Фауна полихет в северо-восточной части Баренцева моря

Автор: Дикаева Д. Р.
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Биологические науки. Науки о Земле
Статья в выпуске: 2 т.25, 2022 года.
Бесплатный доступ
Северо-восток Баренцева моря (район между архипелагами Земля Франца-Иосифа и Новая Земля) является одним из малоизученных районов, отличающихся сложностью структуры гидрологического режима, рельефа дна и структуры донных осадков. Данный район характеризуется преобладанием коричневых грунтов с большим количеством раковин фораминифер, что оказывает влияние на видовой состав и структуру бентосных сообществ. На основе материала, собранного в северо-восточной части Баренцева моря в декабре 2019 г. в ходе экспедиций сотрудников Мурманского морского биологического института на научно-исследовательском судне "Дальние Зеленцы", проанализирован видовой состав и количественные характеристики полихет. В районе исследования в зависимости от условий среды (типа донных осадков, глубины, гидрологических условий) выявлены видовые комплексы полихет. В районах краевых желобов по биомассе и численности доминирует детритофаг Spiochaetopterus typicus, образующий обильные поселения на мягких илисто-песчаных, илисто-глинистых грунтах. Низкие значения биомассы и численности отмечены на коричневых илах с большим количеством фораминифер, где доминирует Laonice cirrata. На склонах подводных возвышенностей, омываемых придонными течениями, на грубых песчаных грунтах с низким содержанием питательных веществ выявлено доминирование Nephtys ciliata. В районе исследования был отмечен также вид Sphaerodoridium kolchaki sp. n. Полученные данные дополняют сведения о видовом составе северо-восточной части Баренцева моря и позволяют проследить за дальнейшими изменениями в составе и структуре донных сообществ.
Полихеты, Баренцево море, биомасса, плотность поселения, Polychaeta, Barents Sea, biomass, abundanсe
Короткий адрес: https://sciup.org/142231970
IDR: 142231970 | УДК: 595.14.142.2(268.45) | DOI: 10.21443/1560-9278-2022-25-2-79-90
Текст статьи Фауна полихет в северо-восточной части Баренцева моря
Мурманский морской биологический институт РАН, г. Мурманск, Россия; e-mail: , ORCID:
Дикаева Д. Р. Фауна полихет северо-восточной части Баренцева моря. Вестник МГТУ. 2022. Т. 25, № 2. С. 79–90. DOI:
Dikaeva, D. R. 2022. Polychaeta fauna in the northeastern Barents Sea. Vestnik of MSTU, 25(2), pp. 79–90. (In Russ.) DOI:
Северо-восток Баренцева моря отличается сложностью структуры гидрологического режима, рельефа дна и структуры донных осадков. Зообентос в районе архипелага Новая Земля изучен достаточно подробно ( Броцкая и др. 1939; Денисенко и др., 1995; Kiyko et al., 1997; Фролова, 2000; Любина и др., 2008; Артюх и др., 2008; Дикаева и др., 2011; Захаров и др., 2018 ). Однако о фаунистическом составе и количественном распределении таксоцена полихет в северо-восточной части Баренцева моря (районе между архипелагами Земля Франца-Иосифа и Новая Земля) в литературе содержится мало сведений. Настоящая работа является продолжением исследования распределения полихет в районах краевых желобов (Северо-Восточного желоба и желоба Святой Анны) ( Фролова и др., 2017 ). Распределение и жизнедеятельность морских организмов предопределяется геоморфологией дна: глубиной, рельефом, донными отложениями. Станции, расположенные в районе исследования, пересекают Северо-Восточный желоб и желоб Святой Анны. Желоба являются районами аккумуляции осадочного вещества, что увеличивает кормовую базу организмов, обитающих в грунте, в результате чего возрастает биомасса и плотность поселения. В северо-восточном районе Баренцева моря преобладает холодная арктическая водная масса; по краевым желобам поступают трансформированные атлантические воды, способствующие увеличению видового разнообразия донных организмов. Данный район характеризуется преобладанием коричневых грунтов с большим количеством раковин фораминифер, что приводит к обеднению видового состава и количественных характеристик бентосных сообществ ( Броцкая и др., 1939; Зенкевич, 1947 ).
В настоящее время отмечено увеличение численности краба-стригуна опилио Chionoecetes opilio на северо-востоке Баренцева моря и у юго-западной оконечности архипелага Новая Земля, что оказывает значительное влияние на донные биоценозы ( Захаров и др., 2018 ).
Целью исследования является определение видового состава, количественных характеристик и особенностей распределения полихет в северо-восточной части Баренцева моря в зависимости от структуры рельефа дна и гидрологического режима.
