Фенотипическая изменчивость Cypripedium calceolus L. (Orchidaceae) на северном пределе распространения
 на северном пределе распространения")
Автор: Кириллова И.А.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 4 (28), 2016 года.
Бесплатный доступ
Проведена оценка фенотипической изменчивости Cypripedium calceolus L. на территории Республики Коми, где вид находится на северной границе своего распространения. Установлено, что фенотипическая пластичность в большей степени связана с признаками вегетативных органов (высота растений, размеры листьев), которые обеспечивают адаптацию растений к различным условиям произрастания. Признаки цветка более стабильны. Выявлена зависимость признаков растений от экологических условий биотопа, погодных условий вегетационного периода и географического положения.
Орхидные, фенотипическая изменчивость, граница ареала
Короткий адрес: https://sciup.org/14992863
IDR: 14992863 | УДК: 582.594:581.16(470.13)
Phenotypic variability of Cypripedium calceolus L. (Orchidaceae) on its northern distribution bound
The phenotypic variability of Cypripedium calceolus L. (Orchidaceae) growing on the northern bound of its distribution in territory of the Komi Republic is described. It is revealed that miniaturization and oligomerization of plants of this species occurs on the northern bound of distribution, compared to specimens growing in the center of the area. The characters of vegetative sphere demonstrate high and medium variation, but flowers morphology is less variable. Weather conditions influence the habitus of plants which is expressed in the size of leaves and bracts and height of shoots. The weather has no effect on number of flowers and leaves. The correlation of characters is increased in unfavourable growing periods. In plants growing in different biotopes the variability of morphometric characteristics caused by ecological differences is observed. Thus plants from forest habitats were larger than specimens growing in swampy areas due to the lack of light in forest phytocenoses. Besides, geographical variation was revealed in studied populations: gradual reduction of plants habitus within latitudinal moving from south to north. The most flexible characters which respond to changing environmental conditions are the height of shoots and size of leaves.
Текст научной статьи Фенотипическая изменчивость Cypripedium calceolus L. (Orchidaceae) на северном пределе распространения
Анализ соответствия между организмом и средой – одна из важнейших проблем современной биологии. Это соответствие проявляется как способность популяций адекватно реагировать на действующие факторы. Поэтому исследование особенностей реализации фенотипического потенциала популяций в различных условиях является важной теоретической задачей на пути понимания механизмов устойчивости биологических систем [1].
Морфологические параметры растений варьируют по ареалу вида. Популяции видов на границе ареала находятся под действием стресса и существуют на пределе своих биологических возможностей, они обладают своеобразной экологической приуроченностью, внутренней организацией, структурой, морфологией и генетикой. Краевые популя- ции часто фрагментарны и более уязвимы к изменениям внешней среды [2]. Многие авторы в краевых популяциях отмечают уменьшение размеров растений [3, 4], изменение размаха изменчивости и скоррелированности признаков [5–7].
В качестве объекта изучения был выбран Cypripedium calceolus L. (башмачок настоящий) ( Orchidaceae ) (см. фото). Этот вид орхидей имеет довольно широкий ареал, но является повсеместно редким (охраняется в России [8] и всех странах Европы [9]). В Республике Коми он находится на северной границе своего распространения. В регионе известно около 100 его местонахождений, вид встречается от самого юга республики до Приполярного Урала. C. calceolus неоднократно становился объектом различных исследований по всему ареалу [9–15 и др.]. Однако работ по краевым популяциям этого вида немного [16–19], и связаны они в ос-

Фото. Башмачок настоящий.
новном с состоянием популяций и репродуктивной биологией. Цель настоящей работы – изучение закономерностей варьирования морфологических параметров C. calceolus на территории Республики Коми.
Материал и методы
Исследования проводили в 2004–2014 гг. на территории Республики Коми. Обследовано 20 це-нопопуляций (ЦП) вида (табл. 1) в разных частях региона: на Вычегодско-Мезенской равнине, Южном и Среднем Тимане, Северном и Приполярном Урале. При изучении морфометрических особенностей растений учитывались высота, число и размеры листьев и цветков. В каждой ЦП проанализировано по 30 побегов генеративного онтогенетического состояния. Данные обработаны вариационностатистическими методами с использованием пакетов STATISTICA 6 и MICROSOFT EXСEL. Для изучаемых морфометрических признаков определяли среднее арифметическое значение (М), ошибку средней (m), коэффициент вариации (СV). Всего проанализировано 840 генеративных побегов. При изучении закономерностей внутривидовой изменчивости использовали методику С.А. Мамаева [20, 21]. Выделяли два типа изменчивости – индивидуальную, которая отражает вариабельность особей в пределах популяций, и межпопуляционную (эколого-географическую), отражающую дифференциацию популяций в пределах ареала вида. Для оценки пластичности использовали индекс фитоценоти-ческой пластичности (I p ) [22]. При сравнении арифметических средних применяли критерий Стьюдента (C St ), достоверность которого оценивали при доверительном интервале 95% [23]. Наряду с одномерной статистикой применяли многомерные ста-
