Филогенетическое разнообразие нитрилутилизирующих бактерий содового шламохранилища и их некоторые физиолого-биохимические особенности

Автор: Сыровацкая Г.А., Максимов А.Ю., Максимова Ю.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 3, 2025 года.
Бесплатный доступ
Исследовано филогенетическое разнообразие нитрилутилизирующих бактерий содового шламохранилища. Галоалкалотолерантные изоляты, полученные при использовании ацетамида и ацетонитрила в качестве единственных источников углерода, идентифицированы методом секвенирования 16S рДНК. Определено, что изоляты принадлежат к филумам Actinomycetota (родам Arthrobacter, Microbacterium, Rhodococcus) и Pseudomonadota (родам Acinetobacter, Ensifer, Pseudomonas, Rhizobium). Установлено, что изоляты обладают нитрилгидратазной и амидазной активностью и содержат гены амидаз двух типов, а также Fe-содержащей нитрилгидратазы. Не обнаружено нитрилазной активности и генов нитрилаз, Co-содержащих нитрилгидратаз и энантиоселективных амидаз. Изоляты способны формировать биопленки. Показано, что биомасса биопленок увеличивается при снижении температуры роста.
Гидролиз нитрилов, нитрилутилизирующие бактерии, содовое шламохранилище, биопленки, алкалофилы, галофилы, галоалкалотолерантные бактерии, нитрилгидратаза, амидаза
Короткий адрес: https://sciup.org/147252078
IDR: 147252078 | УДК: 579.22 : 579.26 | DOI: 10.17072/1994-9952-2025-3-279-288
Phylogenetic diversity of nitrile-utilizing bacteria of soda sludge storage facility and some of their physiological and biochemical features
The phylogenetic diversity of nitrile-utilizing bacteria from a soda sludge storage facility was studied. Haloalkalitolerant bacteria were isolated on acetamide and acetonitrile as the only carbon sources and identified by 16S rDNA sequencing. The isolates were found to belong to the phyla Actinomycetota (Arthrobacter, Microbacterium, Rhodococcus spp.) and Pseudomonadota (Acinetobacter, Ensifer, Pseudomonas, Rhizobium spp.). The isolates were found to have nitrile hydratase and amidase activity and contain genes for two types of amidases and Fe-containing nitrile hydratase. No nitrilase activity or nitrilase genes, Co-containing nitrile hydratases, or enantioselective amidases were found. Isolates are capable of forming biofilms, and biofilm biomass increases with decreasing growth temperature.
Текст научной статьи Филогенетическое разнообразие нитрилутилизирующих бактерий содового шламохранилища и их некоторые физиолого-биохимические особенности
Изучение микробного разнообразия экстремальных биотопов как естественного, так и антропогенного происхождения имеет большую перспективу для развития фундаментальной науки и биотехнологий. К щелочным и/или высокоминерализованным искусственным средам относятся щелочные сточные воды, солеот-валы, хранилища содового шлама. Содовые шламохранилища – это складированные на открытых полигонах отходы производства кальцинированной соды. Содовый шлам имеет щелочную реакцию (рН 11), содержит главным образом хлорид кальция, в нем практически отсутствуют питательные вещества, необходимые микроорганизмам, при этом он нестерилен. Методом метагеномного секвенирования было показано, что в содовом шламе обитают представители филумов Firmicutes, Proteobacteria, Bacteroidetes, Actinobacteria, Acidobac-teria, Verrucomicrobia и некоторых других [Шилова, Максимов, Максимова, 2020].
Способность микроорганизмов к гидролизу нитрилов активно изучается на протяжении последних нескольких десятилетий. Это связано с использованием нитрилгидролизующих микроорганизмов в биотехнологиях, в частности как биокатализаторов синтеза акриламида из акрилонитрила, никотинамида из 3-цианопиридина, и с перспективой получения различных производных амидов и карбоновых кислот в фарминдустрии и химической промышленности [Debabov, Yanenko, 2011]. При этом появление нитрилутилизирующих прокариотов в биотопах естественного происхождения может не быть связано с загрязнением синтетическими нитрилами. Цианогликозиды, синтезируемые растениями, обусловливают распространение нитрилутилизирующих бактерий в природе [Egelkamp, 2019].
