Фенетический анализ краниологических признаков малой лесной мыши (Sylvaemus uralensis Pall.) в условиях загрязнения тяжелыми металлами в среднегорьях Центрального Кавказа
 в условиях загрязнения тяжелыми металлами в среднегорьях Центрального Кавказа")
Автор: Амшокова А.Х.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1-1 т.11, 2009 года.
Бесплатный доступ
Изучено влияние техногенного загрязнения и градиента высоты местности на морфогенез малой лесной мыши в условиях среднегорий Центрального Кавказа. Обнаружено, что высота над ур.м и связанные с ней условия среды оказывают на организм малой лесной мыши более неблагоприятные воздействия, соответственно выступают более стрессовыми, чем условия загрязнения отходами Тырныаузского вольфрамо-молибденового комбината. Косвенным подтверждением этому служит и относительно более высокий уровень флуктуирующей асимметрии (FАnm), выявленный у животных с наибольшей высоты местности (1800 м над ур. м). Предполагается, что сравнительно небольшой уровень FAnm в импактной выборке, по сравнению с контрольными, обусловлен, вероятно, популяционной адаптацией к антропогенно трансформированным условиям среды в связи с длительным обитанием малой лесной мыши (более 100 поколений) на территории, загрязненной техногенными отходами.
Фенетический анализ, краниологические признаки, лесная мышь, центральный кавказ
Короткий адрес: https://sciup.org/148198223
IDR: 148198223 | УДК: 575.17:599.323
Phenetic analysis of craniological characters in the lesser common field mouse (Sylvaemus uralensis Pall.) under heavy metals pollution in highland regions of the Central Caucasus
The effect of technogenic pollution and altitudinal gradient on morphogenesis of the lesser common field mouse in highland regions of the Central Caucasus has been studied. It's revealed that the altitude above sea level and related environments have even more unfavourable impact on the organism of the lesser common field mouse, and produce, correspondingly, much stress than industrial pollution from Tyrnyauz integrated tungsten-and-molybdenum works. The relatively higher level of fluctuating asymmetry (FAnm) found in animals from the highest altitude (1800 m a.s.l.) provides indirect support for this fact. It is suggested that the moderate level of FAnm in the impact sample, in comparison with control samples, is likely to be determined by the population adaptation to anthropogenically transformed environments in connection with prolonged habitation of the lesser common field mouse (more than 100 generations) in the territory polluted by technogenic wastes.
Текст научной статьи Фенетический анализ краниологических признаков малой лесной мыши (Sylvaemus uralensis Pall.) в условиях загрязнения тяжелыми металлами в среднегорьях Центрального Кавказа
Г орные экосистемы, характеризующиеся не только высоким разнообразием флоры и фауны, но и большой уязвимостью представляют значительный интерес для экологических исследований. В этом плане в пределах РФ особенно отличается Кавказская горная страна, где представлены все высотные пояса от низменностей до высокогорий. Высокая ультрафиолетовая радиация, низкое атмосферное давление, и связанное с этим низкое парциальное давление кислорода, разреженность воздуха, недостаток тепла, резкие суточные и сезонные перепады температуры и влажности накладывают отпечаток на образ жизни, морфологию и физиологию обитателей гор [1, 2]. В то же время имеющиеся на Центральном Кавказе предприятия по добыче и переработке молибдена и вольфрама (Тыр-ныаузский вольфрамо-молибденовый комбинат и Нальчикский гидро-металлургический завод) оказывают отрицательное влияние на хрупкие горные экосистемы, делая их еще более уязвимыми. Наличие комплекса естественного и антропогенного факторов в горах затрудняет проведение исследований и бывает достаточно сложно вычленить влияние того или иного фактора на живые организмы. Отмеченные факты определяют необходи-
Альбина Хасманова Амшокова, научный сотрудник лаборатории биоразнообразия позвоночных.
мость проведения исследований с учетом ряда факторов, таких как высота, рельеф местности, принадлежность к тому или иному типу поясности и т.д.
В последние десятилетия для характеристики состояния природных популяций широко используются фенетические методы, позволяющие судить о генетических изменениях, протекающих в популяциях по частотам неметрических пороговых признаков скелета. Важное значение для характеристики состояния природных популяций имеет и используемый в данной работе показатель флуктуирующей асимметрии, дающий оценку стабильности индивидуального развития. В.М. Захаровым [3] установлено, что повышение флуктуирующей асимметрии на групповом уровне указывает на дестабилизацию процесса развития в популяции.
Цель исследования — изучить изменчивость неметрических признаков черепа малой лесной мыши ( Sylvaemus uralensis Pall.), обитающей в зоне загрязнения отходами производства Тырныаузского вольфрамомолибденового комбината (ТВМК), с учетом высотного градиента.
МАТЕРИАЛ И МЕТОДЫ
Объект исследования — малая лесная мышь, широко распространенный и многочисленный вид на Северном Кавказе. По данным А.И. Ильенко и Т.П. Крапивко [4], экологические особенности вида (физиология, пищевая специализация, подвижность и т.д.), влияющие на величину накопления радионуклидов в организме, позволяют считать малую лесную мышь видом радиофором.