Таблица 1
Характеристика изученных ценопопуляций Cypripedium calceolus в Республике Коми
|
Год изучения |
Местонахождение |
Местообитание, фитоценоз |
|
2011, 2014 |
ВМР, пойма р. Сысола |
Сосново-березовый хвощово-осоковый лес |
|
2010, 2011 |
ВМР, пойма р. Сысола |
Ельник разнотравный |
|
2010, 2011 |
ВМР, пойма р. Важъелью |
Осоково-сфагновое болото |
|
2014 |
ВМР, пойма р. Важъелью |
Хвощово-осоково-сфагновое болото |
|
2014 |
ВМР, пойма р. Важъелью |
Ельник разнотравно-сфагновый |
|
2009, 2010, 2011 |
ВМР, пойма р. Дырнос |
Елово-березовый разнотравный лес |
|
2014 |
ВМР, окр. дер. Занулье |
Ельник разнотравно-чернично-зеленомошный |
|
2011 |
ЮТ, левый берег р. Сойва |
Еловое редколесье разнотравно-зеленомошное на склоне юго-западной экспозиции (выходы известняков) |
|
2009, 2011 |
ЮТ, левый берег р. Омра |
Сосновое редколесье разнотравно-осоково-зеленомошное на склоне южной экспозиции (выходы известняков) |
|
2009 |
СТ, левый берег р. Мыла |
Верхняя часть скалистого склона юго-восточной экспозиции, сосняк лишайниковый |
|
2004 |
СУ, левый берег р. Печора |
Ельник разнотравно-сфагновый |
|
2007 |
СУ, левый берег р. Илыч |
Каменистый облесенный склон у края осиново-березового (с елью и пихтой) высокотравного леса |
|
2007 |
СУ, левый берег р. Илыч |
Скалистая осыпь, облесенная березой и елью, разнотравно-зеленомошная |
|
2008 |
СУ, правый берег р. Ыджид-Сотчемъель |
Облесенный склон южной экспозиции, скальные растительные группировки с преобладанием Rubus saxatlis и Antennaria dioica |
|
2010, 2011 |
ПУ, правый берег р. Кожим |
Облесенный лиственницей склон, кустарничково-моховые сообщества |
|
2010, 2011 |
ПУ, левый берег р. Кожим |
Днище отстойника, бывший полигон золотодобычи |
|
2011 |
ПУ, левый берег р. Кожим |
Скалы, кустарничково-лишайниковые сообщества |
|
2010, 2011 |
ПУ, правый берег р. Кожим |
Плато на вершине скал, разнотравный ивняк |
|
2010, 2011 |
ПУ, правый берег р. Кожим |
Плато на вершине скал, пихтово-еловый травяной лес |
|
2011 |
ПУ, правый берег р. Кожим |
Край елово-лиственничного кустарничково-зеленомошного леса |
Примечание: ВМР – Вычегодско-Мезенская равнина; ЮТ – Южный Тиман; СТ – Средний Тиман; СУ – Северный Урал; ПУ – Приполярный Урал.
тистические методы – дискриминантный и кластерный анализ. Кластеризацию осуществляли на основании полученных в дискриминантном анализе квадратов расстояния Махалонобиса. Для выявления связей между морфологическими признаками вегетативных и репродуктивных органов использовали корреляционный анализ. Рассчитывали индекс морфологической интеграции особей – I [24], как отношение числа статистически существенных связей в корреляционной матрице к их общему числу.
Результаты и обсуждение
Индивидуальная изменчивость. Под индивидуальной изменчивостью понимается изменчивость особей в пределах одной популяции [20]. С. сalceolus рассмотрен нами в рамках региональной популяции. В Республике Коми высота генеративных побегов этого вида составляет в среднем 26 (8–53) см. На один побег приходится (2) 3,1 (5) листьев, (4) 9–12 (21) см длиной и (2) 5–7 (15) см шириной. Цветки одиночные, реже в числе двух. Прицветники (брактеи) 7 (2–14) см длиной и 3 (1–7) см шириной. Верхний листочек околоцветника 5 (2–8) см длиной и 2 (1,2–3,3) см шириной. Синсепалум около 5 (2,5–7) см длиной и 2 (0,6–2,6) см шириной. Боковые листочки околоцветника 6 (3,2–8,5) см длиной и 0,6 (0,3–0,9) см шириной. Губа 3,4 (1,5– 5,5) см длиной, 2,4 (1,2–4) см шириной и 1,9 (1,0– 3,5) см высотой. У двуцветковых особей нижний цветок несколько крупнее верхнего, у него достоверно крупнее верхний листочек околоцветника, а также длины боковых листочков околоцветника, размеры губы не меняются. Плод – коробочка, (1,2) 2–3 (4,5) см длиной и 0,5–1 см шириной.
Размеры растений C. calceolus в регионе в целом не выходят за пределы нормы реакции для этого вида, но по сравнению с данными из других частей ареала [25, 12, 14, 3] сдвинуты к нижней границе, т.е. на северном пределе распространения вида происходит уменьшение размеров растений и количества метамеров, что свойственно и другим травянистым растениям в экстремальных условиях существования [4].
Признаки C. calceolus варьируют на высоком и среднем уровнях, а ширина прицветника – на очень высоком. Наиболее изменчивы ширина прицветника (CV=41,1%), высота побега (35,3%) и ширина нижнего листа (35,7%). Данные признаки можно отнести к эколого-биологическим индикаторам [7], которые отражают реакцию организма на условия среды. Наименее вариабельны число листьев и размеры частей цветка (кроме ширины бокового листочка околоцветника). Эти признаки можно отнести к таксоноспецифичным. В целом признаки генеративной сферы менее изменчивы, чем признаки вегетативной. На самом низком уровне варьируют длина и высота губы (CV=14,5% и 14,9% соответственно).