Утилизация нитрилов микроорганизмами может происходить двумя метаболическими путями: одностадийным, при гидролизе нитрилазой (КФ 3.5.5.1) до соответствующих карбоновых кислот, и двустадийным, гидратацией нитрилов до амидов нитрилгидратазой (КФ 4.2.1.84) с последующим гидролизом до карбоновой кислоты амидазой (КФ 3.5.1.4). Биокатализатором этого процесса может быть как фермент, так и целая микробная клетка. Биокатализаторы, используемые в промышленности, в основном нейтрофильны, хотя возникает интерес к нитрилутилизирующим бактериям, выделенным из экстремальных биотопов, т. к. они могут быть полезны при осуществлении цельноклеточного катализа в жестких условиях: при повышенной температуре, крайних значениях рН, в присутствии органических растворителей.
Есть немногочисленные примеры выделения нитрилгидролизующих прокариотов из экстремальных сред. Natronocella acetinitrilica gen. nov. sp., способный к утилизации ацетонитрила как источника углерода и азота при экстремально высоком рН и среднем уровне минерализации, был выделен из осадка содового озера [Sorokin et al., 2007a]. Консорциум, состоящий из нового представителя актинобактерий и гаммапротеобактерий рода Marinospirillum , осуществлял утилизацию изобутиронитрила при рН 10 [Sorokin et al., 2007b]. Представитель рода Halomonas, выделенный из засоленных содовых почв, утилизировал 2-фенилпропионитрил в качестве источника азота при рН 10 [Chmura et al., 2008]. Облигатно алкало-фильный галотолерантный штамм Bacillus alkalinitrilicus sp. nov., растущий в присутствии изобутиронитрила, был выделен из солончаковой почвы [Sorokin, Van Pelt, Tourova, 2008]. Алкалотолерантный штамм R. pyridinivorans NIT-36, продуцирующий нитрилгидратазу, был выделен из горячего источника [Singh et al., 2017]. Выделены не только прокариотические, но и эукариотические микроорганизмы, способные к трансформации нитрилов в экстремальных условиях. «Черные дрожжи» Exophiala oligosperma R1 растут при pH 4, используя фенилацетонитрил в качестве единственного источника углерода, азота и энергии [Rustler et al., 2008]. Cолеустойчивый штамм дрожжей Meyerozyma guilliermondii LM2, способный конвертировать бензонитрил в среде, содержащей до 1.5 М NaCl, был выделен из подводных отложений [Serra et al., 2019]. Галофильные и галотолерантные микроорганизмы, утилизирующие нитрилы, могут функционировать как биокатализаторы в присутствии органических растворителей при низкой активности воды. Внутриклеточные ферменты алкалофилов часто проявляют наибольшую активность в нейтральном диапазоне рН, однако алкало- и галотолерантные нитрилутилизирующие бактерии могут служить цельноклеточным биокатализатором в условиях, отличающихся от физиологических.
Загрязненные среды антропогенного происхождения могут быть источником выделения штаммов, устойчивых как к высоким концентрациям токсикантов, так и к экстремальным воздействиям окружающей среды. Кроме этого, такие микроорганизмы обычно обладают механизмами неспецифической устойчивости, которые позволяют им приспосабливаться к воздействию самых разных неблагоприятных факторов. В связи с этим изучение филогенетического разнообразия нитрилутилизирующих бактерий в таком биотопе, как содовое шламохранилище, представляется достаточно важным. Целью настоящей работы явилось выделение и идентификация нитрилутилизирующих бактерий, изучение их биохимических свойств и способности к биопленкообразованию под воздействием неблагоприятных факторов окружающей среды.
Материалы и методы исследования
Объекты исследования и условия культивирования
Бактериальные изоляты выделяли из проб содового шлама и грунта старой карты содового шламо-хранилища АО «Березниковский содовый завод» (г. Березники, Пермский край) на агаризованной минеральной среде (0.05% NaCl, рН 7.2) c 10 мМ ацетамидом (ацд) или 0.25% ацетонитрилом (ацн) в качестве единственных источников углерода и азота. Грунт отбирали с поверхности, на глубине 5 и 10 см, также для выделения использовали объединенную пробу. Сочетание высокой концентрации соли, щелочной среды и ацетамида/ацетонитрила как единственных источников энергии не позволило получить изоляты на агаризованной среде.