Материалы и методы
Материалом исследования послужили 35 количественных проб, собранных на 12 станциях сотрудниками Мурманского морского биологического института КНЦ РАН в ходе экспедиций на научно-исследовательском судне "Дальние Зеленцы" в декабре 2019 г. (рис. 1). Пробы зообентоса отбирали с помощью дночерпателя Ван-Вина (KC Denmark, Дания) (площадь захвата 0,1 м2) в 3-кратной повторности. Собранный грунт промывали через капроновое сито с ячеей размером 0,75 мм, фиксировали 4%-м формалином с последующим переводом беспозвоночных в 70%-й спирт.
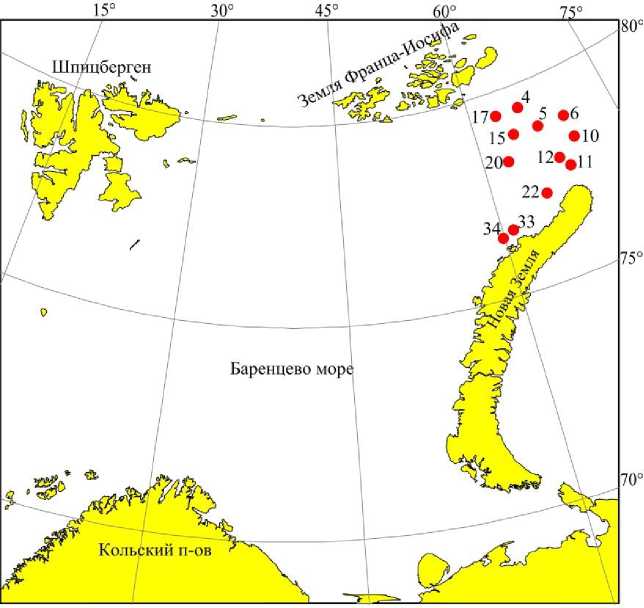
Рис. 1. Карта-схема расположения бентосных станций в 2019 г. Fig. 1. Map-scheme of benthic stations in 2019
В пробе подсчитывали количество особей каждого вида и определяли их сырую массу на торсионных весах с точностью до 0,001 г. Перед взвешиванием полихеты обсушивали на фильтровальной бумаге. Для каждой станции рассчитывали средние значения биомассы и численности организмов на квадратный метр площади дна с определением ошибки средней. Все полихеты взвешивали без трубок, кроме вида Spiochaetopterus typicus и полихет семейства Serpulidae, строящих трубки из вещества, выделяемого самими червями. Обрывки полихет в пробе взвешивали по "головным" частям. Выделение станций по сходству видового состава выполняли с помощью кластерного анализа на основе коэффициента сходства Брея – Кертиса ( Bray et al., 1957 ). В качестве меры обилия для выделения доминантной группы видов использовали показатель относительной интенсивности метаболизма
M = KN0,25B0,75, где N – численность организмов (экз./м2); B – биомасса (г/м2); K – таксоноспецифический коэффициент удельной интенсивности метаболизма (кДж/ч) (Алимов, 1979; Денисенко и др., 2006).
Таксономия полихет приведена в соответствии с World Register of Marine Species1.
Результаты и обсуждение
В районе исследования отмечено 82 таксона многощетинковых червей, из них 65 определены до вида. Видовое богатство варьировало от 13 до 40 видов на каждой станции. Высокое видовое разнообразие отмечено на глубине 106 м на песчаных, глинистых грунтах с камнями. Низкие значения видового разнообразия отмечены на глубине 392 м на песчанистых коричневых илах с глиной и фораминиферами (табл. 1). Биомасса полихет в районе исследования варьирует от 0,8 до 46 г/м2, плотность поселения – от 183 до 2 550 экз./м2. Максимальные значения биомассы и плотности поселения отмечены в районе архипелага Новая Земля (плато Литке) на среднезернистых песках с вязкой глиной, минимальные значения – в районе Северо-Восточного желоба на песчаных коричневых грунтах с серой глиной и фораминиферами. В биогеографическом составе преобладают бореально-арктические виды.