Рассмотрена также индивидуальная изменчивость всех изученных ЦП вида . Полученные результаты приведены в табл. 2. Признаки могут варьировать от очень низкого до очень высокого уровня, причем колебания коэффициентов вариации в разных ЦП у одинаковых признаков могут быть значительными. Наиболее вариабельным признаком является ширина прицветника, коэффициент вариации этого признака составляет в разных ЦП от 20,3 до 48,6%. Такие признаки, как высо-
Таблица 2
Индивидуальная изменчивость морфометрических признаков Cypripedium calceolus
|
Признак |
Средние значения в ЦП (min-max) |
Коэффициент вариации в ЦП (min-max) |
I p |
Средний коэффициент вариации |
|
ВП, см |
13,4-40,5 |
10,3-33,2 |
0,67 |
17,5 |
|
ЧЛ, шт. |
2,6-4,0 |
8,7-24,8 |
0,35 |
15,0 |
|
Д1Л, см |
5,9-14,9 |
7,3-37,9 |
0,60 |
18,8 |
|
Ш1Л, см |
3,3-9,9 |
6,6-36,0 |
0,67 |
20,6 |
|
Д2Л, см |
7,5-17,3 |
6,1-23,1 |
0,57 |
11,5 |
|
Ш2Л, см |
4,0-10,8 |
11,2-27,1 |
0,63 |
17,1 |
|
ДПр, см |
4,3-9,7 |
13,0-38,0 |
0,56 |
21,0 |
|
ШПр, см |
1,9-4,8 |
20,3-48,6 |
0,60 |
33,2 |
|
ДГ, см |
2,4-3,9 |
7,0-17,6 |
0,38 |
9,7 |
|
ШГ, см |
1,6-2,9 |
7,0-21,2 |
0,45 |
11,1 |
|
ВГ, см |
1,6-2,2 |
8,1-15,9 |
0,27 |
10,8 |
|
ДВЛ, см |
3,6-6,3 |
6,9-17,6 |
0,43 |
11,3 |
|
ШВЛ, см |
1,4-2,6 |
4,0-18,1 |
0,46 |
11,4 |
|
ДБЛ, см |
4,0-7,0 |
8,7-15,6 |
0,43 |
11,6 |
|
ШБЛ, см |
0,5-0,7 |
3,0-27,3 |
0,29 |
17,8 |
|
ДНЛ, см |
3,6-5,9 |
8,5-21,2 |
0,39 |
12,7 |
|
ШНЛ, см |
1,1-2,1 |
8,2-19,1 |
0,47 |
13,5 |
|
ЧЦ, шт. |
1,0-1,5 |
18,2-35,5 |
0,33 |
31,4 |
Примечание: здесь и далее в табл. 4, 5: ВП – высота побега, ЧЛ – число листьев, Д1Л – длина первого (нижнего) листа, Ш1Л – ширина первого (нижнего) листа, Д2Л – длина второго листа, Ш2Л – ширина второго листа, ДПр – длина прицветника, ШПр – ширина прицветника, ДГ – длина губы, ШГ – ширина губы, ВГ – высота губы, ДВЛ – длина верхнего листочка околоцветника, ШВЛ – ширина верхнего листочка околоцветника, ДБЛ – длина бокового листочка околоцветника, ШБЛ – ширина бокового листочка околоцветника, ДНЛ – длина нижнего листочка околоцветника, ШНЛ – ширина нижнего листочка околоцветника, ЧЦ – число цветков.
та побега, число листьев и ширина второго листа демонстрируют в основном средний уровень изменчивости, длина прицветника – высокий. Размеры нижнего листа в половине ЦП показали средний уровень изменчивости, в остальных – высокий. Длина второго листа варьирует в ЦП на среднем и низком уровнях. Признаки цветка более стабильны, чем признаки вегетативной сферы. Самый мало изменчивый признак – длина губы, демонстрирует в основном низкий уровень изменчивости. Размеры листочков околоцветника варьируют на среднем, реже – низком уровнях. При этом их ширина – более вариабельный признак, чем длина. Самый вариабельный признак генеративной сферы – ширина бокового листочка околоцветника, в половине ЦП он варьирует на высоком уровне, в остальных – на среднем. Подобные закономерности изменчивости признаков генеративных побегов C. calceolus выявлены и на Южном Урале [26].
Наиболее пластичными параметрами, максимально реагирующими на изменения среды обитания, оказались высота побега, размеры нижнего листа, ширина второго листа и прицветника (I p = 0,60-0,67) (табл. 2). Остальные параметры менее пластичны. Стабильными параметрами с минимальным уровнем пластичности являются высота губы, ширина бокового листочка околоцветника, число цветков и листьев.
Влияние погодных условий на размер растений было прослежено на примере 9 ЦП C. cal-ceolus . На Приполярном Урале 4 ЦП данного вида изучали в 2010 и 2011 гг. Вегетационные периоды этих лет были довольно прохладными, а 2011 г. еще и сухими (сумма осадков была ниже среднемноголетнего значения почти в два раза) (табл. 3). Признаки в ЦП оставались стабильными, за исключением размеров второго листа, длина которого несколько увеличилась в трех из обследованных ЦП в 2011 г. Выросла также скоррелированность признаков в ЦП.
На Южном Тимане ЦП C. calceolus обследована в 2009 и 2011 гг. Первый год изучения был прохладный и сухой, второй – теплый и достаточно влажный. В 2009 г. была достоверно больше высота побега, длина листьев и прицветников, а также ниже скоррелированность признаков (I=41,7%, вместо 69,4% – в 2011 г.).
На Вычегодско-Мезенской равнине исследования влияния погодных условий на габитус растений проводили в 2009–2014 гг. Вегетационный период 2009 г. был прохладным и влажным, 2010 г. – умеренно теплым и относительно сухим, вегетационный период 2011 г. отличался жаркой и сухой погодой (сумма активных температур (>10° С) оказалась выше среднемноголетнего значения за последние пять лет), 2014 г. был умеренно теплым и влажным. Растения, изученные на болоте, в 2010 и 2011 гг. практически не отличались, лишь выросла скоррелированность признаков в 2011 г. с 30,5 до 44,4%. В двух лесных ЦП (ЦП 2 и 6) в 2011 г., по сравнению с предыдущим, уменьшилась высота побегов, размеры листьев, прицветников и цветка и увеличилась скоррелированность признаков. В ЦП6 в 2009 г. была достоверно меньше длина нижнего листа, ширина прицветника, высота губы и длина нижнего листочка околоцветника, а также скорре-лированность признаков, чем в 2010 г. ЦП 1 обследована в 2011 и 2014 гг., в первый год изучения растения были достоверно мельче.