Идентификация полученных изолятов
Видовая идентификация бактериальных изолятов была проведена путем ПЦР-анализа с праймерами к генам 16S рРНК, сконструированными на основе последовательностей, представленных в базе данных GenBank, и синтезированными ООО «Евроген» (г. Москва). Препараты хромосомной ДНК получали методом щелочного лизиса. Амплификацию ДНК проводили с применением термостабильной Taq-полимеразы производства ООО «СибЭнзим» (г. Новосибирск) на термоциклере Т3 («Biometra», Германия). Электрофоретическое разделение продуктов ПЦР-реакции проводили в 1.2–1.5% агарозном геле в трисборатном буфере при напряженности поля 5 В/см. Для оценки молекулярной массы фрагментов ДНК использовали молекулярные маркеры 1 kb и 100 b (ООО «СибЭнзим» и Axigen®). Визуализацию полос и документирование данных осуществляли после окрашивания геля бромистым этидием с использованием системы гельдокументации BioDocAnalyze («Biometra», Германия). Очистку ПЦР-продукта перед секвенированием осуществляли двумя способами: с помощью смеси ферментов ExoSAPMix (Fermentas Life Sciences) и аппарата E--Gel («Invitrogen», США) согласно инструкции фирмы-производителя. Секвенирование проводили на приборе Genetic Analyzer 3500xl («Applied Biosystems», США), следуя инструкциям фирмы-производителя. Гомологию полученных нуклеотидных последовательностей генов 16S рРНК с известными генами микроорганизмов анализировали с использованием программного пакета BLAST ; программы Chromas lite 2.1. Сравнение нуклеотидных последовательностей проводили с применением программ ClustalW 2.0.9 и YACWGUI 1.2.
Определение нитрилгидратазной и амидазной активности изолятов
Нитрилгидратазную активность определяли по образованию акриламида и акриловой кислоты из акрилонитрила. Изоляты выращивали до стационарной фазы, культуральную жидкость (1 мл) центрифугировали 10 мин при 4500 g, клетки отмывали калий-фосфатным буфером (рН 7.2) и центрифугировали повторно. Клеточный преципитат суспендировали в 1 мл фосфатного буфера и вносили акрилонитрил до конечной концентрации 0.3 М, через 24 ч реакцию останавливали добавлением 50 мкл HCl. Амидазную активность определяли по образованию акриловой кислоты из акриламида. Реакцию проводили 24 ч, субстрат добавляли в концентрации 50 мМ. Концентрацию акриламида и акриловой кислоты в реакционной среде определяли методом ВЭЖХ на хроматографе Infinity II JC 1260 (Agilent, Германия) с колонкой Synergi 4u Hydro–RP 80A (250 × 4.6 мм). В качестве подвижной фазы использовали 25 мМ NaH 2 PO 4 с 5% ацетонитрила, скорость потока составляла 0.500 мл/мин при 25°C, детекцию проводили при длине волны 200 нм.
Детекция генов амидаз, нитрилгидратаз и нитрилаз
Амплификацию ДНК проводили с применением термостабильной Taq-SE ДНК-полимеразы (ООО «СибЭнзим», г. Новосибирск) на термоциклере Т100 («Biometra», Германия). Для анализа генов использованы разработанные ранее олигонуклеотидные праймеры [Демаков и др., 2009a; Демаков и др., 2009б; Павлова, Неустроева, Максимов, 2011]. Режим амплификации включал начальный цикл денатурации –
1 мин при 94°С; денатурацию, 94°С – 20 с; отжиг, 53° – 30 с; элонгацию, 72°С – 60 с; (35 циклов) и завершающий этап – 60 с при 72°С. Электрофоретическое разделение ПЦР-продуктов проводили в 1.2% агарозном геле в трис-боратном буфере при напряженности электрического поля 6 В/см. Праймеры для выявления генов ферментов, участвующих в трансформации нитрилов и амидов, представлены в табл. 1 .