Таблица 1. Количественные характеристики полихет северо-восточной части Баренцева моря Table 1. Quantitative parameters of polychaete communities in the northeastern Barents Sea
|
Номер станции |
Глубина, м |
Количество видов |
Биомасса, г/м2 |
Численность, экз./м2 |
Доминирующий вид (по метаболизму) |
|
4 |
427 |
21 |
4 ± 2 |
327 ± 90 |
Spiochaetopterus typicus |
|
5 |
343 |
37 |
19 ± 4 |
563 ± 150 |
Spiochaetopterus typicus |
|
6 |
507 |
28 |
14 ± 5 |
573 ± 43 |
Spiochaetopterus typicus |
|
10 |
445 |
29 |
13 ± 7 |
710 ± 172 |
Laonice cirrata |
|
11 |
497 |
30 |
32 ± 6 |
1 470 ± 199 |
Spiochaetopterus typicus |
|
12 |
430 |
24 |
3 ± 1 |
620 ± 191 |
Lumbrineridae g. sp., Spiochaetopterus typicus |
|
15 |
361 |
27 |
15 ± 2 |
600 ± 82 |
Spiochaetopterus typicus |
|
17 |
186 |
28 |
4 ± 1 |
427 ± 82 |
Aglaophamus malmgreni |
|
20 |
392 |
13 |
0,8 ± 0,2 |
183 ± 81 |
Lumbrineridae g. sp. |
|
22 |
106 |
40 |
46 ± 19 |
2 550 ± 234 |
Spiochaetopterus typicus |
|
33 |
105 |
21 |
11 ± 1 |
1 235 ± 355 |
Lumbrineridae g. sp. |
|
34 |
127 |
24 |
19 ± 12 |
1 247 ± 488 |
Nephtys ciliatа |
Сравнение видовых списков полихет методом кластерного анализа позволило выделить в районе исследования три видовых комплекса, отличающихся друг от друга по видовому составу и количественным характеристикам (рис. 2).
Первый комплекс (станции 11, 22, 15, 5, 6) расположен на глубинах от 106 до 497 м на коричневых илисто-песчаных, глинистых грунтах с примесью фораминифер. В этом комплексе отмечено 72 вида полихет. По относительной интенсивности метаболизма и биомассе преобладает собирающий детритофаг Spiochaetopterus typicus . Этот комплекс включает две группы станций, различающихся по количественным характеристикам.
Первая группа станций (11, 22) расположена вблизи северной оконечности архипелага Новая Земля на глубинах от 106 до 497 м на илисто-песчаных, глинистых грунтах. В данном районе отмечены высокие значения биомасса (39 ± 13 г/м2) и плотности поселения полихет (2 010 ± 216 экз./м2).
Вторая группа станций (15, 5, 6) расположена в районе Северо-Восточного желоба и желоба Святой Анны на глубинах от 343 до 507 м, где преобладают коричневые илисто-глинистые грунты с примесью песка и фораминифер. В данном районе отмечено снижение биомассы (16 ± 4 г/м2) и плотности поселения полихет (580 ± 90 экз./м2).
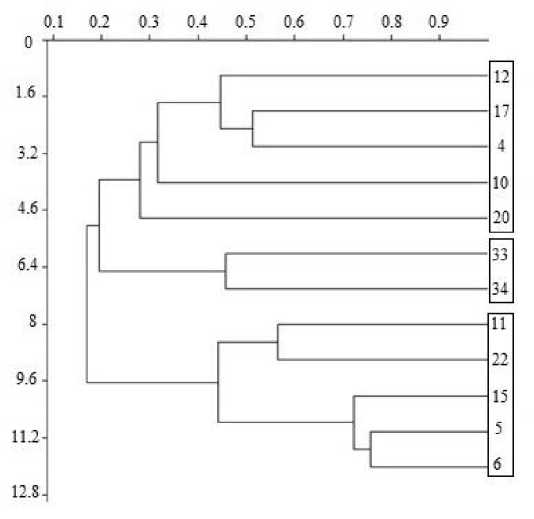
Рис. 2. Дендрограмма сходства видового состава полихет в северо-восточной части Баренцева моря в 2019 г.
Fig. 1. Dendrogram of the polychaete species similarity in the northeastern Barents Sea in 2019
Второй комплекс (станции 33, 34) расположен вдоль западного побережья северного острова Новой Земли и отличается небольшими глубинами (105, 127 м), где преобладают крупнозернистые песчаные грунты с примесью камней и серой вязкой глины. В данном комплексе отмечено 33 вида полихет. По уровню метаболизма преобладает Nephtys ciliata, по биомассе – Scoletoma fragilis, Nephtys ciliata. Комплекс отличается невысокими значениями биомассы (15 ± 7 г/м2) и средней плотностью поселения (1 240 ± 421 экз./м2).
Третий комплекс (станции 12, 17, 4, 10, 20) находится в районе Северо-Восточного желоба и желоба Святой Анны на глубинах от 186 до 445 м, где преобладают коричневые илисто-песчаные, глинистые грунты с большим количеством фораминифер. Здесь отмечено 54 вида полихет. По уровню метаболизма доминируют Laonice cirrata. Комплекс характеризуется минимальными значениями биомассы (5 ± 2 г/м2) и плотности поселения (453 ± 123 экз./м2).
