Фитоценотическое разнообразие березовых неморальнотравных лесов юго-западного Подмосковья

Автор: Беляева Н.Г., Нешатаева В.Ю.
Журнал: Фиторазнообразие Восточной Европы @phytodiveuro
Статья в выпуске: 1 т.11, 2017 года.
Бесплатный доступ
Для юго-западной части Московской области выявлено ценотическое разнообразие сообществ группы ассоциаций неморальнотравных березовых лесов. В результате разработанной эколого-фитоценотической классификации было выделено 2 группы ассоциаций, 4 ассоциации, 5 субассоциаций и 6 вариантов. Проанализированы особенности видового состава выделенных синтаксонов. Проанализировано расположение геоботанических описаний в экологическом пространстве. Установлено, что сообщества ассоциаций образуют в пространстве переменных факторов (видов) непересекающиеся кластеры.
Антропогенные леса неморальнотравные березняки классификация ординация московская область
Короткий адрес: https://sciup.org/148314592
IDR: 148314592 | УДК: 581.526.425
Текст научной статьи Фитоценотическое разнообразие березовых неморальнотравных лесов юго-западного Подмосковья
В настоящее время производные мелколиственные сообщества в Подмосковье занимают большие площади. Они формируются в результате зарастания заброшенных сельхозугодий, также возникают на месте вырубок или плохо прижившихся лесных культур. Московский регион подвергался сильному антропогенному воздействию на протяжении многих столетий, что привело к полному исчезновению здесь коренных лесов. О составе лесного покрова в прошлом известно по «Экономическим примечаниям к планам генерального межевания». Так, уже к концу XVIII в. леса исследуемой территории в подавляющем большинстве были образованы осиновыми и березовыми («дровяными») древостоями (Беляева, Попов, 2016). В настоящее время производные мелколиственные сообщества также играют большую роль в составе лесного покрова, что связано с зарастанием сельхозугодий на обширных площадях (Hansen et al., 2016). Несмотря на это, эти сообщества изучены довольно слабо (Ниценко, 1972). В последнее время проблема классификации производных лесных сообществ, и не только мелколиственных, встает все более остро (Цвирко и др., 2016). Сложности их классификации связаны со значительным флористическим и ценотиче-ским разнообразием производных лесов, нестабильностью их динамического состояния и не столь очевидными связями сообществ с условиями произрастания.
В настоящей работе мы рассматриваем наиболее широко распространенные сообщества мелколиственных суходольных лесов Европейской части России – березовые леса, а среди них – неморальнотравные березняки. Березовые леса Подмосковья образованы двумя видами берез – Betula pendula и Betula pubescens. Хотя эти виды и различаются морфологически, в природе они нередко произрастают вместе, образуя гибриды и имея «…самые разнообразные комбинации их отличительных признаков…» (Сукачев, 1938:15). А.А. Ниценко (1972) указывает, что почти никто из исследователей восточноевропейских березняков не выделял отдельные формации по видам берез. Вслед за А.А. Ниценко (1972) и В.И. Василевичем (1996) мы также рассматриваем сообщества березняков, объединяя их в одну формацию Betuleta pendulae-pubescentis. При анализе
вторичных лесов наиболее полным является подход, учитывающий генезис березняков, с одной стороны, и особенности антропогенного влияния на них, с другой. Так, А.А. Ни-ценко (1972) выделил большое количество мелких ассоциаций, которые он объединил в группы как по изначальным условиям местообитания, в котором возникло производное сообщество, так и по свойствам, которое это сообщество приобрело в процессе трансформации (например, асс. «нитрофильные березняки» и асс. «олуговелые нитрофиль-ные березняки»). Практически для каждого типа местообитания в его работе выделена олуговелая группа, которая возникает при значительном изменении почвенного и растительного покрова, а также при сильном осветлении. Ю.Д. Абатуров с соавт. (1982) выделяют группы типов леса на основе их динамических связей с коренными еловыми и сосновыми лесами южной тайги. В настоящей работе мы не рассматриваем вопросы антропогенного влияния на сообщества березовых лесов, поскольку все изученные нами неморальнотравные березняки в настоящее время его не испытывают. Что касается их динами- ческих связей с коренными типами леса, то мы, придерживаясь мнения С.Ф. Курнаева (1968), считаем коренными лесами водораздельных пространств территории исследований липово-еловые неморальнотравные леса. Неодинаковое участие широколиственных пород в составе двух выделенных групп ассоциаций, скорее всего, объясняется различным антропогенным влиянием на них в прошлом. Объекты и методы
Территория исследований находится в западной части Наро-Фоминского района Московской области, на отрогах Смоленско-Московской возвышенности (рис. 1).
Фундамент территории исследований сложен известняками среднего карбона, который перекрыт мореной, водноледниковыми отложениями, а сверху, зачастую, покровными суглинками. Почвенный покров сформирован на покровных и водноледниковых суглинках. Почвы преимущественно дерново-среднеподзолистые, по понижениям –оглеенные (Анненская и др., 1997). Территория исследований находится в зоне хвойно-широколиственных лесов вблизи ее южной границы.