Материалом для исследования послужили серии черепов половозрелых особей, хранящихся в коллекционном фонде Института экологии горных территории КБНЦ РАН. Всего изучено 303 черепа мышей . Во избежание влияния высотного фактора на результаты сравнительного анализа контрольной (пос. Эльбрус — 1800 м) и импактной выборок (пос. Былым, хвостохранилище ТВМК — 1200 м), использовалась третья — контрольная (окр. пос. Гунделен), происходящая с такой же высоты, как и импактная. Выборкам даны номера и условные рабочие названия, соответствующие месту их сбора 1 (^) — 2 ( $$ ) эльбрусская, 3 (^) — 4 ( $$ ) гунделенская, 5 ( ^^ ) — 6 ( $$ ) бы-лымская. Все три пункта находятся в пределах восточно-северокавказского типа поясности [5].
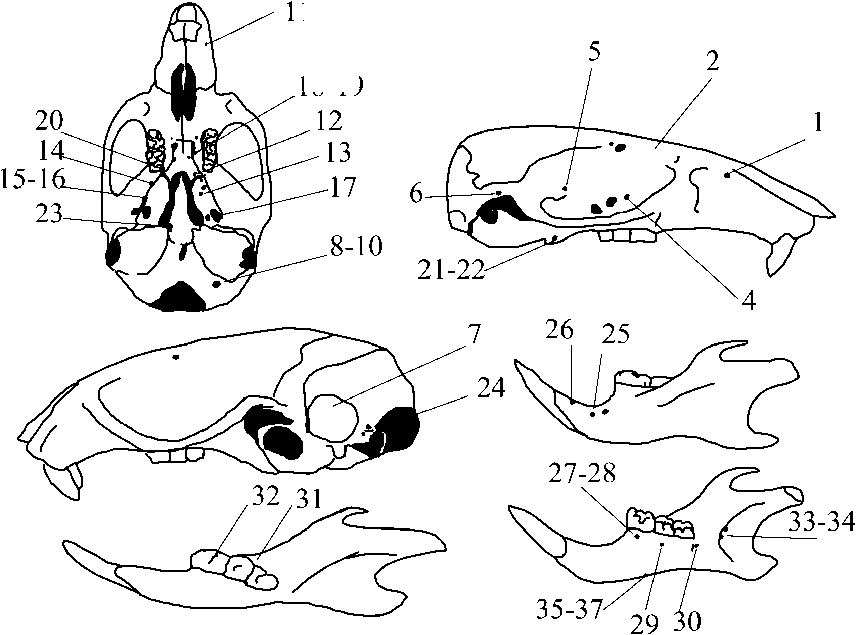
При проведении фенетического анализа (выделении, описании фенов, обработке результатов) использовали методы фенетики, разработанные и предложенные А.Г. Васильевым [6, 7]. Поиск и учет фенов проводили с помощью микроскопа МБС-10 при увеличении 2х8. Изначально было выявлено 83 дискретных состояния (фена) неметрических признаков. Большая часть фенов, используемых в работе, взята из литературных источников [8-11]. Связь проявления фенов с полом, возрастом, друг с другом оценивали на основе расчета непараметрических коэффициентов корреляции Спирмена. После проведения такой процедуры небольшую часть признаков, проявивших сильную значимую корреляцию с указанными факторами, исключили из дальнейшего анализа, что уменьшает вероятность получения артефактов за счет смещения оценок, обусловленных названными причинами, и повышает надежность косвенной генетической интерпретации фенетических различий [7]. Перечень 37 признаков, оставшихся после первичной выбраковки, приведен в табл. 1, а их расположение показано на рис. 1.
Фенетические дистанции (Mean Measure of Divergence — MMD) между выборками и их средние стандартные отклонения (Mean Standard Deviation — MSD) рассчитывали по формулам, предложенным С. Хартманом
[12]. Различия статистически значимы на уровне р < 0,05 при MMD > 2 MSD. Для сравнения выборок также использовали показатель средней уникальности выборок (MMU — Mean Measure of Uniqueness) — усредненная дистанция данной выборки со всеми остальными [11]. Уровень внутрипопуля-ционного разнообразия оценивали с помощью показателя ц , а концентрацию редких фенов с помощью h по Л.А. Животовскому [13]. Расчеты перечисленных параметров проведены с использованием пакета прикладных программ PHEN 3.0 [12].
КРАТКАЯ ХАРАКТЕРИСТИКА РАЙОНА ИССЛЕДОВАНИЙ
Поселок Эльбрус расположен в долине р. Баксан, географические координаты 43°15 ' с. ш. и 42°38 ' и в. д. Рельеф эльбрусского варианта имеет сравнительно мягкие очертания и относительно ровные платообразные формы с пологими северо-восточными и восточными склонами, в связи с этим сухие воздушные потоки Прикаспийской низменности не задерживаясь передовыми хребтами, легко достигают гор и обуславливают ксерофитизацию ландшафта и континентальность климата [5]. Основная площадь занята горно-луговыми дерновыми почвами формирующимися под субальпийскими лугами. Для растительного покрова характерны различные луговые ценозы. Древесно-кустарниковая растительность представлена в основном березовыми и сосновыми лесами, а также родоретами и зарослями субальпийских можжевельников [14]. Первичный материал собирался в окрестностях пос. Эльбрус на высоте около 18001900 м. н.у.м.