В Баренцевом море был отмечен вид Sphaerodoridium kolchaki sp. n.: в 2003 и 2004 гг. – в районе 72° с. ш., 38° в. д. и 76°30´ с. ш., 47° в. д. ( Gagaev, 2015 ); в 2017 г. – в районе архипелага Шпицберген на станциях с координатами 77° с. ш., 25° в. д. и 77° с. ш., 19° в. д. на глубинах 79–151 м на илистых, глинистых грунтах с камнями и ракушей ( Дикаева и др., 2018 ). В ходе настоящего исследования Sphaerodoridium kolchaki sp. n. был встречен на двух станциях в районе желоба Святой Анны на коричневых илисто-глинистых грунтах с небольшим количеством камней на глубинах 445–497 м.
В районе исследования в зависимости от состава грунта и глубины определено изменение видового состава и количественных характеристик полихет. В районах краевых желобов максимальные значения биомассы и плотности полихет отмечены на илисто-песчаных, илисто-глинистых грунтах в области обильного развития детритофага Spiochaetopterus typicus. В глубоководных районах на грунтах с преобладанием коричневого ила с большим количеством фораминифер отмечен видовой комплекс с доминированием спиониды Laonice cirrata , отличающийся низкими значениями биомассы и плотности поселения полихет. Значительное снижение биомассы бентоса в глубоководных районах на мягких коричневых илах отмечено в Карском море ( Филатова и др., 1957 ). Глубоководные районы с преобладанием коричневых грунтов характеризуются неблагоприятным газовым режимом придонного слоя и низким содержанием органического вещества, что приводит к снижению количественных характеристик полихет.
С уменьшением глубины на склонах подводных возвышенностей северного острова Новой Земли отмечено доминирование плотоядной полихеты Nephtys ciliata . Здесь на жестких грунтах с преобладанием грубообломочного материала в зоне активной гидродинамики в результате взаимодействия теплых и холодных течений, где процессы переноса осадков доминируют над процессами их образования, созданы неблагоприятные условия для развития детритофагов, что приводит к смене доминирующих видов и снижению биомассы полихет. Появление нового, недавно описанного вида Sphaerodoridium kolchaki sp. n . ( Gagaev, 2015 ) связано с недостаточной изученностью фауны в этом районе.
Таблица 2. Видовой состав и средняя биомасса полихет (г/м2) в северо-восточной части Баренцева моря в 2019 г. Table 2. Species composition and biomass of polychaetеs (g/m2) in the northeastern Barents Sea in 2019
|
Таксон |
Станция |
|||||||||||
|
4 |
5 |
6 |
10 |
11 |
12 |
15 |
17 |
20 |
22 |
33 |
34 |
|
|
Сем. Phyllodocidae |
||||||||||||
|
Eteone flava (Fabricius, 1780) |
– |
0,010 |
– |
0,027 |
0,133 |
0,007 |
– |
– |
– |
0,187 |
0,030 |
0,047 |
|
Phyllodoce groenlandica Örsted, 1842 |
0,007 |
– |
0,007 |
– |
0,350 |
– |
0,080 |
– |
– |
0,013 |
0,001 |
0,003 |
|
Phyllodocidae g. sp. |
– |
– |
– |
0,003 |
– |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Polynoidae |
||||||||||||
|
Harmothoe impar impar (Johnston, 1839) sensu Malmgren, 1865 |
– |
0,080 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Enipo torelli (Malmgren, 1865) |
– |
– |
– |
– |
0,127 |
– |
– |
– |
– |
0,013 |
0,090 |
0,210 |
|
Polynoidae g. sp. |
– |
0,033 |
0,007 |
– |
– |
– |
– |
– |
– |
0,013 |
0,003 |
0,017 |
|
Gattyana cirrhosa (Pallas, 1766) |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,173 |
– |
0,193 |
|
Сем. Sigalionidae |
||||||||||||
|
Pholoe longa (O. F. Müller, 1776) |
– |
0,040 |
0,010 |
– |
0,017 |
0,020 |
0,013 |
0,002 |
– |
0,567 |
0,110 |
0,073 |
|
Pholoe sp. |