Больше всего погодные условия влияют на размеры листьев и прицветников. На ряд признаков (число листьев, цветков) условия вегетационного периода воздействия не оказывают. С теплообес-печенностью коррелируют в основном признаки вегетативной сферы, в более холодные вегетационные периоды размеры листьев, а в некоторых ЦП и высота побегов увеличиваются. В более суровых условиях (на Приполярном Урале) растения меньше зависят от погодных условий, в них повышается выровненность признаков. Обнаружена связь количества осадков со скоррелированностью признаков, в вегетационные периоды с более низким количеством осадков скоррелированность признаков выше. Н.С. Ростовой [6, 7] на примере многих травянистых растений было установлено, что неблагоприятные условия среды вызывают значительное повышение силы связей между признаками. Это явление указывает на хорошую приспособляемость растений к неблагоприятным условиям (недостатку влаги).
Таблица 3
Метеоусловия вегетационных периодов 2009–2014 гг. в разных частях Республики Коми
|
Параметр |
2009 г. |
2010 г. |
2011 г. |
2014 г. |
|
Вычегодско-Мезенская равнина |
||||
|
Сумма активных температур (>10° С) |
1260,5 |
1388,8 |
1414,4 |
1284,5 |
|
Сумма осадков за период с температурой >10° С |
239,2 |
146,1 |
122,1 |
247,4 |
|
Гидротермический коэффициент Селянинова |
1,9 |
1,1 |
0,9 |
1,9 |
|
Южный Тиман |
||||
|
Сумма активных температур (>10° С) |
1064,4 |
1211,6 |
1137,3 |
1121,0 |
|
Сумма осадков за период с температурой >10° С |
138,2 |
127,6 |
155,2 |
201,2 |
|
Гидротермический коэффициент Селянинова |
1,3 |
1,1 |
1,4 |
1,8 |
|
Приполярный Урал |
||||
|
Сумма активных температур (>10° С) |
785,3 |
665,6 |
647,7 |
782,0 |
|
Сумма осадков за период с температурой >10° С |
99,2 |
128,0 |
71,5 |
186,5 |
|
Гидротермический коэффициент Селянинова |
1,3 |
1,9 |
1,1 |
2,4 |
Экологическая изменчивость . С целью изучения экологической изменчивости рассматривали ЦП C. calceolus , произрастающие в разных экологических условиях в пределах одного орографического выдела. В южной части региона, на Вычегодско-Мезенской равнине, вид встречается на притеррасных болотах в долинах рек, облесенных окраинах болот, приручейных или заболоченных лесах. Кластерный анализ, проведенный для 7 ЦП с Вычегодско-Мезенской равнины с использованием всех признаков вегетативной и генеративной сферы, показал деление на две группы – болотную (ЦП 3, 4) и лесную (ЦП 1, 2, 5, 6), отдельно вышла ЦП с самого юга Вычегодско-Мезенской равнины (ЦП 7) (рис. 1). Лесная группа разбилась еще на две – еловых лесов (ЦП 1) и сосняков (ЦП 2, 5, 6). Дискриминантный анализ показал, что в разделение групп наибольший вклад внесли такие признаки, как ширина листьев, длина верхнего листочка околоцветника (по первой дискриминантной оси), высота побегов и число листьев (по второй).
ния поверхности улавливания света. В разных условиях произрастания морфологические признаки варьируют сходным образом, меняется лишь размах варьирования, на болоте признаки более изменчивы, чем в лесу (табл. 4).
Географическая изменчивость отражает пространственно закрепленное влияние большого ряда экологических факторов: климат, макрорельеф и т.д. Характерной ее чертой является та или иная степень пространственной обособленности, что обеспечивает и генетическую изоляцию [20]. В рамках изучения географической изменчивости нами проводилось рассмотрение клинальной изменчивости признаков по региону. С юга на север при ухудшении условий для произрастания растений происходит постепенное снижение габитуса (например, высота побегов изменяется от 40,5 до 16 см) и числа листьев (табл. 5). Также происходит уменьшение размеров цветка. На Приполярном Урале длина листочков околоцветника в 1,7 раза меньше, чем на юге региона. Число цветков значи-
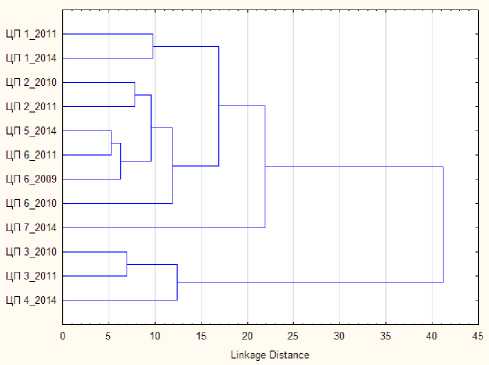
Рис. 1. Распределение ценопопуляций Cypripedium calceolus с Вычегодско-Мезенской равнины при проведении кластерного анализа (нумерация ЦП, та же, что и в табл. 1).
a юг РК □ ВМР ^ ЮТ в СУ □ ПУ
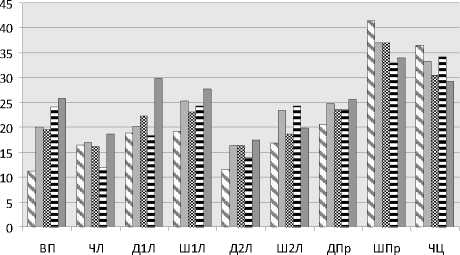
Рис. 2. Изменчивость морфометрических признаков Cypripedium calceolus , растущих в разных частях Республики Коми (юг РК – юг Республики Коми; ВМР – Вычегодско-Мезенская равнина; ЮТ – Южный Тиман; СУ – Северный Урал; ПУ – Приполярный Урал) (по оси абсцисс – признаки (обозначения даны в табл. 2); по оси ординат – коэффициент вариации, %).