В окрестностях пос. Гунделен исследовали биотопы, расположенные в ущелье р. Тызыл (левого притока р. Баксан). Рассматриваемая территория, так же как и пос. Эльбрус, лежит в пределах эльбрусского варианта восточно-северокавказского типа поясности и относится к поясу остепненных лугов [5]. Рельеф местности гористый с высотами 10002000 м н.у.м., климат умеренный [15], с повышенным увлажнением в весенний и зимний периоды, туманами и неустойчивым снежным покровом. Почвы горно-луговые, местами заболоченные, растительность переходная от среднегорных лугов к субальпийским, местами значительная примесь степных элементов. Северо-восточные склоны ущелья на исследованных участках заняты лесом [16]. GPS координаты исследованной местности - 43°35 '
с. ш. и 43°50 ' в. д. Первичный материал собирался на высоте 1200 м н.у.м.
Импактная выборка (окр. пос. Былым) происходит из зоны хвостохранилища ТВМК, введенного в эксплуатацию в 1940 г. Накопленный объем промышленных хвостов к настоящему времени составляет более 110 млн. м3, отходы заполняют хвостохранилище на максимальную глубину 105 м. Часть хвостов (до 30%) находится в сухом состоянии, но большая часть - в жидком. Кроме вольфрама и молибдена, отходы содержат мышьяк, свинец, ртуть и другие тяжелые металлы. Их содержание на 2-3 порядка превышает ПДК в почвогрунтах и воде хвостохранилища. Несмотря на то, что Былымское месторождение находится в горной местности на высоте 1100
- 1300 не. Со
м н.у.м., оно расположено в котлови-всех сторон котловина окружена вы-1
сокими горами и только на север и на юг, прорезая ее, уходит Баксанское ущелье. Замкнутость котловины и определяет особые климатические условия. Для этой местности характерен сухой климат, с особо горностепным ландшафтом и типичными представителями флоры и фауны. По склонам карьера произрастает древесно-кустарниковая растительность: облепиха крушиновая, ива козья, береза повислая, абрикос обыкновенный и тамарикс. Травянистая растительность разнообразная: вейник наземный, душевка тимьянная, коровяк, липучка обыкновенная, полынь горькая, иван-чай кавказский, василек кубанский, ковыль волосатик, тысячелистник обыкновенный, пиретрум девичьелист-ный и т.д. Согласно типизации А.К. Тембо-това [17], исследуемая территория относится к эльбрусскому варианту поясности.
18-19
1. Расположение фенов неметрических признаков на черепе малой лесной мыши: 1 — удвоенное
Рис.
предглазничное отверстие; 2 — дополнительное лобное отверстие впереди основного; 3 — отсутствие лобного отверстия; 4 — одиночное нижнеглазничное отверстие; 5 — одиночное переднее отверстие чешуйчатой кости; 6 — одиночное отверстие височного хода; 7 — отсутствие окна на сосцевидной кости; 8 — одиночное подъязычное отверстие; 9 — утроенное подъязычное отверстие; 10 — боковое подъязычное отверстие; 11 — переднее боковое верхнечелюстное отверстие; 12 — одиночное отверстие в основании крыловидного отростка; 13 — удвоенное отверстие в крыловидной ямке; 14 — одиночное дополнительное круглое отверстие; 15 — удвоенное отверстие на перегородке между овальным и круглым отверстиями; 16 — утроенное отверстие на перегородке между овальным и круглым отверстиями; 17 — дополнительное отверстие в области овального отверстия с вентральной стороны в крыловидной ямке; 18 — одиночное дополнительное небное отверстие; 19 — более трех отверстий на небной кости; 20 — одиночное малое отверстие на уровне М 3 ; 21 — одиночное отверстие на крыловидном крючке; 22 — утроенное отверстия на крыловидном крючке; 23 — латеральное отверстие на основной клиновидной кости; 24 — множественные отверстия на затылочной кости в сочленовной ямке; 25 — дополнительное подбородочное отверстие впереди основного; 26 — нижнечелюстное отверстие II; 27 — одиночное отверстие с лингвальной стороны альвеолы Мр 28 — удвоенное отверстие с лингвальной стороны альвеолы Мр 29 — одиночное отверстие с лингвальной стороны альвеолы М 2 ; 30 — удвоенное отверстие с лингвальной стороны альвеолы М 3 ; 31 — удвоенное отверстие в области М 2 на внутренней поверхности; 32 — одиночное отверстие в области М 1 на внутренней поверхности; 33 — удвоенное дополнительное отверстие в крыловидной ямке нижней челюсти; 34 — более трех дополнительных отверстий в крыловидной ямке нижней челюсти; 35 — одиночное отверстие на «теле» нижней челюсти; 36 — удвоенное отверстие на «теле нижней челюсти»; 37 — более трех отверстий на «теле» нижней челюсти
Находясь в пределах одной широтной зоны и формируясь в сходных климатических и ландшафтных условиях, исследуемые выборки позволяют избежать влияния неоднородности высотно-поясной структуры горных ландшафтов.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Частота встречаемости фенов неметрических признаков. Во всех трех выборках просчитана непараметрическая корреляция Спирмена, позволившая выявить связь с полом для двух признаков — наличие более трех отверстий в крыловидной ямке и наличие одинарного дополнительного отверстия в крыловидной ямке нижней челюсти. Далее оценена связь встречаемости признаков с возрастом также во всех трех выборках при сравнении животных трех возрастных групп: subadultus, adultus, senex, при этом возрастные различия проявились по шести признакам. Различия обнаружены и по встречаемости признаков на левой и правой сторонах черепа, т.е. связь со стороной тела (направленная асимметрия) выявлена по восьми признакам. Около 37 % признаков оказались связанными друг с другом. Все перечисленные признаки были удалены из дальнейшего анализа. В итоге из 83 первоначально выделенных признаков расчеты проводили по 37 (табл. 1).