Праймеры для выявления генов амидаз, нитрилгидратаз, нитрилаз
Таблица 1
[Primers for detecting the genes of amidases, nitrile hydratases, and nitrilases]
|
Выявляемые гены |
Праймер |
Последовательность, 5’-3’ |
Длина ПЦР-продукта |
|
Амидаза R. erythropolis (E12517) и R. rhodochrous N-774 (X54074) |
ARh499f ARh1002r |
GAAGCAGGTGGATCATCCGG TCCGTGGCGATCACGTTCCA |
503 |
|
Амидаза R. erythropolis (M88714) |
AReR-1 AReR-2 |
ATGCGACACGGTGACATCTCCTCGA TTACGCTTCGACGGTCTTCTCGAC |
1094 |
|
Амидаза R. rhodochrous J1 (D16207) |
AmiJ1-1 AmiJ1-2 |
ATGTCTTCGTTGACTCCCCCCAATT TTATGTCAGGGTGCCGGCTGCAGC |
1547 |
|
Амидаза Rhodococcus sp. (A19131) |
ARsp-1 ARsp-2 |
ATGGGCTTGCATGAACTGACGCTCG TCAAAGCGGCGCCAGTCGCGGCCA |
1388 |
|
α-субъединица Со-нитрилгидратазы R. rhodochrous J1 (D67027) |
NHCoAF NHCoAR |
GTGATACATATGAGCGAGCACGTCAAT ATGCATCATACGATCACTTCCTG |
611 |
|
β- субъединица Со-нитрилгидратазы R. rhodochrous J1 (D67027) |
NHCoBF NHCoBR |
GTGATACATATGGATGGTATCCACGAC ATGCATCACGCAGAGATCAGGTA |
689 |
|
α- субъединица Fe-нитрилгидратазы R. rhodochrous N-774, (X54074) |
NHFeAF NHFeAR |
СATATGTCAGTAACGATCGAC ATGCATCAGACGGTGGGAACCTG |
623 |
|
β- субъединица Fe-нитрилгидратазы R. rhodochrous N-774 (X54074) |
NHFeBF NHFeBR |
СATATGGATGGAGTACACGAT ATGCATCAGGCCGCAGGCTCGAG |
638 |
|
Нитрилаза R. rhodochrous K22 (D12583) |
NitK22F NitK22R |
ATGTCCAGCAATCCAGAGCTCAAGTACAC CTAGGCCTCCGCCTTGGCCC |
1151 |
|
Нитрилаза R. rhodochrous J1 (D67026) |
NitJ1F NitJ1R |
ATGGTCGAATACACAAACACATTCAAAG TCAGATGGAGGCTGTCGCCC |
1100 |
Определение массивности биопленок
Биопленки выделенных изолятов выращивали в 96-луночном полистироловом планшете (Медполи-мер, Россия) в БТН-бульоне (Биотехновация, Россия), варьируя условия культивирования. Для изучения влияния минерализации в среду добавляли 5, 10, 50 г/л NaCl, рН среды доводили с помощью гидроксида натрия (NaOH), культивирование проводили при температуре 8, 15, 30 и 37°С. Биопленки в среде с различной концентрацией соли и рН выращивали при температуре 30°С. Влияние температуры на образование биопленок изучали на среде БТН с рН 7. После 7‒10 суток инкубации планктонные клетки удаляли из лунок декантацией, дважды отмывали биопленку 200 мкл калий-фосфатным буфером и определяли массивность образованной биопленки. Биопленку окрашивали 0.1% кристаллическим фиолетовым в течение 40 мин в темноте, удаляли краситель, отмывали 1 раз калий-фосфатным буфером и экстрагировали краситель 200 мкл 96% спирта. Формирование биопленок оценивали по оптической плотности раствора красителя при λ 540 нм на планшетном ридере Infinite M1000 pro, «TECAN» (Швейцария).
Статистическую обработку проводили с использованием t-критерия Стьюдента, различия считали значимыми при p < 0.05.
Результаты и их обсуждение
Из содового шлама и восстанавливающихся после эксплуатации грунтов территории содового шла-мохранилища АО «Березниковский содовый завод» на минеральной среде с ацетамидом и ацетонитрилом в качестве единственного источника углерода и энергии были выделены изоляты нитрил- и амиду-тилизирующих бактерий, которые идентифицировали методом секвенирования гена 16S рДНК. Результаты идентификации представлены в табл. 2.
