю со" CIS о и 52 о СО О ч S m см" хо сб Н |
0J о PQ О Ч 2 2 |
2 2 О О 2 ХО |
си о 2 си 2 |
04 |
ко |
04 |
04 |
40 |
04 |
||||||||
|
ХО О |
2 0J 2 2 2 |
н о 2 |
04 +1 |
ко |
40 ко +1 04 04 |
+1 40 |
+1 КО со 04 |
04 +1 04 04 |
О ко +1 СО |
КО +1 КО ко |
4? |
4? 04 КО |
|||||
|
J3 "53 g |
1 |
КО +1 КО |
|||||||||||||||
|
С) |
g |
3 3 CD |
1 |
04 +1 |
|||||||||||||
|
С) |
о с с g |
04 4-1 tn 00 |
4-1 |
||||||||||||||
|
о |
S 2 си С |
40 4-1 04 04 |
04 +1 04 ко |
||||||||||||||
|
щ |
g "S g S |
Th tn 4-1 tn 40 |
ш 4-1 Ш |
||||||||||||||
|
С) |
С "о .3 3 3 |
44* КО |
к S ч Щ |
||||||||||||||
|
Ьч |
Ҥ |
КО 04 +1 40 04 |
ОО |
О О<] |
I |
||||||||||||
|
С) |
g "S О |
1 |
ч Щ |
СО +1 04 ко |
в S к ч Щ |
+1 04 |
ть 4-1 04 |
КО +1 ОО |
Th 4-1 00 |
||||||||
|
С) |
В |
о 04 4-1 00 |
04 04 4-1 Th 04 |
о 4-1 40 04 |
S ч Щ |
4? 04 04 |
tn 4-1 |
о 4-1 00 40 |
ко +1 40 |
Th 4-1 40 |
|||||||
|
о ЯГ си о |
£5 |
X |
|||||||||||||||
|
си ч ГД |
си 2 2 0J К О 2 О 2 |
си 2 2 2 2 2 2 |
2 2 2 2 со 2 2 |
2 си 2 Л § |
в* В1 2 |
2 га 2 О 2 2 2 |
0J 2 2 Л 2 2 СО |
2 2 2 2 СО 2 |
си 2 л § |
2 2 В1 2 |
2 си g. 2 2 2 2 |
си 2 2 л 2 |
|||||
|
2 о 2 2 2 2 |
со О и |
2 О 5 |
СП 2 2" |
2 О 5 |
о к ч |
Я л Е^ си |
|||||||||||
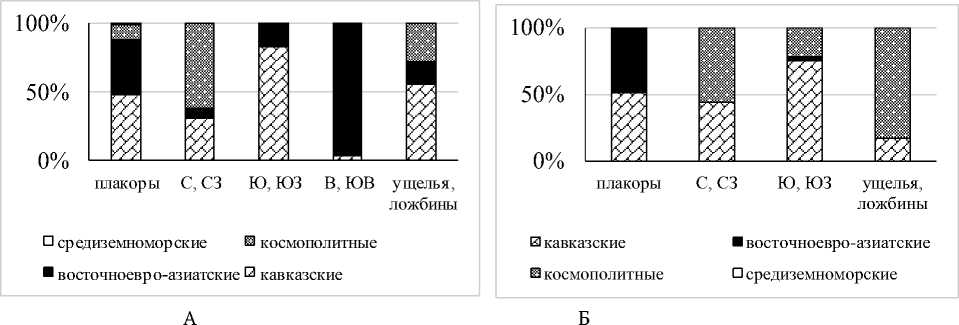
Рис. 2. Соотношение численности дождевых червей с разным типом ареала: А – хр. Джинальский (ущ. р. Золка Южная) и Качкорташ (ущ. р. Чегем), Б – ущ. р. Тызыл
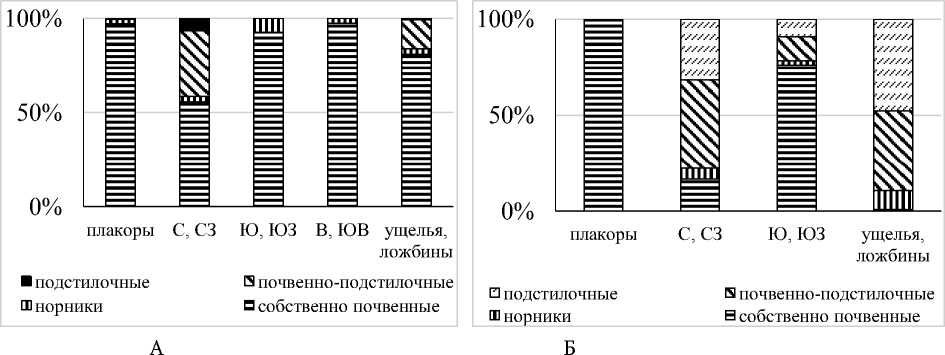
Рис. 3. Соотношение численности морфо-экологических групп дождевых червей: А – хр. Джинальский (ущ. р. Золка Южная) и Качкорташ (ущ. р. Чегем), %, Б – ущ. р. Тызыл
основной процент которых составляет E. fetida , занимали преимущественно теневые склоны.