Таблица 1
Частоты встречаемости фенов неметрических признаков черепа в контрольных и импактной выборках малой лесной мыши, %
|
Номер и код признака |
Эльбрус (контроль) |
Гунделен (контроль) |
Былым (импактная) |
Сравниваемые пары выборок |
||||||||
|
1 |
2 |
3 |
1 4 |
5 |
6 |
Хи-квадрат |
||||||
|
ss |
?? |
ss |
?? |
ss |
?? |
|||||||
|
n=130 |
n=114 |
n=36 |
n=3 8 |
n=144 |
n=144 |
1-3 |
1-5 |
3-5 |
2-4 |
2-6 |
4-6 |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
1. FPodu |
7,7 |
2,6 |
8,3 |
7,9 |
2,1 |
3,5 |
||||||
|
2. FFracan |
4,6 |
3,5 |
2,8 |
7,9 |
2,1 |
0,7 |
* |
|||||
|
3. FFracan (-) |
3,9 |
3,5 |
5,6 |
2,6 |
4,9 |
5,6 |
||||||
|
4. FIO |
67,7 |
79,0 |
47,2 |
36,1 |
40,6 |
34,5 |
*** |
* |
*** |
*** |
||
|
5. FTmacan |
2,3 |
0,9 |
0 |
5,3 |
2,1 |
0,7 |
||||||
|
6. MeTm |
13,9 |
17,5 |
25 |
13,2 |
18,8 |
18,1 |
*** |
|||||
|
7. FeMs (-) |
47,7 |
36,0 |
16,7 |
0 |
20,1 |
11,1 |
* |
*** |
*** |
*** |
** |
|
|
8. FHg |
70,3 |
57,1 |
86,1 |
63,9 |
57,5 |
60,6 |
*** |
|||||
|
9. FHgtr |
2,3 |
1,8 |
0 |
0 |
1,4 |
2,1 |
||||||
|
10. FHgla |
3,9 |
1,8 |
0 |
0 |
9,7 |
5,6 |
* |
|||||
|
11. FPmla |
0,8 |
4,4 |
0 |
0 |
1,4 |
1,4 |
||||||
|
12. FPT |
5,4 |
8,8 |
19 , 4 |
21,1 |
9,0 |
5,6 |
* |
** |
||||
|
13. FRTI |
22,3 |
14,9 |
8,3 |
13,2 |
20,1 |
16,8 |
||||||
|
14. FPD |
23,1 |
31,6 |
22,2 |
39,5 |
29,2 |
37,5 |
||||||
|
15. FLTIdu |
23,1 |
24,6 |
13,9 |
13,2 |
21,5 |
11,8 |
** |
|||||
|
16. FLTItr |
3,9 |
3,5 |
2,8 |
5,3 |
3,5 |
2,1 |
||||||
|
17. Fasac |
30,0 |
40,4 |
16,7 |
5,3 |
23,6 |
8,3 |
*** |
*** |
* |
|||
|
18. FPl |
22,3 |
26,3 |
22,2 |
23,7 |
32,6 |
43,8 |
** |
|||||
|
19. FPl(mx) |
3,1 |
1,8 |
0 |
0 |
2,1 |
2,1 |
||||||
|
20. FPLmn |
12,3 |
9,7 |
0 |
5,3 |
7,6 |
7,6 |
** |
* |
||||
|
21. FHP |
9,7 |
9,3 |
6,25 |
11,1 |
12,1 |
14,1 |
||||||
|
22. FHPtr |
1,5 |
0,9 |
2,8 |
2,6 |
2,8 |
2,8 |
||||||
|
23. FBsla |
36,9 |
41,6 |
13,9 |
13,2 |
30,6 |
29,2 |
** |
* |
* |
** |
* |
|
|
24. FFsOc(mx) |
66,2 |
68,4 |
61,1 |
42,1 |
48,6 |
47,9 |
** |
*** |
** |
|||
|
25. FMTA |
10,0 |
4,5 |
8,3 |
15,8 |
5,0 |
8,3 |
* |
|||||
|
26. FMDS |
0,0 |
0,0 |
0 |
0 |
2,1 |
0,0 |
||||||
|
27. FMtlg |
33,9 |
35,4 |
16,7 |
28,9 |
16,7 |
20,4 |
** |
* |
** |
|||
|
28. FMtlgdu |
1,5 |
5,3 |
2,8 |
0 |
0,0 |
0,0 |
** |
|||||
|
29. FMblg |
23,1 |
22,1 |
33,3 |
42,1 |
26,4 |
34,5 |
* |
* |
||||
|
30.FMblgpodu |
24,6 |
20,4 |
38,9 |
26,3 |
12,5 |
19,4 |
** |
*** |
||||
Окончание табл. 1
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
31. FPCdu (M3) |
26,9 |
27,2 |
8,3 |
10,5 |
12,5 |
13,9 |
** |
* |
** |
* |
||
|
32. FPC (M 1 ) |
0,0 |
0,0 |
2,8 |
0 |
1,4 |
2,1 |
||||||
|
33. FMbmst |
3,9 |
8,8 |
8,3 |
2,6 |
2,8 |
7,7 |
||||||
|
34. FMbmst (mx) |
4,0 |
3,9 |
0 |
0 |
15,0 |
7,7 |
** |
*** |
* |
|||
|
35. FOM |
10,8 |
7,9 |
10 |
5,3 |
9,9 |
11,2 |
||||||
|
36. FOMdu |
19,2 |
14,0 |
11,1 |
13,2 |
21,8 |
18,8 |
||||||
|
37. FOMtr |
16,9 |
21,1 |
55,6 |
63,2 |
16,7 |
13,9 |
*** |
*** |
*** |
*** |
Примечание : N — число изученных сторон черепа. Уровень значимости межгрупповых различий по отдельным признакам (Хи-квадрат): * — p < 0,05; ** — p < 0,01; *** — p < 0,001.