– |
– |
– |
0,093 |
– |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Nephtyidae |
||||||||||||
|
Aglaophamus malmgreni (Théel, 1879) |
0,967 |
– |
0,013 |
0,053 |
– |
0,133 |
0,487 |
0,960 |
0,067 |
– |
– |
– |
|
Nephtys ciliatа (Müller, 1788) |
– |
0,067 |
– |
0,140 |
0,217 |
– |
– |
– |
– |
– |
1,810 |
5,593 |
|
Nephtys sp. |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,053 |
|
Nephtyidae g.sp. |
– |
0,013 |
– |
0,007 |
– |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Glyceridae |
||||||||||||
|
Glycera lapidum Qautrefages, 1866 |
– |
0,160 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Nereididae |
||||||||||||
|
Nereis zonata Malmgren, 1867 |
– |
0,447 |
– |
– |
0,160 |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Hesionidae |
||||||||||||
|
Hesionidae g. sp. |
– |
– |
0,003 |
– |
– |
– |
– |
– |
– |
0,107 |
– |
0,007 |
|
Сем. Syllidae |
||||||||||||
|
Syllidae g. sp. |
– |
– |
– |
– |
– |
– |
0,003 |
0,030 |
– |
0,007 |
– |
0,037 |
|
Сем. Sphaerodoridae |
||||||||||||
|
Sphaerodoropsis philippi (Fauvel, 1911) |
– |
– |
0,023 |
– |
– |
0,003 |
– |
0,020 |
– |
0,002 |
– |
|
|
Sphaerodoridum kolchaki sp. n. |
– |
– |
– |
0,003 |
0,007 |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Onuphidae |
||||||||||||
|
Nothria hyperborea (Hansen, 1878) |
– |
0,067 |
– |
0,003 |
– |
– |
– |
0,207 |
– |
1,067 |
– |
– |
|
Сем. Lumbrineridae |
||||||||||||
|
Scoletoma fragilis ( O. F. Müller, 1776) |
– |
– |
– |
0,002 |
0,060 |
0,005 |
– |
– |
– |
0,010 |
0,160 |
0,038 |
|
Lumbrineridae g. sp. |
0,487 |
0,103 |
0,037 |
0,407 |
1,093 |
0,447 |
0,353 |
0,393 |
0,180 |
1,593 |
1,590 |
0,693 |
|
Сем. Dorvilleidae |
||||||||||||
|
Dorvilleidae g. sp. |
0,033 |
0,033 |
0,003 |
0,003 |
– |
– |
0,033 |
0,017 |
– |
0,003 |
– |
– |
|
Сем. Spionidae |
||||||||||||
|
Laonice cirrata (M. Sars, 1851) |
– |
0,067 |
0,457 |
7,393 |
3,927 |
– |
0,057 |
0,140 |
– |
– |
– |
– |
|
Prionospio cirrifera Wirén, 1883 |
0,043 |
0,043 |
0,093 |
0,057 |
0,360 |
0,170 |
0,117 |
0,030 |
0,020 |
– |
– |
– |
|
Dipolydora coeca (Örsted, 1843) |
– |
– |
– |
0,017 |
– |
– |
– |
– |
– |
– |
– |
– |
|
Dipolydora caulleryi (Mesnil, 1897) |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,003 |
|
Spio limicola Verrill, 1879 |
– |
– |
– |
0,003 |
0,033 |
– |
– |
– |
– |
0,253 |
0,050 |
0,093 |
|
Spio armata (Thulin, 1957) |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,013 |
– |
– |
|
Spiophanes kroyeri Grube, 1860 |
0,120 |
0,017 |
0,113 |
– |
0,003 |
– |
0,300 |
0,010 |
– |
– |
– |
– |
|
Сем. Apistobranchidae |
||||||||||||
|
Apistobranchus tullbergi (Théel, 1879) |
– |
0,027 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,013 |
|
Сем. Trochochaetidae |
||||||||||||
|
Тrochochaeta carica (Birula, 1897) |
– |
– |
0,020 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Chaetopteridae |
||||||||||||
|
Spiochaetopterus typicus M Sars, 1856 |
1,833 |
13,080 |
12,460 |
1,033 |
14,670 |
0,653 |
12,770 |
0,207 |
– |
19,027 |
0,710 |
0,773 |
|
Сем. Cossuridae |
||||||||||||
|
Cossura longocirrata Webster & Benedict, 1887 |
– |
– |
0,002 |
0,005 |
0,010 |
0,005 |
– |
– |
0,002 |
0,015 |
0,010 |
0,008 |
|
Сем. Cirratulidae |
||||||||||||
|
Cirratulidae g.sp. |
0,067 |
– |
0,047 |
0,233 |
0,473 |
0,240 |
0,037 |
0,080 |
0,100 |
0,707 |
0,260 |
0,170 |
|
Сем. Flabelligeridae |
||||||||||||
|
Brada inhabilis (Rathke, 1843) |
– |
– |
– |
– |
– |
– |
– |
– |
– |
2,020 |
– |
– |
|
Сем. Scalibregmatidae |
||||||||||||