В Тызыльском ущелье доминирование собственно почвенных видов отмечено только на плакорах и солярных склонах. На северных, северо-западных склонах и днище ущелья преобладали виды, питающиеся на поверхности почвы – подстилочные и почвенно-подстилочные.
Синэкологические индексы
Максимальное значение индекса Шеннона отмечено у дождевых червей хребтов Джиналь-ского и Качкорташ на теневых склонах, в Ты-зыльском ущелье – на днище. Минимум в обоих случаях наблюдался на солярных склонах. Вы-ровненность видовых обилий в обоих выборках имела верхний экстремум на плакорах, тогда как нижний в первой выборке наблюдался на восточных и юго-восточных склонах, во второй – на северо-западных. Доминирование в обоих выборках было сильнее выражено на солярных склонах, а его наименьшие значения у дождевых червей хребтов Джинальского и Качкорташ зарегистрированы на теневых склонах, а в Тызыль-ском ущелье – на плакорах.
Средняя общая численность дождевых червей
На Джинальском хребте и хребте Качкорташ наибольшая численность дождевых червей отмечена в ущельях и ложбинах, наименьшая – на солярных склонах (рис. 5). В Тызыльском ущелье наибольшее обилие наблюдалось на солярных склонах, наименьшее – на днище ущелья.
ЗАКЛЮЧЕНИЕ
В пределах пояса остепненных лугов эльбрусского варианта поясности отмечены 12 видов дождевых червей. Ядро зональной фауны составляли неморально-степные виды D. tellermanica (доминант), собственно почвенная форма D. schmidti и норник D. mariupolienis mariupolienis . D. tellermanica помимо плакоров доминировал на наиболее крутых восточных и юго-восточных склонах. На днищах ущелий и ложбин, теневых и части солярных (южных и юго-восточных) склонов по численности преобладали собственно почвенная, реже – почвенно-подстилочная формы D. schmidti. A. rosea на высотах от 1000 м встречался в пойменных почвах и по берегам ручьев.
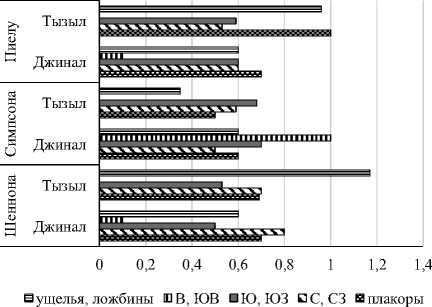
Рис. 4. Синэкологические индексы дождевых червей хребтов Джинальского+Качкорташ и Тызыльского ущелья
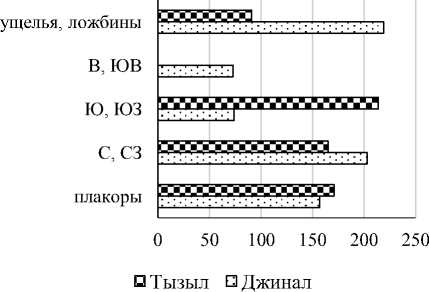
Рис. 5. Средняя общая численность дождевых червей хребтов Джинальского+Качкорташ и Тызыльского ущелья
Почвенно-подстилочные люмбрициды были агрегированы на теневых склонах и суперакваль-ных позициях на днище ушелья. Максимальное видовое богатство дождевых червей отмечено на плакорах, минимальное – на восточных и юго-восточных склонах. В остепненных лугах превалирующим фактором является характер распределения влаги, которая аккумулируется в отрицательных формах рельефа и на пологих теневых склонах, где отмечены максимальные показатели обилия дождевых червей. От поясов широколиственных лесов и луговых степей [4] пояс остепненных лугов отличается более «кавказским» типом населения – в большинстве сообществ преобладали крымско-кавказские субэндемики и вид кавказского происхождения D. tellermanica. Не смотря на то, в пределах пояса выявлены все морфо-экологические формы люмбрицид, доминировали собственно почвенные дождевые черви. Преобладание видов этой морфо-экологической группы обусловлено орографическими условиями эльбрусского варианта поясности. Сухие воздушные потоки Прикаспийской низменности, не задерживаемые передовыми хребтами, придают климату «степной» характер, вызывая общую ксерофи-тизацию ландшафтов эльбрусского варианта поясности [1]. На Центральном Кавказе, в долинах, расположенных выше 1000 м над ур. м., наблюдается явление температурной инверсии, при которой массы воздуха скапливаются в них, образуя «озера», температурный режим которых не всегда соответствует климатическим условиям местности [16]. Значительные отличия от «типичного варианта» ландшафтного распределения дождевых червей наблюдаются при изменении положения ущелий к направлению основных потоков переноса воздушных масс, формирующих климат пояса остепненных лугов. Зональный тип населения сохранялся на плакорах, где отмечено доминирование собственно почвенных неморально-степных видов. Преобладали виды этой группы и на солярных склонах, в отличие от хребтов Джинальского и Качкорташ, уже обогащенных лесными элементами фауны. Население теневых склонов и пойменных террас состояло в основном из лесных видов. Доминировали люмбрициды, питающиеся на поверхности почвы – подстилочные и почвенно-подстилочные. На днище ущелья отмечено преобладание видов с космополитным типом ареала и наименьшие показатели обилия, максимум которых смещен в отличие от типичного случая на солярные склоны. Согласно синэкологическим индексам, наиболее благоприятны были для дождевых червей в расположенном «поперек» ущелье днища ущелий, тогда как в типичном случае - теневые склоны и плакоры. Биоразнообразие, как функция влажности биогеоценоза, всегда убывало на солярных склонах. Однако, на хребтах Джинальском и Кач-корташ индекс Шеннона возрастал на теневых склонах, тогда как в Тызыле – на днище ущелья.