При сравнении особей разного пола для каждой из трех сравниваемых выборок оказалось, что половой диморфизм по частоте встречаемости отдельных признаков наиболее выражен в импактной выборке. Достоверные различия обнаружены по следующим признакам: 7, 15, 17, причем эти признаки обнаружены чаще у самцов нежели самок (табл. 1). В эльбрусской выборке подобные различия наблюдаются только по одному признаку 8, а в гунделенской выборке по двум - 7 и 8, частота встречаемости отмеченных признаков также выше у самцов, чем самок.
Попарное сравнение самцов трех выборок показало, что в эльбрусской выборке доля самцов обладающих признаками 4, 7, 24, 27, 31 достоверно выше, чем в былымской и гунделенской выборках. По сравнению с другими у животных импактной зоны чаще встречаются признаки 10 и 34, по частоте встречаемости этих признаков самцы данной выборки достоверно превосходят особей двух других. К числу подобных у самцов из окрестностей пос. Гунделена можно отнести признаки 8, 12, 30, 37. Необходимо также отметить, что у самцов отмеченной выборки полностью отсутствуют признаки 5, 9, 10, 11, 19, 20, 34, тогда как в других выборках они хотя и с низкой частотой, но все-таки встречаются. Аналогичное сравнение самок из контрольной (эльбрусская) и импактной выборок выявило значимые различия по частоте встречаемости девяти признаков: 4, 7, 15, 17, 23, 24, 27, 28, 31. Высокий процент самок обладающих этими признаками, отмечен в пос. Эльбрус. Причем, как видно из табл. 1, признаки 4, 7, 24, 27, 31 были отмечены и у самцов, из чего можно предположить, что данные признаки маркируют эльбрусскую выборку в целом. Самки импактной выборки отличаются от остальных относительно высокой частотой встречаемости одного признака - 18 (43.8%), что почти в 2 раза превышает его встречаемость в других выборках (26,3% и 23,7 %). В окрестностях Гунделена встречаются самки с признаками 2 (7,9%), 12 (21,1%), 37 (63,2 %), что заметно выше по сравнению с частотой встречаемости этих признаков у самок импактной выборки - 0,7%, 5,6%, 13,9 %. Практически все перечисленные признаки для самцов гунделенской выборки (за исключением 4 и 20) отсутствуют и у самок (табл. 1).
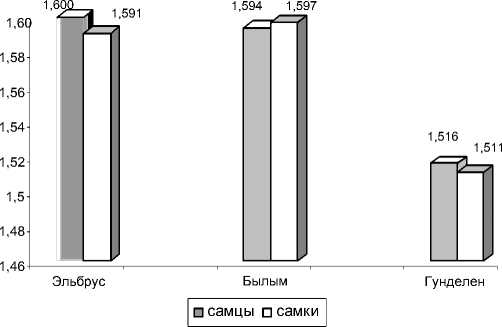
Рис. 2. Уровень фенетического разнообразия самцов и самок трех выборок малой лесной мыши
Таблица 2
Фенетические MMD-дистанции (верхняя треугольная матрица), среднее стандартное отклонение — MSD (нижняя треугольная матрица) и средняя уникальность (MMU) трех выборок малой лесной мыши на Центральном Кавказе
|
Выборки |
пол |
Эльбрус (1800 м) |
Гунделен (1200 м) |
Былым (1200 м) |
MMU |
|||
|
33 |
?? |
33 |
?? |
33 |
?? |
|||
|
Эльбрус |
33 |
- |
0,0020* |
0,0596 |
0,1117 |
0,0362 |
0,0585 |
0,055 |
|
?? |
0,0038 |
- |
0,0658 |
0,1170 |
0,0446 |
0,0687 |
0,060 |
|
|
Гунделен |
33 |
0,0082 |
0,0085 |
- |
0,0017* |
0,0556 |
0,0484 |
0,046 |
|
?? |
0,0079 |
0,0081 |
0,0125 |
- |
0,0686 |
0,0461 |
0,069 |
|
|
Былым |
33 |
0,0034 |
0,0037 |
0,0080 |
0,0077 |
- |
0,0068 |
0,042 |
|
?? |
0,0034 |
0,0037 |
0,0080 |
0,0077 |
0,0032 |
- |
0,046 |
|
Примечание : * - различия статистически недостоверны.