|
Scalibregma inflatum Rathke, 1843 |
– |
0,010 |
– |
0,007 |
0,047 |
0,033 |
0,000 |
0,013 |
0,017 |
0,133 |
– |
0,160 |
|
Pseudoscalibregma parvum (Hansen, 1879) |
0,100 |
– |
0,487 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Сем. Opheliidae |
||||||||||||
|
Ophelina abranchiata Støp-Bowitz, 1948 |
0,010 |
– |
– |
– |
0,013 |
0,033 |
– |
0,070 |
– |
– |
– |
– |
|
Ophelina acuminata Örsted, 1843 |
– |
0,010 |
– |
– |
– |
– |
– |
– |
– |
0,013 |
0,140 |
– |
|
Ophelina cylindricaudata (Hansen, 1879) |
0,003 |
0,013 |
0,008 |
– |
0,012 |
0,000 |
0,022 |
0,023 |
0,040 |
0,002 |
– |
– |
|
Сем. Orbiniidae |
||||||||||||
|
Leitoscoloplos acutus (Verrill, 1873) |
– |
– |
– |
0,007 0,433 0,460 |
– |
– |
0,010 |
0,253 |
0,150 |
0,120 |
||
|
Сем. Paraonidae |
||||||||||||
|
Aricidea (Strelzovia) hartmanae (Strelzov, 1968) |
0,003 |
– |
– |
0,020 |
– |
0,003 |
– |
– |
0,007 |
0,023 |
0,005 |
– |
|
Aricidea ( Strelzovia ) quadrilobata Webster & Benedict, 1887 |
0,008 |
– |
– |
– |
– |
0,007 |
0,013 |
– |
– |
– |
– |
0,007 |
|
Aricidea nolani Webster & Benedict, 1887 |
– |
– |
0,002 |
– |
– |
– |
– |
– |
– |
– |
0,055 |
0,050 |
|
Aricidea sp. |
– |
– |
0,003 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Levinsenia gracilis (Tauber, 1879) |
– |
– |
– |
0,002 |
0,060 |
0,005 |
– |
– |
– |
0,010 |
0,160 |
0,038 |
|
Сем. Maldanidae |
||||||||||||
|
Maldane sarsi Malmgren, 1865 |
0,167 |
0,000 |
0,007 |
2,360 |
3,967 |
– |
– |
– |
– |
2,933 |
– |
0,417 |
|
Maldanidae g. sp |
– |
0,017 |
0,020 |
– |
– |
– |
– |
– |
– |
– |
5,040 |
– |
|
Nicomache lumbricalis (Fabricius, 1780) |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,527 |
– |
– |
|
Notoproctus oculatus Arwidsson, 1906 |
– |
0,015 |
0,123 |
– |
– |
0,013 |
– |
0,030 |
– |
– |
– |
– |
|
Praxillella gracilis (M. Sars, 1861) |
– |
– |
0,122 |
0,133 |
5,070 |
0,087 |
– |
0,567 |
– |
– |
– |
0,147 |
|
Praxillella praetermissa (Malmgren, 1865) |
– |
0,017 |
– |
– |
– |
– |
– |
0,040 |
0,007 |
0,340 |
– |
– |
|
Praxillura longissima Arwidsson, 1906 |
– |
0,150 |
– |
– |
0,020 |
– |
0,013 |
0,340 |
– |
– |
– |
– |
|
Сем. Capitellidae |
||||||||||||
|
Heteromastus filiformis (Claparède, 1864) |
0,013 |
0,127 |
– |
0,300 |
0,213 |
0,123 |
– |
0,053 |
0,023 |
0,030 |
0,080 |
0,007 |
|
Notomastus latericeus Sars, 1851 |
– |
0,007 |
0,023 |
– |
– |
– |
0,400 |
– |
– |
0,760 |
– |
– |
|
Сем. Oweniidae |
||||||||||||
|
Galathowenia oculata (Zachs, 1923) |
– |
0,003 |
0,003 |
0,010 |
0,097 |
0,020 |
– |
– |
– |
0,013 |
– |
– |
|
Myriochele heeri Malmgren, 1867 |
– |
0,003 |
0,033 |
– |
0,003 |
0,007 |
0,003 |
0,040 |
– |
0,003 |
– |
– |
|
Owenia fusiformis Delle Chiaje, 1844 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,367 |
– |
– |
|
Oweniidae g. sp. |
– |
0,010 |
0,010 |
– |
– |
– |
0,003 |
0,003 |
– |
– |
– |
– |
|
Сем. Pectinariidae |
||||||||||||
|
Cistenides hyperborea Malmgren, 1866 |
– |
– |
– |
0,477 0,053 0,213 |
– |
– |
0,233 |
5,100 |
0,660 |
0,067 |
||
|
Сем. Ampharetidae |
||||||||||||
|
Ampharete finmarchica (M. Sars, 1865) |
0,007 |
– |
– |
0,133 |
– |
– |
– |
– |
– |
2,280 |
0,005 |
– |
|
Ampharetidae g. sp. |
– |
0,005 |
– |
– |
– |
– |
0,010 |
0,002 |
– |
0,003 |
– |
0,003 |
|
Sosane wireni (Hessle, 1917) |
0,003 |
0,003 |
– |
– |
– |
– |
0,007 |
– |
– |
– |
– |
– |
|
Glyphanostomum pallescens (Théel, 1879) |
0,003 |
0,010 |
– |
0,013 |
– |
– |
0,013 |
– |
– |
– |
– |
– |
|
Lysippe labiata Malmgren, 1866 |
– |
– |
– |
0,013 |
0,010 |
