Феномен засухоустойчивости видов рода Salix L.: экспериментальная характеристика особенностей водного режима

Автор: Кулагин А.Ю.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биология и экология
Статья в выпуске: 2 т.5, 2003 года.
Бесплатный доступ
Особенности водного режима обусловливают успешность произрастания отдельных видов во внепойменных техногенных ландшафтах. Установлены значительные различия между широко распространенными видами ив по транспирации и обратимому водному дефициту листьев ив, по зимней водоудерживающей способности одревесневших побегов. Показана лабильность показателей водного режима ив.
Короткий адрес: https://sciup.org/148197757
IDR: 148197757 | УДК: 574
Drought resistance phenomena of the Salix L. species: the experimental characteristic of features of water regime
The features of water regime of willows determine the growth successfulness of individual species in noninundated technogenous landscapes. The remarkable differences between widespread willows species on transpiration and convertible water deficit of willows leaves, on winter soil moisture retention capacity of ligneous shoots have been determined. There is indicated the predictors lability of water regime of willows.
Текст научной статьи Феномен засухоустойчивости видов рода Salix L.: экспериментальная характеристика особенностей водного режима
Институт биологии Уфимского научного центра РАН
Особенности водного режима обусловливают успешность произрастания отдельных видов во внепойменных техногенных ландшафтах. Установлены значительные различия между широко распространенными видами ив по транспирации и обратимому водному дефициту листьев ив, по зимней водоудерживающей способности одревесневших побегов. Показана лабильность показателей водного режима ив.
В последние годы исследования водного режима растений переместились на уровень изучения механизмов водопроведения и регуляции водного обмена в лесных экосистемах (Молчанов, Молчанова, 2001). Между тем, выявление особенностей водного режима с учетом видовых особенностей позволяет прогнозировать успешность их произрастания в экстремальных условиях (Кулагин, 1998).
Эколого-биологическая характеристика древесных пород, анализ отношения их к условиям загрязнения позволяют отметить, что засухоустойчивые и солеустойчивые древесные одновременно являются и газоустойчивыми (Илюшин, 1953; Кулагин, 1974; Кондратюк и др., 1980). В.С.Николаевский (1979) отмечает, что водоудерживающая способность является показателем устойчивости растений к экстремальным факторам (низкие температуры, засуха, газы). Отмечено, что «ивовые генетически приспособлены к избыточному увлажнению» (Смирнов, 1972, с.121). Однако ранее (Скворцов, 1961) указывалось, что неправильно представление об ивах, как крайне гигрофильных растениях. Ивы в защитных лесополосах в условиях сухой степи по засухоустойчивости не уступают тополю бальзамическому (Populus balsamifera L.) (Савельева, 1976). А.У.Усма-нов (1973) считает, что ивы можно отнести к мезофитам. Действительно, ивы способны выносить большую сухо сть почвы, что
В.Н.Сукачев (1952) связывал с наличием мощной, глубоко идущей корневой системы, которая со второго-третьего года после посадки черенков обеспечивает засухоустойчивость растений.
С целью объяснения феномена поселения и успешного произрастания ив во вне-пойменных ландшафтах техногенного происхождения, где отдельные виды ив проявляют определенную засухоустойчивость, в сравнительном плане были изучены особенности водного режима. Особенно важно учитывать особенности водного режима древесных растений при создании насаждений на промышленных отвалах, где в летний период высока вероятность пересыхания, а зимой - вымерзания почвогрунтов (Баталов и др., 1989).
Водный режим характеризовался следующими показателями: обратимый водный дефицит листьев, утренний водный дефицит, полуденный водный дефицит, транспирация листьев, зимняя водоудерживающая способность однолетних побегов (Цельникер, 1955; Острикова, 1966; Penka, 1967; Генкель, 1982; Elias, 1979; Garnier et al, 1988).
Для изучения особенностей водного режима ив использовались 4-5-летние растения, произрастающие на экспериментальном участке лаборатории лесоведения (территория Ботанического сада-института УНЦ РАН, г.Уфа).
Для определения обратимого водного дефицита листьев с черешком, предварительно взвешенные на торзионных весах марки ВТ, подсушивали в лабораторном помещении при температуре 20-25 С и относительной влажности воздуха 70-80% на рассеянном свету в течение 1-5 часов (листья располагались морфологически нижней поверхностью вверх). Затем листья взвешивали и помещали на 3 часа в эксикатор с водой. В эксикаторе листья, укрепленные черешками в пенопластовом плотике, восстанавливали потерянную воду. После этого листья, обсушенные фильтровальной бумагой, взвешивали в третий раз. Затем производился расчет веса потерянной и восстановленной воды в процентах к исходному весу листьев. Подобный метод применялся для определения засухоустойчивости древесных растений и в связи с газоус-тойчивостью (Сидорович, Гетко, 1979).