Результаты сравнения животных по показателю внутрипопуляционного разнообразия -р Л.А. Животовского приведены на рис. 2. Видно, что наибольший уровень фенетиче-ского разнообразия наблюдается у самцов контрольной (пос. Эльбрус) и импактной выборок. Схожая картина вырисовывается и при сравнении самок, что в свою очередь позволяет говорить о повышенном фенетиче-ском разнообразии контрольной выборки происходящей с наибольшей высоты местности 1800 м н.у.м. (пос. Эльбрус) по сравнению с двумя другими.
Для оценки уровня феногенетических различий между контрольными и импактной выборками были вычислены фенетические дистанции (MMD) и средняя мера уникальности (MMU) выборок. Результаты, приведенные в табл. 2, показывают, что средняя фенетиче-ская дистанция, выявленная в двух контрольных выборках при сравнении особей разного пола оказалась статистически недостоверной и составила в эльбрусской выборке 0,0020±0,0038, а в гунделенской 0,0017±0,0125. В импактной выборке напротив различия между полами оказались статистически значимыми MMD = 0,0068±0,0032. Межгрупповые различия по полу проявились во всех вариантах сравнения, как у самцов, так и самок. Так, при сравнении самцов максимальные значения MMD = 0,0596±0,0082 обнаружены между контрольными выборками, располагающимися на высотном удалений друг от друга в 600 — 700 м. Минимальные значения MMD = 0,0362±0,0034 отмечены для самцов эльбрусской и импактной выборок .
Между двумя выборками, располагающимися на одном высотном уровне, но в биото пах с разным уровнем антропогенной нагрузки, отмечены также значения MMD, близкие к максимальным — 0,0556±0,0080. Аналогичное сравнение самок также позволяет говорить о большей фенетической удаленности пары контрольных выборок MMD = 0,1170±0,0081, причем значения полученные для самок почти в 2 раза превышают таковые самцов. Самки импактной и контрольной (эльбрусской) выборок также находятся на большем феногенетическом уклонении друг от друга MMD = 0,0687±0,0037 по сравнению с особями противоположного пола этих же выборок. Исключение составляет значение MMD = 0,0461±0,0077 самок гунделенской и импактной выборок, которое немного ниже чем у самцов.
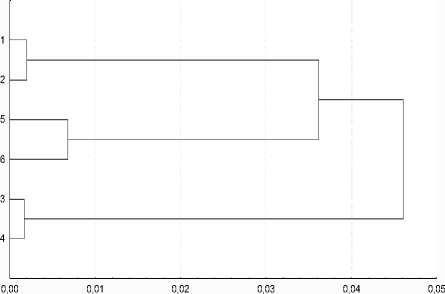
Кластерный анализ матрицы MMD-дистанций также показал, что половые различия выражены наиболее сильно в импактной группе, что касается уровня фенетиче-ских различий между выборками, здесь четко видно, что наиболее удаленными являются контрольные выборки и напротив более близки к друг другу импактная и эльбрусская выборки, образующие один кластер (рис. 3).
Средняя мера фенетической уникальности выборок варьировала от 0,042 до 0,069, при этом наиболее своеобразными в фенетическом отношении оказались самки всех трех выборок.
Флуктуирующая асимметрия. Оценку флуктуирующей асимметрии (ФА) у зверьков из техногенной и контрольной зоны отдельно для каждого пола проводили по коэффициенту FAnm (средней доле ассимет-рично проявившихся неметрических признаков на особь), данные приведены в табл. 3. Значимость различий между выборками оце- нивали с помощью метода непараметрической статистики Краскела-Уоллиса [7].
Фенетическая дистанция
Рис. 3. Кластерный анализ (UPGMA) изученных выборок малой лесной мыши на основе матрицы MMD — дистанций: 1( ^^ ) — 2 ( $$ ) эльбрусская, 3 ( ^^ ) — 4 ( $$ ) гунделенская, 5 ( ^^ ) — 6 ( $$ ) былымская
Оценивали FAnm отдельно для самцов и самок путем сравнения трех выборок. При этом достоверные различия выявлены между самками импактной и контрольной (эльбрусской) H = 11,91; N = 129; p = 0,0006, а также самками двух контрольных выборок H = 7,41; N = 76; p = 0,007. Интересно, что между самками из Гунделена и Былыма достоверных различий не обнаружено. У самцов не выявлено значимых различий во всех вариантах сравнения.