– |
– |
– |
– |
0,087 |
– |
0,027 |
|
Сем. Melinnidae |
||||||||||||
|
Melinna elisabethae McIntosh, 1914 |
– |
– |
– |
– |
– |
– |
– |
0,107 |
– |
– |
– |
– |
|
Melinnopsis arctica (Annenkova, 1931) |
– |
– |
– |
– |
– |
– |
0,047 |
– |
– |
– |
– |
– |
|
Сем. Terebellidae |
||||||||||||
|
Amphitrite cirrata Müller, 1776 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,173 |
– |
– |
|
Artacama proboscidea Malmgren, 1866 |
– |
– |
– |
– |
– |
– |
– |
0,260 |
– |
– |
– |
– |
|
Laphania boecki Malmgren, 1866 |
– |
– |
– |
– |
0,013 |
– |
– |
– |
– |
0,140 |
– |
– |
|
Polycirrus medusa Grube, 1850 |
– |
– |
– |
– |
– |
0,240 |
– |
– |
– |
– |
– |
– |
|
Thelepus cincinnatus (Fabricius, 1780) |
– |
4,040 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
Terebellidae g. sp. |
– |
0,027 |
– |
– |
– |
– |
0,007 |
– |
– |
– |
0,015 |
0,120 |
|
Сем. Trichobranchidae |
||||||||||||
|
Тerebellides stroemii Sars, 1835 |
0,003 |
0,020 |
0,007 |
– |
0,040 |
– |
0,007 |
0,013 |
– |
– |
0,185 |
1,492 |
|
Terebellides gracilis Malm, 1874 |
– |
– |
– |
– |
– |
– |
0,013 |
– |
– |
– |
– |
– |
|
Сем. Sabellidae |
||||||||||||
|
Chone sp. |
0,007 |
0,017 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,013 |
|
Chone infundibuliformis Krøyer, 1856 |
– |
0,023 |
– |
– |
– |
– |
0,013 |
– |
– |
– |
– |
– |
|
Chone murmanica Lukasch, 1910 |
– |
0,020 |
– |
– |
– |
0,003 |
0,010 |
0,002 |
– |
– |
– |
– |
|
Euchone analis (Kröyer, 1856) |
– |
– |
– |
– |
– |
0,007 |
– |
– |
0,060 |
0,000 |
– |
– |
|
Сем. Serpulidae |
||||||||||||
|
Bushiella (Jugaria) similis (Bush, 1905) |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,007 |
|
Spirorbinae g.sp. |
– |
– |
– |
– |
– |
– |
– |
– |
– |
0,007 |
– |
– |
По данным Броцкой В. А. и Зенкевича Л. А. (1939), в районе между архипелагами Земля Франца-Иосифа и Новая Земля отмечены низкие значения биомассы полихет (0,5–8 г/м2, 12–16 % от общей биомассы). По данным 1995 г., в желобе Святой Анны на глубинах 135–620 м на коричневых илах доминировал детритофаг Spiochaetopterus typicus , а в западной части желоба по биомассе преобладала спионида Laonice cirrata ( Фролова, 2009 ). В предыдущих исследованиях нами было изучено распределение таксоцена полихет в районах краевых желобов (Медвеженский желоб, желоб Франц-Виктория и желоб Святой Анны) ( Фролова и др., 2017 ). В данных районах биомасса полихет варьировала от 126 до 5 г/м2, плотность поселения – от 5 460 до 260 экз./м2; в желобе Святой Анны биомасса составляла 15 г/м2, плотность поселения – 420 экз./м2. В районах краевых желобов по биомассе и численности доминировал детритофаг Spiochaetopterus typicus , образующий обильные поселения в районе Медвеженского желоба, подверженного сильному влиянию атлантических водных масс. По данным 2003–2006 гг., сообщество с доминированием Spiochaetopterus typicus было отмечено на глубине 200–250 м на мягких илистых, илисто-песчаных грунтах в районе Новоземельской возвышенности, а также на склонах Новоземельского желоба и полуострова Адмиралтейства ( Захаров и др., 2018 ).
В результате сравнения современных данных с данными предыдущих исследований существенных изменений в составе доминирующих видов не выявлено. Во все периоды исследований в северо-восточной части Баренцева моря в глубоководных районах на илисто-песчаных, илисто-глинистых грунтах по биомассе доминировал детритофаг Spiochaetopterus typicus ; на коричневых илах в районе желоба Святой Анны отмечалось преобладание спиониды Laonice cirrata .