Наблюдается сильное различие по многим морфометрическим признакам растений болотных и лесных местообитаний (табл. 4). При этом происходит достоверное уменьшение практически всех признаков вегетативных органов и цветка у растений болотных ассоциаций, число цветков не меняется, а число листьев больше у болотных растений. Растения, произрастающие в сосняках и ельниках, также отличаются между собой. Генеративные особи C. calceolus , произрастающие в еловых лесах, достоверно выше, с более широкими листьями и прицветниками, с более короткими, но широкими листочками околоцветника.
Все эти изменения связаны главным образом с разными условиями освещения. Уменьшение количества света ведет к сокращению числа листьев, но увеличению их размеров. То есть, адаптивным ответом на слабое освещение является увеличение площади листьев для максимального увеличе- мо не отличается, в любой части региона в ЦП вида отмечен примерно одинаковый процент растений с двумя цветками.
Признаки варьируют в разных частях региона сходным образом. При продвижении на север при уменьшении размеров растений происходит увеличение вариабельности большинства признаков (рис. 2). Возможно, это помогает отдельным особям в стрессовых условиях оптимально использовать ресурсы среды.
Сравнивались также популяции, растущие в сходных условиях (выходы известняков) из разных частей региона (рис. 3). Кластерный анализ, проведенный для 11 ЦП, показал деление на два кластера, в первый вошли ЦП с Тимана и Северного Урала (ЦП 8-10, 12-14), во второй – с Приполярного Урала (ЦП 15-18, 20). Первый кластер при этом разделился на два – в первый вошли ЦП с Южного и Среднего Тимана (ЦП 8-10), во второй – с Север-
Таблица 4
Сравнение морфометрических признаков у групп ценопопуляций Cypripedium calceolus, растущих в различных экологических условиях
|
Признак |
Болото |
Лес |
C st |
Сосняки |
Ельники |
C st |
|
M±m (min-max) CV |
M±m (min-max) CV |
M±m (min-max) |
M±m (min-max) |
|||
|
ВП, см |
26,2±0,47 (18,0-36,4) 17,0 |
35,3±0,36 (22,0-53,0) 15,3 |
15,4* |
34,1±0,54 (16,5-26,5) |
35,8±0,44 (22,0-53,0) |
2,4* |
|
ЧЛ, шт. |
3,5±0,07 (2-5) 19,1 |
3,3±0,03 (2-5) 15,9 |
2,3* |
3,3±0,06 (3-4) |
3,3±0,04 (2-5) |
0,1 |
|
Д1Л, см |
9,4±0,17 (5,5-12,0) 16,9 |
11,8±0,14 (6,5-16,5) 17,8 |
11,2* |
11,7±0,28 (7,0-15,8) |
11,9±0,16 (6,5-16,5) |
0,6 |
|
Ш1Л, см |
5,3±0,13 (2,9-8,5) 24,1 |
7,3±0,10 (2,8-12,0) 21,0 |
11,9* |
6,5±0,18 (2,8-9,2) |
7,5±0,11 (4,4-12,0) |
4,8* |
|
Д2Л, см |
10,8±0,12 (8,2-14,2) 10,8 |
14,4±0,10 (11,0-18,0) 10,3 |
22,5* |
14,7±0,17 (11,4-17,5) |
14,3±0,12 (11,0-18,0) |
1,9 |
|
Ш2Л, см |
5,6±0,13 (3,2-10,0) 23,0 |
7,9±0,09 (4,8-12,5) 17,1 |
14,6* |
7,3±0,13 (4,8-9,5) |
8,2±0,11 (5,2-12,5) |
5,4* |
|
ДПр, см |
6,3±0,15 (2,4-10,5) 24,5 |
8,8±0,12 (4,0-14,0) 20,6 |
13,3* |
8,8±0,23 (5,5-13,5) |
8,8±0,13 (4,0-14,0) |
0,1 |
|
ШПр, см |
2,5±0,09 (0,8-4,7) 37,1 |
3,5±0,06 (0,8-7,3) 34,7 |
8,6* |
3,0±0,13 (0,8-5,7) |
3,7±0,09 (0,8-7,3) |
4,3* |
|
ДГ, см |
3,2±0,03 (1,5-4,5) 11,0 |
3,6±0,03 (2,7-7,0) 11,3 |
9,1* |
3,6±0,04 (2,8-4,2) |
3,6±0,03 (2,7-7,0) |
1,4 |
|
ШГ, см |
2,3±0,04 (1,6-4,0) 16,0 |
2,6±0,02 (1,8-3,4) 11,7 |
7,6* |
2,6±0,03 (1,9-3,2) |
2,7±0,02 (1,8-3,4) |
1,5 |
|
ВГ, см |
1,8±0,03 (1,3-3,2) 16,7 |
2,0±0,02 (1,4-3,5) 14,0 |
4,8* |
2,0±0,03 (1,5-2,5) |
1,9±0,02 (1,4-3,5) |
2,9* |
|
ДВЛ, см |
4,9±0,05 (3,5-6,0) 11,1 |