Утренний и полуденный водный дефицит листьев (за основу принят метод Ю.Л.-Цельникер, 1955) определяли в ясную погоду с 08-00 до 10-00 (температура окружающего воздуха в среднем 15,8 С) и с 12-00 до 14-00 (температура - 19,9 С); водонасыщение листьев проводили в эксикаторе на рассеянном свету в течение одного часа.
Транспирация листьев определялась методом быстрого взвешивания на торзионных весах (Иванов и др., 1950) с последующим экспонированием на рассеянном свету в течение 3 мин и повторным взвешиванием. Расчет интенсивности транспирации в мг воды на 1 г сырых листьев за 1 час (мг/г час).
Опыты по зимнему водному режиму ив проводилась с целью выявления видов, которые наиболее устойчивы к зимнему иссушению. Целесообразность проведения таких экспериментов связана с тем, что в зимнее время на отвалах нередко наблюдается ограниченное снегонакопление и общая низкая вла-гоемкость отвальных грунтов приводит к формированию весьма жестких для растений условий зимовки и весенней вегетации. Изучение динамики содержания воды проводили на однолетних побегах. За исходную была принята обводненность побегов в момент срезки их с растений. Начало опыта - декабрь, то есть время, когда растения уже выходят из глубокого периода покоя (Сабинин, 1963).
Опытные побеги подвешивались над поверхностью снега на высоте 1-2 м на открытом воздухе; пробы побегов на обводненность и жизнеспособность брались через 15 дней. После определения первоначального веса побеги высушивались в термо стате при 105 С. Обводненность рассчитывали по разнице веса до и после высушивания. Критерий жизнеспособности растений при определенном уровне обводненности - распускание листьев при помещении побегов в воду в лабораторном помещении. В каче стве контрольных использовали побеги из крон растений.
В результате проведения экспериментов было установлено, что утренний водный дефицит листьев (таблица 1) выражен у ивы белой (Salix alba L.), ивы козьей (S.caprea L.), ивы корзиночной (S.viminalis L.) и отсутствует у ивы шерстистопобеговой (S.dasyclados Wimm.). К полудню водный дефицит несколько возрастает, но различия между видами сглаживаются. Наиболее интенсивная транспирация утром отмечена у ивы белой. В утренние часы наименьшая транспирация отмечается у ивы серой (S.cinerea L.) и ивы трехтычинковой (S.triandra L.). В полдень интенсивность транспирации ивы трехтычинковой (форма одноцветная) достигает максимальной величины в ряду изученных видов ив, а наименьшая потеря воды отмечается у ивы серой. К вечеру транспирация у ивы шер-стистопобеговой резко снижается, в то время как у других ив снижение транспирации незначительно (ива серая, ива козья) или даже отмечается повышение ее интенсивности (ива трехтычинковая форма двуцветная). В ряду изученных видов ива остролистная (S.acutifolia Willd.) характеризуется средними значениями показателей водного режима листьев. В целом величины интенсивности транспирации и водного дефицита листьев ив согласуются с опубликованными данными И.А.Смирнова (1972).