Таблица 2
Идентификация нитрилгидролизующих бактерий, выделенных на территории содового шламохранилища АО «Березниковский содовый завод»
[Identification of nitrile hydrolyzing bacteria isolated on the territory of the soda sludge storage facility of JSC Bereznikovsky Soda Plant]
|
Изолят |
Источник выделения |
Вид |
Типовой штамм |
Сходство, % |
Количество прочт. нукл-ов |
Таксономия |
|
Источник углерода – ацетонитрил |
||||||
|
ССОЗ1 (ацн) |
Грунт, старая карта |
Rhizobium radiobacter |
ATCC 19358(T) |
100.00 |
809 |
Bacteria, Pseudo-monadota, Alphaproteobacteria, Rhizobiales, Rhizobi-aceae, Rhizobium |
|
ССОП 1 (ацн) |
Поверхность грунта, старая карта |
Rhizobium nepotum |
39/7(T) |
99.75 |
807 |
|
|
ССОП (ацн) |
Поверхность грунта, старая карта |
Pseudomonas mandelii |
NBRC 103147(T) |
99.87 |
782 |
Bacteria, Pseudo-monadota, Gammaproteobacteria, Pseudomonadales, Pseudomonadaceae, Pseudomonas |
|
ССОЗ 1.1(ацн) |
Грунт, старая карта |
Microbacterim hatanonis |
JCM 14558(T) |
98.42 |
699 |
Bacteria, Actinomyce-tota, Actinomycetes, Micrococcales, Microbacteriaceae, Microbacterium |
|
ОШ2 / 5Э |
Шлам, новая карта |
Rhodococcus qingshengii |
JCM 15477(Т) |
99.42 |
537 |
Bacteria, Actinomyce-tota, Actinomycetes, Corynebacteriales, Nocardiaceae, Rhodococcus |
|
ССОЗ 5 2 / 6Э |
Грунт, 5 см глубины, старая карта |
Rhodococcus erythropolis |
NBRC 15567(Т) |
99.84 |
651 |
|
|
Источник углерода – ацетамид |
||||||
|
ССОЗ1 (ацд) |
Грунт, старая карта |
Ensifer morelensis |
Lc04(T) |
98.99 |
701 |
Bacteria, Pseudo-monadota, Alphaproteobacteria, Hyphomicrobiales, Rhizobiaceae, Ensifer |
|
CCОЗ 1.3 (ацд) |
Грунт, старая карта |
Ensifer morelensis |
Lc04(T) |
99.87 |
781 |
|
|
ССОЗ 10 4 (ацд) |
Грунт, старая карта, 10 см глубины |
Ensifer morelensis |
Lc04(T) |
100.00 |
847 |
|
|
ССОЗ 1.2 (ацд) |
Грунт, старая карта |
Pseudomonas sp. |
DSM 18928(T) |
99.53 |
853 |
Bacteria, Pseudo-monadota, Gammaproteobacteria, Pseudomonadales, Pseudomonadaceae, Pseudomonas |
|
ССОЗ 3(ацд) |
Грунт, старая карта |
Pseudomonas mandelii |
NBRC 103147(T) |
98.00 |
751 |
|
|
CCОЗ 1.4 (ацд) |
Грунт, старая карта |
Rhodococcus sp. |
JCM 15477(T) |
100 |
666 |
Bacteria, Actinomyce-tota, Actinomycetes, Corynebacteriales, Nocardiaceae, Rhodococcus |
|
ССОЗ 10 3 |
Грунт, старая карта, 10 см глубины |
Rhodococcus erythropolis |
NBRC 15567 (Т) |
99.85 |
701 |
|
Окончание табл. 2
|
Изолят |
Источник выделения |
Вид |
Типовой штамм |
Сходство, % |
Количество прочт. нукл-ов |
Таксономия |
|
ССОЗ 10 1 (ацд) |
Грунт, старая карта, 10 см глубины |
Arthrobacter globiformis |
NBRC 12137(T) |
100.00 |
771 |
Bacteria, Actinomyce-tota, Actinomycetes, Micrococcales, Micrococcaceae, Arthrobacter |
|
ССОП 1 (ацд) |
Грунт, старая карта, поверхность |
Acinetobacter calcoaceticus |
DSM 30006(T) |
99.87 |
876 |
Bacteria, Pseudo-monadota, Gammaproteobacteria, Pseudomonadales, Moraxellaceae, Aci-netobacter |
|
ССОП 1.2 (ацд) |
Грунт, старая карта, поверхность |
Rhizobium nepotum |
39/7(T) |
99.77 |
876 |
Bacteria, Pseudo-monadota, Alphaproteobacteria, Rhizobiales, Rhizobi-aceae, Rhizobium |
Нитрилутилизирующие бактерии были выделены главным образом из грунта старой карты шламо-хранилища, который характеризовался высокой минерализацией и был слабощелочным (рН 8) [Шилова, Максимов, Максимова, 2020]. Ранее из различных сред содового шламохранилища (содового шлама, грунта восстанавливающихся территорий) были выделены бактериальные штаммы, обладающие различной гидролитической активностью [Шилова, Максимов, Максимова, 2021] и устойчивостью к широкому спектру рН и высокой минерализации [Maksimova, Eliseeva, Maksimov, 2024]. На грунте старой карты шламохранилища наблюдалось восстановление растительности. Содовый шлам состоял из ионов кальция, хлора, карбонатов и гидрокарбонатов и имел рН 11, что приводило к значительному снижению микробного разнообразия по сравнению с грунтом старой карты. Из содового шлама на ацетонитриле был изолирован только один штамм галоалкалотолерантных родококков. Выделенные изоляты принадлежали филумам Actinomycetota и Pseudomonadota.