5,7±0,05 (2,0-8,0) 13,6 |
11,2* |
6,1±0,10 (4,5-8,0) |
5,6±0,05 (2,0-5,7) |
4,1* |
|
ШВЛ, см |
2,2±0,03 (1,4-3,0) 14,7 |
2,2±0,02 (1,4-3,3) 14,6 |
1,7 |
2,1±0,03 (1,4-2,6) |
2,3±0,02 (1,4-3,3) |
4,0* |
|
ДБЛ, см |
5,3±0,07 (3,6-7,5) 12,7 |
6,3±0,05 (4,0-8,5) 13,1 |
12,2* |
6,7±0,11 (4,5-8,5) |
6,2±0,06 (4,0-8,2) |
3,9* |
|
ШБЛ, см |
0,6±0,01 (0,4-0,8) 20,8 |
0,6±0,01 (0,4-0,9) 20,3 |
0,4 |
0,5±0,01 (0,4-0,8) |
0,6±0,01 (0,4-0,9) |
4,1* |
|
ДНЛ, см |
4,6±0,05 (3,4-5,8) 11,4 |
5,4±0,05 (3,3-7,4) 13,4 |
11,2* |
5,7±0,09 (4,0-7,4) |
5,3±0,05 (3,3-7,4) |
4,0* |
|
ШНЛ, см |
1,7±0,03 (0,8-2,6) 17,7 |
1,8±0,02 (1,2-2,5) 15,3 |
3,0* |
1,7±0,03 (1,2-2,2) |
1,9±0,02 (1,2-2,5) |
4,9* |
|
ЧЦ, шт. |
1,2±0,04 (1-2) 34,7 |
1,2±0,02 (1-2) 32,6 |
1,2 |
1,2±0,06 (1-2) |
1,2±0,03 (1-2) |
1,5 |
Таблица 5
Морфометрические параметры генеративных особей Cypripedium calceolus в разных частях Республики Коми
|
Признак |
Юг Республики Коми1 |
Вычегодско-Мезенская равнина |
C st |
Южный Тиман3 |
C st |
Северный Урал4 |
C st |
Приполярный Урал5 |
C st |
|
ВП, см |
40,5±0,86 (32,0-51,5) |
33,0±0,36 (18,0-53,0) |
8,0* |
24,8±0,52 (15,0-40,0) |
13,0* |
23,8±0,62 (12,0-44,0) |
1,2 |
16,0±0,26 (8,0-29,2) |
11,6* |
|
ЧЛ, шт. |
3,8±0,12 (2-5) |
3,4±0,03 (2-5) |
3,6* |
3,1±0,05 (2-4) |
4,2* |
3,0±0,04 (2-4) |
1,0 |
2,8±0,03 (2-4) |
3,5* |
|
Д1Л, см |
14,9±0,53 (9,8-20,7) |
11,2±0,12 (5,5-16,5) |
6,8* |
8,8±0,21 (4,4-14,0) |
10,1* |
9,9±0,20 (6,6-14,6) |
4,1* |
6,9±0,12 (3,0-13,5) |
13,0* |
|
Ш1Л, см |
9,9±0,36 (6,0-13,3) |
6,7±0,09 (2,8-12,0) |
8,5* |
4,6±0,11 (2,2-7,5) |
14,2* |
5,2±0,14 (2,8-8,6) |
3,1* |
4,1±0,07 (1,7-6,8) |
7,0* |
|
Д2Л, см |
17,3±0,37 (14,0-21,5) |
13,5±0,12 (8,3-18,5) |
9,5* |
11,1±0,20 (7,5-16,4) |
10,7* |
12,1±0,18 (9,2-16,6) |
3,7* |
8,7±0,09(5, 0-14,7) |
16,3* |
|
Ш2Л, см |
10,8±0,34 (8,2-15,0) |
7,3±0,09 (3,2-12,5) |
9,9* |
5,5±0,11 (3,8-8,0) |
12,2* |
6,2±0,16 (3,2-10,0) |
3,3* |
4,6±0,06 (2,2-6,6) |
9,2* |
|
ДПр, см |
9,7±0,35 (5,3-13,0) |
8,1±0,11 (3,0-14,0) |
4,1* |
6,3±0,15 (3,5-10,4) |
10,1* |
7,5±0,18 (4,0-12,0) |
5,1* |
5,1±0,08 (2,0-9,5) |
11,3* |
|
ШПр, см |
4,8±0,47 (1,4-7,7) |
3,3±0,06 (0,8-7,3) |
3,3* |
2,6±0,10 (0,5-6,2) |
5,2* |
3,5±0,13 (1,7-6,8) |
5,5* |
2,2±0,05 (0,5-4,6) |
9,7* |
|
ДГ, см |
3,8±0,07 (3,3-4,4) |
3,5±0,02 (1,5-4,6) |
4,4* |
3,0±0,05 (2,0-3,7) |
8,8* |
- |
2,5±0,06 (1,6-3,3) |
6,0* |
|
|
ШГ, см |
2,5±0,05 (2,2-2,8) |
2,5±0,02 (1,6-4,0) |
0,3 |
1,9±0,04 (1,3-2,4) |
13,2* |
- |
1,6±0,03 (1,2-1,8) |
6,3* |
|
|
ВГ, см |
2,0±0,05 (1,8-2,5) |
1,9±0,01 (1,3-3,5) |
1,7 |
1,8±0,03 (1,4-2,2) |
2,6* |
- |
1,5±0,04 (1,0-2,0) |
6,2* |
|
|
ДВЛ, см |
6,2±0,15 (5,5-7,5) |
5,5±0,04 (2,0-8,0) |
4,6* |
4,6±0,09 (3,3-5,6) |
8,9* |
- |
3,6±0,09 (3,0-4,6) |
7,8* |
|
|
ШВЛ, см |
2,3±0,06 (1,8-2,8) |
2,2±0,02 (1,4-3,3) |
1,8 |
1,7±0,03 (1,2-2,2) |
12,7* |
- |
1,4±0,02 (1,2-1,7) |
6,9* |
|
|
ДБЛ, см |
6,8±0,19 (5,6-8,3) |
6,0±0,05 (3,6-8,5) |
3,9* |
5,2±0,10 (3,6-7,0) |
6,9* |
- |
4,0±0,12 (3,2-5,5) |
7,5* |
|
|
ШБЛ, см |
0,7±0,03 (0,5-0,9) |
0,6±0,01 (0,4-0,9) |
3,4* |
0,5±0,01 (0,3-0,8) |
8,2* |
- |
0,5±0,03 (0,3-0,7) |
1,3 |
|
|
ДНЛ, см |
5,8±0,02 (4,8-7,3) |
5,2±0,04 (3,2-7,4) |
3,5* |
4,4±0,10 (3-5,5) |
7,4* |
- |
3,6±0,10 (2,5-4,6) |
5,1* |
|
|
ШНЛ, см |
2,0±0,06 (1,4-2,3) |
1,8±0,02 (0,8-2,6) |
2,9* |
1,4±0,04 (0,6-1,9) |
11,1* |
- |
1,3±0,03 (1,2-1,6) |
0,6 |
|
|
ЧЦ, шт. |