Таблица 3
Уровень флуктуирующей асимметрии (FAnm, %) самцов и самок из трех выборок
|
Выборки |
N |
FAnm |
cv% |
О |
|
1. Эльбрус |
65 |
23,1±0,91 |
31,6 |
7,31 |
|
2. Эльбрус |
57 |
25,2±0,98 |
29,2 |
7,37 |
|
3. Гунделен |
18 |
21,1±1,82 |
36,6 |
7,71 |
|
4. Гунделен |
19 |
19,5±1,82 |
40,8 |
7,94 |
|
5. Былым |
72 |
22,9±1,07 |
39,5 |
9,04 |
|
6. Былым |
72 |
20,5±0,98 |
40,4 |
8,30 |
ЗАКЛЮЧЕНИЕ
Полученные результаты свидетельствуют о сопоставимом влиянии на морфогенез малой лесной мыши, как техногенного загрязнения, так и высоты местности. Общеизвестно, что с подъемом в горы общий уровень радиации увеличивается. Видимо, этим и можно объяснить более низкую степень фенетических различий выявленный между импактной (пос. Былым) и контрольной (пос. Эльбрус) выборками, находящимися на хорошем высотном удалений (700 м) друг от друга. Воз можно, что различия накладываемые высотой местности при сравнении этих выборок нивелируются присутствием повышенного уровня содержания тяжелых металлов в отходах ТВМК, под воздействием которых находятся природные компоненты, в том числе и мелкие млекопитающие, импактной зоны. Наиболее четко влияние высоты местности и сопровождающих ее условий среды можно проследить при сравнении двух контрольных выборок (поселки Эльбрус и Гунделен). Несмотря на то, что эти выборки находятся на таком же высотном удалении, как и предыдущие две, тем не менее уровень фенетических различий между ними значительно выше и составил для самцов MMD = 0,0596±0,0082, а самок MMD = 0,117±0,0081 (табл. 2). Можно предположить, что фенети-ческие различия в данном случае происходят в результате отсутствия сглаживающего (сближающего эти выборки) фактора, в качестве которого в первом варианте сравнения выступает загрязнение в техногенно нарушенных экосистемах.
Следующий аспект исследования был связан с изучением степени влияния техногенного загрязнения на исследуемые выборки. Изменчивость неметрических признаков черепа малой лесной мыши в условиях загрязнения изучали, сравнивая гунделенскую и импакт-ную выборки, находящиеся на одном высотном уровне (1200 м). Фенетические дистанции между отмеченными выборками по частотам фенов оказались статистически высоко значимыми для особей обоего пола и составили 0,0556 у самцов и 0,0461 у самок.
Таким образом, полученные результаты позволяют заключить, что исследуемые факторы (высота и техногенное загрязнение), оказывают существенное влияние на изменчивость неметрических признаков черепа малой лесной мыши. В рассматриваемом случае высотная разница, составляющая 600-700 м, служит фактором, усиливающим уровень дифференциации между выборками, причем приводя к большей взаимной фенетической удаленности самок, нежели самцов.
Результаты сравнительного анализа изученных выборок по уровню флуктуирующей асимметрии оказались противоположными. Так, установлено, что минимальное значение FAnm характерно для животных из окрестностей пос. Гунделен, а максимальное — для мышей из окрестностей пос. Эльбрус. Относительно более высокий уровень FAnm, как у самцов, так и самок эльбрусской выборки, указывает на снижение стабильности индивидуального развития в этой выборке. При этом не наблюдается существенного преобладания величин FAnm в импактной выборке. По данному показателю особи обоих полов импактной выборки слабо отличаются от таковых контрольной выборки с той же высоты над уровнем моря. Самки контрольной выборки с большей высоты местности из окрестностей пос. Эльбрус, характеризуются существенно (t = 3,4) более низким показателем FAnm в сравнении с самками техногенно загрязненной территории. У самцов различия не выявлены.
Как следует из данных литературы [18-25] на нарушение стабильности развития у растений и животных оказывают влияние неблагоприятные условия среды обитания.
В контексте изложенного выше можно заключить следующее. Во-первых, высота над уровнем моря и связанные с ней условия среды оказывают на организм малой мыши более неблагоприятные воздействия, соответственно, выступают более стрессовыми, чем условия загрязнения отходами ТВМК, что более присуще самкам.
Во-вторых, относительно небольшой уровень FAnm в импактной выборке в сравнении с контрольной выборкой происходящей с той же высоты местности, вероятно, обусловлен длительным обитанием малой лесной мыши (более 100 поколений) на территории, загрязненной техногенными отходами, и, возможно, связан с популяционной адаптацией к антропогенно трансформированным условиям обитания. Полученные данные согласуются с таковыми по красной полевке, обитающей на территории Восточно-Уральского радиоактивного следа (ВУРС) в течение по крайне мере 100 поколений с момента Кыштымской аварии на Южном Урале [7, 8]. Авторы установили, что импактная популяция Clethrionomys rutilus не отличается по индексу FAnm от контрольных, и «ведет себя» как «нормальная популяция». На основании чего было сделано заключение, что в импактной популяции уже возникли устойчивые эпигенетические изменения в организации развития, а сам процесс развития близок к нормальному протеканию в новом эпигенетическом русле [8].
Выражаю искреннюю благодарность д.б.н., зав. лабораторией ИЭРиЖ УрО РАН А.Г. Васильеву и д.б.н. И.А. Васильевой за неоценимую помощь при освоении методики и постоянные консультации. Также хочу поблагодарить своего научного руководителя чл.-корр. РАН Ф.А. Темботову за всестороннюю помощь и поддержку на всех этапах работы.