Водоудерживающая способность является показателем устойчивости растений к экстремальным факторам (Николаевский, 1979), показателем степени засухоустойчивости (Смирнов, 1972). Опыты по обратному
Таблица 1. Некоторые показатели водного режима листьев видов рода Salix L. (средние значения)
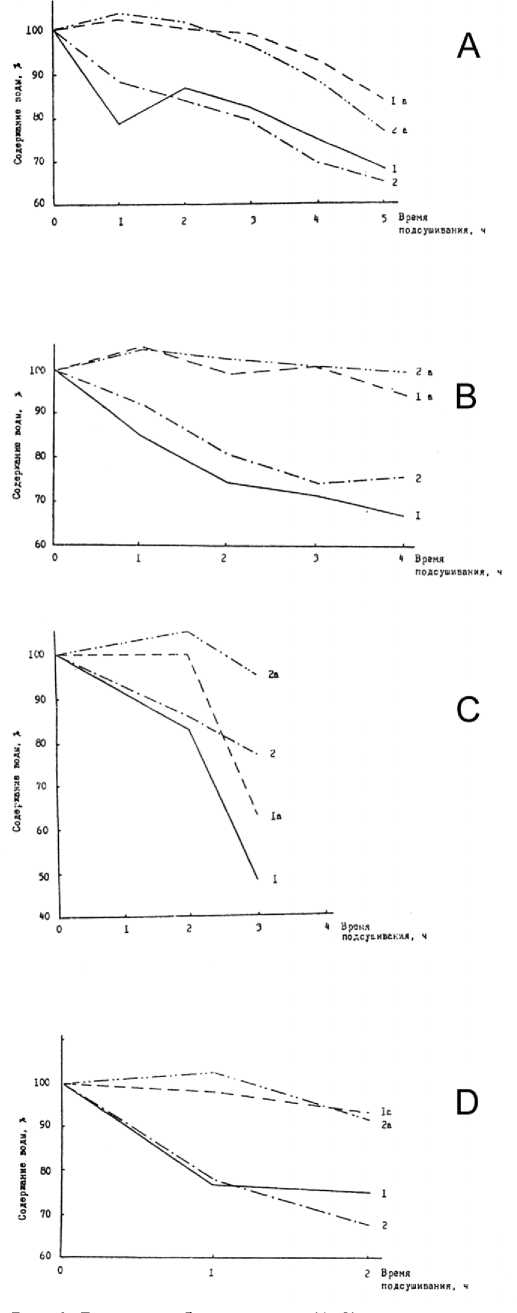
Ивы серая и козья успешно выдерживают 1-, 2-, 3-, 4-часовое подсушивание (рис.1), что проявляется в восстановлении потерянной воды. Однако следует указать, что ива козья имеет несколько повышенную водоудерживающую способность. Наличие воскового налета на нижней стороне листа позволяет считать форму двуцветную более ксе-роморфной в сравнении с формой одноцветной ивы трехтычинковой. В опытах по обратимому водному дефициту листьев (рис.1) выявилась относительная ксерофитность форм двуцветной. Двухчасовое подсушивание листья обеих форм перенесли успешно и восстановили водный дефицит. Трехчасовое подсушивание форма двухцветная переносит хорошо, а форма одноцветная не восстановила потерянную воду даже наполовину.
Ивы белая и остролистная незначительно различаются по обратимому водному дефициту листьев (рис.1) как между собой, так и от других видов ив. Можно отметить, что при двухчасовом подсушивании листья ивы остролистной обезвоживаются в большей степени по сравнению с ивой белой.
Опыты по зимней водоудерживающей способности однолетних побегов ив (рис.2) показывают, что по содержанию воды, соответствующему порогу летального обезвоживания, ивы распределяются на три группы. Первая группа - ива корзиночная и форма двуцветная ивы трехтычинковой, для которых пороговое содержание воды было 10-15%. Вторая группа - ивы белая, козья, остролистная, форма одноцветная ивы трехтычинковой, для которых пороговое значение обводненности побегов составляет 25-30%. Третья группа - ива серая и шерстистопобеговая, для этих видов пороговое содержание воды и однолетних побегов находилась в пределах 3040%. Отметим, что содержание воды в побегах в кроне растений (контроль) было стабильным с декабря до апреля, затем обводненность увеличивается, что связано с весенней активизацией растений. Следует указать, что пороговые значения содержания воды в побегах ивы белой и ивы серой отмечались в марте, в то время как у ивы остролистной, ивы козьей, формы одноцветной ивы трехтычинковой - в апреле. Однолетние побеги ивы корзиночной и формы двуцветной ивы трехтычинковой обладают большой жизнеспособностью, сильной водоудерживающей способностью и при высушивании достигают порогового содержания воды (10-15%) в мае. Добавим, что начало вегетации изученных видов и форм ив приходится на III декаду апреля - I декаду мая. В целом установлено, что разные виды ив в течение одного и того же периода теряют различное количество воды, что отмечается и для других древесных растений (Оликер, 1965).
Полученные материалы характеризуют особенности водного режима ив и свидетельствуют о значительных межвидовых различиях, что согласуются с устойчивостью отдельных видов ив к загрязнителям (Кулагин, 1998). В целом устойчивость ив к недостатку влаги основана на экологической видос-пецифичности и базируется на морфо-физиологическом комплексе структур и свойств.
I
2 Время подсушивания, ч
А
5 Время подсушивания, ч
* Время подсушивания, ч подсусивскля, ч
S
ВО
Рис. 1. Динамика обезвоживания (1, 2) и ликвидации водного дефицита (1а, 2а) листьев ивы корзиночной (А - 1,1а), ивы шерстистопобеговой (А - 2,2а), ивы серой (В - 1,1а), ивы козьей (В - 2,2а), ивы трехтычинковой формы одноцветной (С - 1,1а), ивы трехтычинковой формы двуцветной (С - 2,2а), ивы белой (D - 1,1а) и ивы остролистной (D - 2,2а).