1,3±0,06 (1-2) |
1,2±0,02 (1-2) |
0,7 |
1,1±0,04 (1-2) |
1,3 |
1,2±0,04 (1-2) |
1,3 |
1,1±0,02 (1-2) |
1,9 |
Примечание: 1 – выборка 30 растений; 2 – выборка 360 растений; 3 – выборка 90 растений; 4 – выборка 100 растений; 5 – выборка 255 растений. В таблице приведены среднее значение, стандартная ошибка, минимальное и максимальное значения признаков и критерий Стьюдента для попарных сравнений признаков (СSt); звездочкой отмечены значения критерия Стьюдента, достоверные при 95%-ном доверительном интервале.
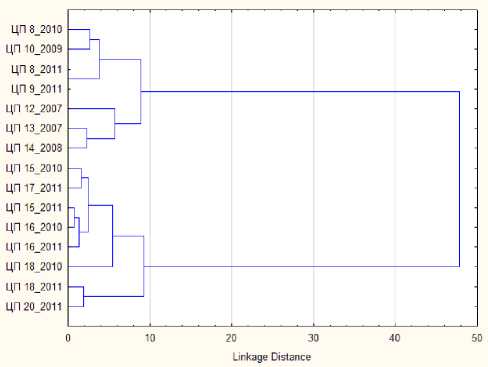
Рис. 3. Распределение скальных ценопопуляций Cypripedium calceolus при проведении кластерного анализа (нумерация ЦП, та же, что и в табл. 1).
ного Урала (ЦП 12-14). В разделение ЦП наибольший вклад по первой дискриминантной оси внесли такие признаки, как размер нижнего листа и длина прицветника, по второй – длина второго листа и высота побегов.
Межпопуляционная изменчивость не ограничивается клинальной изменчивостью. Результаты ее оценки представлены на рис. 4, где приводятся коэффициенты вариации признаков при межпопуляционной и индивидуальной изменчивости. Отмечен большой размах межпопуляционной изменчивости, что связано с довольно широким распространением вида в регионе и произрастанием в различных биотопах. Признаки вегетативной сферы варьируют в основном на высоком уровне (кроме числа листьев), показатели частей цветка – на среднем (ширина губы, размеры листочков околоцветника) и низком (длина и высота губы, ширина бокового листочка околоцветника), число цветков – на низком уровнях.
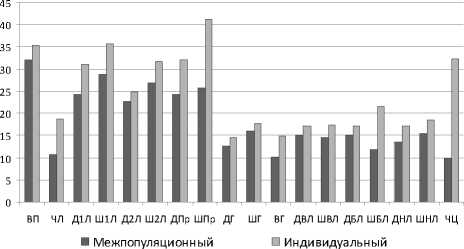
Рис. 4. Варьирование морфометрических признаков Cypripedium calceolus на межпопуляционном и индивидуальных уровнях (по оси абсцисс – признаки (обозначения даны в табл. 2); по оси ординат – коэффициент вариации, %).
Уровень межпопуляционной изменчивости в целом ниже уровня индивидуальной изменчивости (рис. 4), однако для некоторых признаков схож. Так, примерно на том же уровне изменчивости остаются длина второго листа, длина и ширина губы. Для остальных признаков уровень межпопуляционной изменчивости понижается по сравнению с уровнем индивидуальной изменчивости.
Заключение
Изучение фенотипической изменчивости Cy-pripedium calceolus L. на территории Республики Коми показало, что на северной границе ареала происходит миниатюризация и олигомеризация растений этого вида по сравнению с особями, произрастающими в центре ареала. Признаки вегетативной сферы варьируют на высоком и среднем уровнях, признаки цветка менее изменчивы. Погодные условия оказывают влияние на габитус растений. Больше всего их влиянию подвержены размеры листьев, прицветников и высота побегов. На число цветков и листьев они не оказывают влияния. В неблагоприятные вегетационные периоды увеличивается скоррелированность признаков. У растений, произрастающих в разных биотопах, наблюдается изменчивость морфометрических признаков, обусловленная различиями в экологических условиях. Так, растения из лесных местообитаний оказались крупнее особей, растущих на болотах, что связано, прежде всего, с недостатком света в лесных фитоценозах. Выявлена географическая изменчивость: с юга на север региона происходит постепенное уменьшение габитуса растений. Самыми пластичными признаками, которые реагируют на изменение условий среды, оказались высота побегов и размеры листьев.