Список литературы Фенетический анализ краниологических признаков малой лесной мыши (Sylvaemus uralensis Pall.) в условиях загрязнения тяжелыми металлами в среднегорьях Центрального Кавказа
- Калмыкова В.Г. Некоторые специфические черты растений и животных горных территорий//Проблемы географии. Калинин,1975.
- Булатова Н.Н. Особенности высокогорных животных//Тр. ИЭМЖ. 1962. Вып. 41.
- Захаров В.М. Асимметрия животных. М.: Наука, 1987.
- Ильенко А.И. Крапивко Т.П. Экологические последствия радиоактивного загрязнения на популяции мелких млекопитающих -стронциефоров//Экологические последствия радиоактивного загрязнения на Южном Урале. М.: Наука, 1993.
- Соколов В.Е., Темботов А.К. Позвоночные Кавказа: Насекомоядные. М.: Наука, 1989.
- Васильев А.Г. Пакет прикладных программ PHEN 3.0. 1995. (http://ecoinf.uran.ru)
- Васильев А.Г. Эпигенетические основы фенетики: на пути к популяционной мерономии. Екатеринбург: Изд-во «Академкнига», 2005.
- Васильев А.Г., Васильева И.А., Большаков В.Н. Фенетический мониторинг популяций красной полевки (Clethrionomys rutilus Pall.) в зоне Восточно-Уральского радиоактивного следа//Экология. 1996. № 2.
- Песков В.Н., Емельянов И.Г. Фенетика и феногеография водяной полевки (Arvicola terrestris L.)//Вест. зоологии. 2000. № 3.
- Песков В.Н., Емельянов И.Г. Фенетический анализ популяционной структуры водяной полевки (Arvicola terrestris L.) Центральной и Восточной Палеарктики//Вест. зоологии. 2000. № 6.
- Васильева И.А., Васильев А.Г., Любошевский Н.М. и др. Феногенетический анализ популяций малой лесной мыши (Apodemus uralensis Pall. 1811) в зоне влияния Восточно-Уральского радиоактивного следа//Экология. 2003. № 6.
- Hartman S.E. Geographic variation analysis of Dipodomys ordii using nonmetric cranial traits//J. Mammal. 1980. V. 61. № 3.
- Животовский Л.А. Показатели популяционной изменчивости по полиморфным признакам//Фенетика популяций. М.: Наука, 1982.
- Гроссгейм А.А. Растительный покров Кавказа. М.: МОИП. 1948.
- Ёжкина А.Н. Растительность северного склона Скалистого хребта между рр. Баксаном и Тызылом//Уч. Зап. Сер. Биол. Вып.16. Нальчик. 1962.
- Рапопорт И.Б. Первые сведения о дождевых червях ущелья реки Тызыл (Центральный Кавказ)//Горные экосистемы и их компоненты: Труды междунар. конф. М.: Т-во науч. изд. КМК, 2007.
- Темботов А.К., Шебзухова Э.А., Темботова Ф.А., Темботов А.А., Ворокова И.Л. Проблемы экологии горных территорий. Учебное пособие. Майкоп: Издательство АГУ. 2001.
- Гилева Э.А., Нохрин Д.Ю. Флуктуирующая асимметрия краниометрических признаков у восточноевропейской полевки из зоны радиационного неблагополучия//Экология. 2001. № 1.
- Кряжева Н.Г., Чистякова Е.К., Захаров В.М. Анализ стабильности развития березы повислой в условиях химического загрязнения//Экология. 1996. № 6.
- Егоров Д.О., Егорова А.О. Оценка уровня загрязненности окружающей среды с использованием биоиндикаторов//«Современные проблемы экологии, микробиологии и иммунологии»: Пермь, 1999.
- Шадрина Е.Г., Протопопова В.В. Реакция насекомоядных млекопитающих на техногенные воздействия в Центральной Якутии//Биология насекомоядных млекопитающих: Тез. докл. междунар. конф. Кемерово. 1999.
- Шадрина Е.Г., Шадрин Д.Я. Показатели стабильности развития мелких млекопитающих Западной Якутии в зоне антропогенного воздействия//Териофауна России и сопредельных территорий: Материалы Междунар. совещ. Москва, 2003.
- Макеева Т.И., Никонова Г.Н. Оценка антропогенной нагрузки на территории по показателям стабильности развития растений//Проблемы и пути их решения: Материалы науч.-практ. конф.: Москва, 30-31 окт., 2002. конференции. М., 2002.
- Силкин А.А., Логинов В.В., Слепов А.В., Мокров И.В. Стабильность развития популяций мелких млекопитающих, амфибий и растений как показатель качества среды пойм малых рек//Малые реки: Современное экологическое состояние, актуальные проблемы: Междунар. научн. конф. Тольятти: ИЭВБ РАН, 2001.
- Устюжанина О.А. Биоиндикационная оценка качества окружающей среды по стабильности развития и фенетике бесхвостых амфибий Rana ridibunda, R. lessonae, R. esculenta, R. temporaria//Калуга, 2002.