Определена активность выделенных изолятов. Показано, что изоляты обладают как амидазной активностью, которая проявляется при гидролизе акриламида, так и нитрилгидратазной активностью, выявляющейся при трансформации акрилонитрила. Когда субстратом реакции является акрилонитрил, в реакционной смеси обнаруживается как акриламид, так и акриловая кислота, что подтверждает работу нит-рилгидратазно-амидазной ферментативной системы. При гидролизе акриламида в реакционной среде детектировали акриловую кислоту. Ферментативные активности изолятов представлены в табл. 3. Нит-рилгидратазная активность обычно значительно превышает амидазную, особенно у селекционированных продуцентов, однако у большинства изолятов, выделенных с территории содового шламохранилища, амидазная активность была выше нитрилгидратазной.
Известно, что у родококков встречается несколько структурно-различных групп амидаз. Из щелочного высокоминерализованного биотопа были выделены бактериальные изоляты различной филогенетической принадлежности, в том числе изоляты родококков. Методом ПЦР определено наличие в геномах исследованных алкалотолерантных изолятов генов, соответствующих генам амидаз следующих типов: амидаза R. erythropolis (E12517) и R. rhodochrous N-774 (X54074), алифатическая амидаза R. erythropolis (M88614, Genbank), энантиоселективная амидаза R. erythropolis (AY026386, Genbank). Установлено, что в ДНК всех исследованных экстремотолерантных изолятов есть гены амидаз 1-го и 2-го типов (рис. 1), но не обнаруживается энантиоселективная амидаза. У всех штаммов обнаружены гены Fe-cодержащей нит-рилгидратазы (рис. 2) и отсутствует Co-содержащая. Гены нитрилаз не обнаружены.
Образование биопленок является адаптивной реакцией бактерий. Ранее нами было показано, что Rhodococcus qingshengii IEGM 1416 (изолят ОШ2 / 5Э) и R. erythropolis IEGM 1417 (изолят ССОЗ 5 2 / 6Э) формировали более массивные биопленки при увеличении рН и снижении температуры, а R. qingshengii IEGM 1416 – еще и при повышении концентрации соли в среде [Maksimova, Syrovatskaya, Maksimov, 2025]. Изучено биопленкообразование выделенных на ацетамиде и ацетонитриле изолятов нитрилутилизирующих бактерий. Изоляты формировали биопленки, биомасса которых, оцененная по ОП 540 экстрагированного красителя кристаллического фиолетового, составляла от 0.2 до 0.4 единиц. Наибольшую биомассу (ОП 540 = 0.4–0.5) отмечали у биопленок Acinetobacter calcoaceticus , Pseudomonas mandelii и
Rhizobium nepotum. Оценили воздействие 5–50 г/л хлорида натрия, 7–12 рН и 8–37ºС на формирование биопленок у изолятов, проявивших наибольшую способность к биопленкообразованию (рис. 3).