Искренне благодарю Ф.А. Темботову постоянное внимание, являющееся стимулом к работе. Т.С. Всеволодову-Перель – за бесценные для меня консультации, Н.Л. Цепкову – за определение гербарного материала и описание биотопов. А также В.И. Ланцова, А.А. Мокаеву, З.М. Юсупова, Ю.М. Саблирову, О.В. Умерову за участие во взятии почвенных проб.
Список литературы Фауна, население и особенности ландшафтного распределения дождевых червей (Oligochaeta, Lumbricidae) пояса остепненных лугов Центрального Кавказа
- Соколов, В.Е. Млекопитающие. Насекомоядные/В.Е. Соколов, А.К. Темботов. -М.: Наука, 1989. -С. 3-27.
- Рапопорт, И.Б. Видовой состав, синэкологические характеристики и ландшафтное распределение дождевых червей (Oligochaeta, Lumbricidae) субальпийского пояса Центрального Кавказа (терский вариант)/И.Б. Рапопорт//Изв. Самар. НЦ РАН. -2011. -Т. 13, № 1(5). -С. 1140-1245.
- Рапопорт, И.Б. Экология и хорология дождевых червей Кабардино-Балкарского государственного высокогорного заповедника (Центральный Кавказ)/И.Б. Рапопорт//Бюл. МОИП. -2012. -№2. -С. 65-71.
- Рапопорт, И.Б. Фауна, экология и высотно-поясное распределение дождевых червей центральной части Северного Кавказа: автореф. дис. канд. биол. наук. 22.02.2011/Рапопорт Ирина Борисовна. -Тольятти, 2011. -306 с.
- Рапопорт, И.Б. Высотное распределение дождевых червей (Oligochaeta, Lumbricidae) в центральной части Северного Кавказа/И.Б. Рапопорт//Зоол. журн. -2013. -№ 1. -С. 3-10.
- Гиляров, М.С. Учет крупных беспозвоночных (мезофауны)/М.С. Гиляров//Методы почвенно-зоологических исследований. -М.: Наука, 1975. -С. 12-29.
- Перель, Т.С. Распространение и закономерности распределения дождевых червей фауны СССР/Т. С. Перель. -М.: Наука, 1979. -275 с.
- Всеволодова-Перель, Т.С. Дождевые черви фауны России. -М.: Наука, 1997. 102 с.
- Нагуманова, Н.Г. Почвенные беспозвоночные в гидрологическом ряду Бузулукского бора/Н.Г. Нагуманова//Вестник ОГУ. -2006. -№5. -С. 96-103.
- Чупахин, В.М. Физическая география Северного Кавказа/В.М. Чупахин. -Ростов-на-Дону: Рост. ун-т, 1974. -200 с.
- Исаченко, И.Г. Ландшафтоведение и физико-географическое районирование/И.Г. Исаченко. -М.: Высшая школа, 1991. -366 с.
- Солошенко, М.Н. Распределение основных ландшафтообразующих климатических характеристик в соответствии с условиями рельефа на территории Северного Кавказа//Тр. ВГИ. -М.: Гидрометеоиздат, 1988. -Вып. 71, № 9. -С. 49-62.
- Почвы Кабардино-Балкарской АССР и рекомендации по их использованию. -Нальчик: СевКавНИИГИПРоЗем, 1984. -201 с.
- Ёжкина, А.Н. Растительность северного склона Скалистого хребта между р.р. Баксаном и Тызылом/А.Н. Ёжкина//Ученые записки. Сер. биол. -Вып. 16. -Нальчик, 1962. -С. 57-64.
- Omodeo, P. Particolaritá della Zoogeografia dei Lombrichi/Р. Omodeo//Boll. Zool. -1952. -№ 9. -P. 349-359.
- Разумов, В.В. Экосистемы гор Центрального Кавказа и здоровье человека/В.В. Разумов, Х.А. Курданов, Л.А. Разумова, А.Г. Крохмаль, Л.М. Батырбекова. -М.-Ставрополь, 2003. -447 с.