Работа выполнена частично при поддержке проекта № 15-12-4-1 «Разнообразие растительного мира и почвенного покрова ландшафтов, перспективных для включения в состав объекта Всемирного наследия ЮНЕСКО «Девственные леса Коми». Программа Президиума РАН «Живая природа: современное состояние и проблемы развития».
Список литературы Фенотипическая изменчивость Cypripedium calceolus L. (Orchidaceae) на северном пределе распространения
- Васильев А.Г., Фалеев В.И., Галактионов Ю.К. и др. Реализация морфологического разнообразия в природных популяциях млекопитающих. Новосибирск: Издательство СО РАН, 2004. 232 с
- Kawecki T.J. Adaptation to marginal habitats//Annual Review of Ecology, Evolution and Systematics. 2008. Vol. 39. Р. 321-342
- Блинова И.В. Особенности географической изменчивости ряда наземных европейских орхидных//Экология. 2012. Т. 34. № 2. С. 106-111
- Мазуренко М.Т. Биоморфологические адаптации растений к условиям Крайнего Севера. М.: Наука, 1986. 209 с
- Годин В.Н. Структура и изменчивость системы связей морфологических признаков цветка Pentaphylloides fruticosa (Rosaceae) в различных условиях горного Алтая//Сибирский экологический журнал. 2004. № 6. С. 885-892
- Ростова Н.С. Корреляционный анализ в популяционных исследованиях//Экология популяций. М., 1991. С. 69-86
- Ростова Н.С. Корреляции: структура и изменчивость. СПб.: Изд-во СПб. ун-та, 2002. 308 с
- Абрамова Т.И., Аверкова Г.П., Аверьянов Л.В. и др. Красная книга Российской Федерации (растения и грибы). М.: Товарищество научных изданий КМК, 2008. 855 с
- Kull T. Cypripedium calceolus L.//Biological flora of the British Isles. J. Ecol. 1999. Vol. 87. P. 913-924
- Фардеева М.Б. Онтогенез башмачка настоящего, или Венерина башмачка (Cypripedium calceolus L.)//Онтогенетический атлас лекарственных растений. Т. III. Йошкар-Ола, 2002. С. 134-138
- Brzosko E. Dynamics of island populations of Cypripedium calceolus in the Biebrza river valley (northeast Poland)//Bot. J. Linn. Soc., 2002. Vol. 139. Р. 67-77
- Состояние ценопопуляций некоторых видов сем. Orchidaceae на Южном Урале. Сообщение 2. Корневищные виды/М.М.Ишмуратова, И.В.Суюндуков, А.Р.Ишбирдин, Т.В.Жирнова, М.И. Набиуллин//Растительные ресурсы. 2003. Т. 39. Вып. 2. С. 18-37
- Михальчук Н.В. К вопросу о выделении онтогенетических групп в ценопопуляциях Cypripedium calceolus L.//Биологический вестник. 2003. Т. 7. № 12. С. 46-48
- Vakhrameeva M.G., Tatarenko I.V., Varlygina T.I. et al. Orchids of Russia and adjacent countries (within the borders of the former USSR). Germany, 2008. 690
- Antonelli A., Dahlberg C.J., Carlgren K.H.I., Appelqvist T. Pollination of the Lady’s slipper orchid (Cypripedium calceolus) in Scandinavia -taxonomic and conservational aspects//Nordic Journal of Botany, 2009. 27. Р. 266-273
- Пучнина Л.В. Состояние ценопопуляций Cypripedium calceolus L. (Orchidaceae) в карстовых ландшафтах Севера европейской России//Ботан. журн., 1999. Т. 84, № 9. С. 75-81
- Blinova I.V. A northernmost population of Cypripedium calceolus L. (Orchidaceae): demography, flowering, pollination//Selbyana, 2002. Vol. 23. № 1. P. 113-120
- Башмачок настоящий/В.А.Мартыненко, И.И.Полетаева, Б.Ю.Тетерюк, Л.В.Тетерюк//Биология и экология редких растений Республики Коми. Екатеринбург, 2003. С. 28-42
- Кириллова И.А., Тетерюк Л.В., Пестов С.В., Кириллов Д.В. Репродуктивная биология Cypripedium calceolus L. (Orchidaceae) на европейском Северо-Востоке России//Ботанический журнал. 2012. Т. 97. № 12. С. 1516-1532
- Мамаев С.А. Формы внутривидовой изменчивости древесных растений. М., 1972. 284 с
- Мамаев С.А. Основные принципы методики исследования внутривидовой изменчивости растений//Индивидуальная и эколого-географическая изменчивость растений. Свердловск, 1975. С. 3-14
- Злобин Ю.А., Скляр В.Г., Клименко А.А. Популяции редких видов растений: теоретические основы и методика изучения. Сумы, 2013. 439 с
- Зайцев Г.Н. Математическая статистика в экспериментальной ботанике. М., 1984. 424 с
- Злобин Ю.А. Принципы и методы изучения ценотических популяций растений. Казань, 1989. 146 с
- Delforge P. Orchids of Britain and Europe. London, 1995. 480 p
- Суюндуков И.В. Стратегии жизни некоторых видов сем. Orchidaceae (Juss.) и вопросы охраны орхидей на Южном Урале: Автореф. дис… доктора биол. наук. Уфа, 2014. 43 с