Таблица 3
Нитрилгидратазные и амидазные активности бактериальных изолятов, выделенных на территории содового шламохранилища АО «Березниковский содовый завод»
[Nitrile hydratase and amidase activities of bacterial isolates isolated on the territory of the soda sludge storage facility of JSC Bereznikovsky Soda Plant]
|
Субстрат реакции (фермент) Изолят |
Акрилонитрил (нитрилгидратаза) |
Акриламид (амидаза) |
|
Активность фермента, ммоль/г/ч |
||
|
ССОП (ацн) |
1.63 |
2.84 |
|
ССОП 2 (ацд) |
3.31 |
33.38 |
|
ССОЗ 10 2 (ацд) |
27.19 |
14.21 |
|
ССОЗ 10 4 (ацд) |
15.23 |
42.99 |
|
ССОЗ 10 3 (ацд) |
26.79 |
10.73 |
|
ССОЗ 5 1 (ацн) |
6.50 |
28.19 |
|
ССОЗ 2 (ацн) |
26.14 |
38.03 |
|
ССОЗ 2 (ацд) |
23.67 |
37.61 |
|
ССОЗ 10 (ацн) |
1.89 |
16.18 |
|
ССОЗ 4 (ацд) |
19.09 |
49.05 |
|
ССОЗ 10 1 (ацд) |
0.97 |
0.87 |
|
ССОП 1 (ацд) |
0.42 |
3.39 |
|
ССОЗ 3 (ацд) |
19.76 |
33.23 |
|
ССОЗ 1 (ацн) |
7.77 |
14.49 |
|
ССОЗ 3 (ацн) |
1.60 |
17.41 |
|
ССОЗ 1 (ацд) |
33.24 |
22.80 |
Примечание: независимые эксперименты по культивированию изолятов и определению ферментативной активности их клеток проведены в двухкратной повторности. В таблице приведены результаты единичного эксперимента.
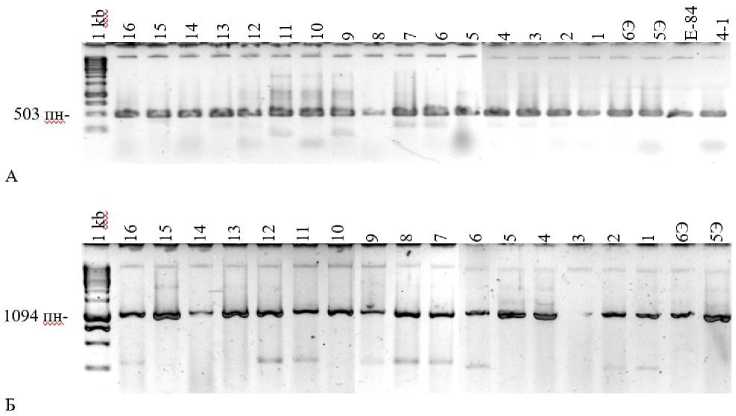
Рис. 1. ПЦР-продукты, соответствующие генам амидаз, гомологичных таковым из R. erythropolis , E12517, R. rhodochrous N-774, X54074 (А) и R. erythropolis , M88714 (Б).
Изоляты: 1 – ССОП (ацн), 2 – ССОП2 (ацд), 3 – ССОЗ 10 2 (ацд), 4 – ССОЗ 10 4 (ацд), 5 – ССОЗ 10 3 (ацд) 6 – ССОЗ 5 1 (ацн), 7 – ССОЗ2 (ацн), 8 – ССОЗ2 (ацд), 9 – ССОЗ 10 (ацн), 10 – ССОЗ4 (ацд), 11 – ССОЗ 10 1 (ацд), 12 – ССОП1 (ацд), 13 – ССОЗ3 (ацд), 14 – ССОЗ1 (ацн), 15 – ССОЗ3 (ацн), 16 – ССОЗ1 (ацд), R. qingshengii IEGM 1416 (5Э), R. erythropolis IEGM 1417 (6Э), R. erythropolis Е-84, R. erythropolis 4-1
[PCR products corresponding to the genes of amidases homologous to those from R. erythropolis , E12517, R. rhodochrous N-774, X54074 (A) and R. erythropolis , m88714 (B).
Isolates: 1 – SSOP (acn), 2 – SSOP2 (acd), 3 – SSOZ102 (acd), 4 – SSOZ104 (adc), 5 – SSOZ103 (adc), 6 – SSOZ51 (adc), 7 – SSOZ2 (adc), 8 – SSOZ2 (adc), 9 – SSOZ10 (adc), 10 – SSOZ4 (adc), 11 – SSOZ101 (adc), 12 – ssop1 (ADC), 13 – ccos3 (ADC), 14 – ccos1 (ADC), 15 – ccos3 (ADC), 16 – ccos1 (ADC), R. qingshengii IEGM 1416 (5E), R. erythropolis IEGM 1417 (6E), R. erythropolis E-84, R. erythropolis 4-1]
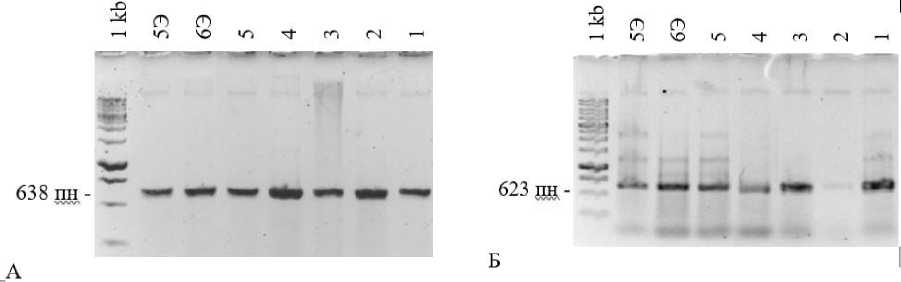
Рис. 2. ПЦР-продукты, соответствующие генам альфа (А) и бета (Б) субъединиц Fe-содержащей нитрилгидратазы, гомологичных таковым из R. rhodochrous N-774, X54074.