На близлежащих станциях, расположенных в районе Новоземельского мелководья, определено ( Дикаева и др., 2011 ) снижение биомассы полихет и уменьшение доли собирающих детритофагов в 2019 г. (станции 33, 34) по сравнению с этими показателями, полученными в 2006 г. (станции 25, 26) (рис. 3, 4). Снижение общей биомассы полихет происходило за счет снижения биомассы собирающего детритофага Spiochaetopterus typicus (рис. 3). Данные изменения связаны с разной глубиной отбора проб в разные периоды исследований: в 2006 г. пробы были отобраны на глубинах 107–79 м; в 2019 г. - на глубинах 105–127 м. Также снижение биомассы полихет в районе Новоземельского мелководья может быть связано с воздействием на донные биоценозы краба-стригуна опилио, увеличивающего свои максимальные поселения в данном районе.
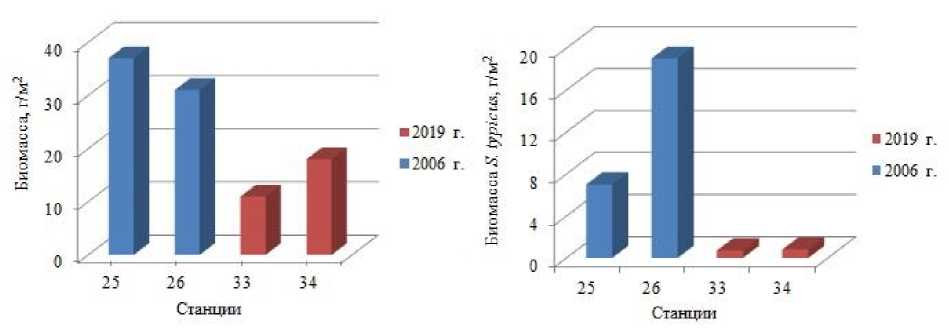
Рис. 3 Распределение общей биомассы полихет и биомассы Spiochaetopterus typicus на станциях в северо-западной части Новой Земли в 2006, 2019 гг.
Fig. 3. Distribution of biomass and biomass of Spiochaetopterus typicus at stations in the north-western part of Novaya Zemlya in 2006, 2019
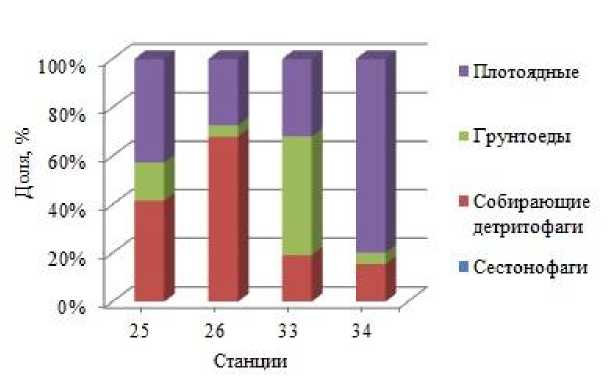
Рис. 4. Распределение биомассы трофических групп на станциях в северо-западной части Новой Земли в 2006, 2019 гг.
Fig. 4. Distribution of biomass of trophic groups at stations in the north-western part of Novaya Zemlya in 2006, 2019
Заключение
В результате проведенных исследований получены данные о видовом составе и количественных характеристиках полихет в северо-восточной части Баренцева моря. Изменение количественных характеристик и видового состава полихет в районе исследования обусловлено в первую очередь характером грунта, а также глубиной и придонной гидродинамикой.
В районах краевых желобов, покрытых мягкими илисто-песчаными, глинистыми грунтами, определены высокие количественные характеристики полихет доминирующего вида Spiochaetopterus typicus , образующего обильные поселения в данных районах.
Низкие значения количественных характеристик полихет отмечены в глубоководных районах, покрытых коричневыми илами с большим количеством раковин фораминифер, отличающихся неблагоприятным газовым режимом придонного слоя и низким содержанием питательных веществ.
Полученные данные существенно дополняют сведения о видовом составе и количественных характеристиках полихет северо-восточной части Баренцева моря и позволяют проследить за дальнейшими изменениями в составе и структуре донных сообществ.
Работа выполнена в рамках государственного задания Мурманского морского биологического института РАН "Донные биоценозы Баренцева моря, его водосборного бассейна и сопредельных вод в современных условиях" (номер государственной регистрации 122020900044-2). Автор выражает благодарность участникам экспедиции Гарбулю Е. А., Зиминой О. Л., Москвину К. К., Румянцевой З. Ю. за участие в сборе бентосных проб.