Обозначение изолятов как на рис. 1
[PCR products corresponding to the alpha (A) and beta(B) subunit genes of Fe-containing nitrile hydratase, homologous to those of R. rhodochrous N-774, X54074.
Designation of isolates as shown in Fig. 1]
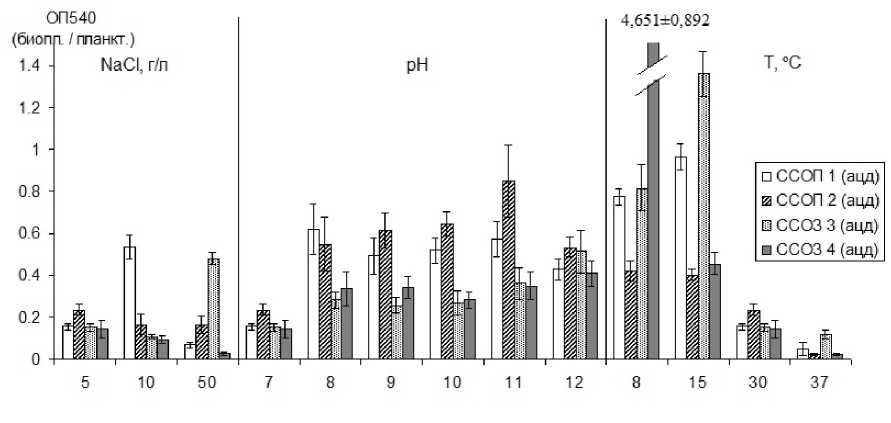
Рис. 3. Влияние концентрации NaCl, рН и температуры на биопленкообразование изолятов нитрилутилизирующих бактерий содового шламохранилища
[Influence of NaCl concentration, pH, and temperature on biofilm formation of isolates of nitrilutylizing bacteria from soda sludge storage]
Высокие концентрации соли подавляли рост изолятов, только у A. calcoaceticus ССОП 1 и P. mandelii значительно увеличивалась биомасса биопленки в среде с 10 и 50 г/л NaCl соответственно. Влияние рН было более выражено для R. nepotum, биомасса биопленок увеличивалась при возрастании рН от 7 до 11, но снижалась при рН 12. У всех изолятов образование биопленок было достоверно выше в щелочной среде (p < 0.05), чем при рН 7. Температурное воздействие на биопленкообразование было сходным у всех изученных изолятов: при температуре 37ºС формирование биопленок подавлялось, тогда как планктонная культура росла достаточно активно (ОП 540 = 0.7–0.8). Снижение температуры до 8–15ºС интенсифицировало процесс образования биопленок.
Заключение
Таким образом, с поверхности, глубины 5–10 см и из объединенной пробы грунта старой карты содового шламохранилища на ацетамиде и ацетонитриле как единственном источнике углерода были выделены изоляты нитрилутилизирующих бактерий, относящиеся к филумам Actinomycetota и Pseudomonadota. Единственный штамм родококков, способный к утилизации нитрилов, был выделен из содового шлама с экстремально высокими рН и минерализацией. Изоляты обладали активностью нит-рилгидратазы и амидазы, причем амидазная активность была выше нитрилгидратазной. Обнаружены гены неэнантиоселективных амидаз двух типов и Fe-cодержащей нитрилгидратазы. Выделенные изоляты формировали биопленки, биомасса которых увеличивалась при снижении температуры роста. Воздействие других изученных факторов (концентрации NaCl и рН) на образование биопленок было штаммоспецифично.