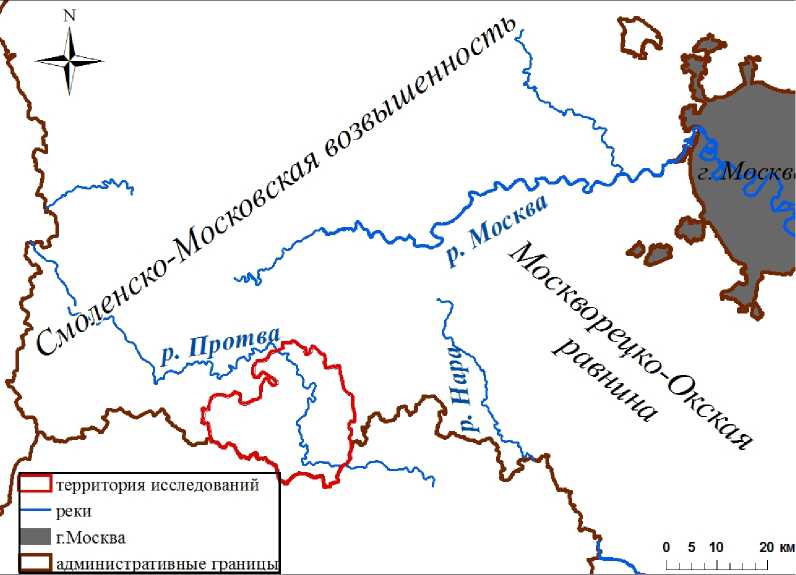
Рис. 1. Расположение территории исследований
Location of the study area
Геоботанические описания (n=16) были выполнены в течение 4-х полевых сезонов (2013-2016 гг.). Размер пробной площади составлял 20×20 м. Проективное покрытие видов определяли в процентах. При классификации сообществ березовых лесов мы использовали эколого-фитоценотический подход. Группы ассоциаций выделяли по составу видов древостоя (принадлежность берез к двум разным видам не учитывали), а также по преобладающим эколого-биоморфологическим группам травянокустарничкового яруса. Кроме того, учитывали структуру сообществ: например, наличие кустарникового яруса. Так, мы выделяли березняки неморальнотравные (простые) и березняки неморальнотравные сложные (со значительным участием в древостое широколиственных пород). Термин «сложные» первоначально был использован при классификации еловых лесов (Сукачев, 1938); однако впостледствии его стали употреблять для обозначения производных сообществ с участием широколиственных пород (Абатуров, 1982; Василевич 1996). Основной низшей единицей классификации является ассоциация, принадлежность к которой определяли, используя принципы отечественной геоботанической школы. К одной ассоциации относили фитоценозы, сходные по составу доми-нантов выраженных ярусов, в том числе – по их количественному соотношению, а также с учетом сходства группы менее обильных второстепенных видов (Шенников, 1964). Таким образом, учитывали видовой состав и структуру сообществ. Субассоциации выделяли по различиям в обилии доминантов и содоминантов (субдоминантов), а также по наличию содоминантов (субдоминантов), не типичных для ассоциации (Нешатаева, 2009). Названия видов сосудистых растений приводим по С.К. Черепанову (1995). Анализ распределения сообществ в экологическом пространстве был проведен с помощью метода непрямой ординации – многомерного неметрического шкалирования NMDS ( non-metric multidimensionalscaling ).
Результаты
Несмотря на то, что при анализе мы не делили сообщества на формации по видовой принадлежности берез, можно приблизительно оценить преобладание какого-либо вида в различных сообществах. Участие видов берез в сложении древостоя зависит от типа сообществ (Абесадзе, 1983). Так, в лесах с участием Tilia cordata отмечена только Betula pubescens , а в лесах с дубом – Betula pendula . Основной породой подлеска является лещина ( Coryllus avellana ): она формирует сомкнутый кустарниковый ярус, определяя световой режим в сообществах. Стоит отметить большое участие Athyrium filix-femina в сообществах березняков исследуемой территории. Данный вид в лесных сообществах приурочен к пониженным элементам рельефа с достаточной обеспеченностью влагой, с одной стороны, и к осветленным участкам, с другой стороны (Биологическая…, 1983). Экологический режим березовых неморальнотравных лесов, если они находятся в пониженных и влажных местообитаниях, благоприятен для Athyrium filix-femina . Этот папоротник пышно развивается в небольших осветленных местах, в промежутках между ажурными кронами берез. К тому же, он не требует наличия древесных остатков (вале-жа) для своего развития, которых не бывает в сообществах, образовавшихся на вырубках или залежах, в отличие, например, от Dryop-teris filix-mas (Биологическая…, 1983). Моховой покров в неморальнотравных березовых лесах выражен очень слабо. Наиболее часто здесь встречается Atrichum undulatum. Среднее проективное покрытие мхов составляет 9 %, значения общего покрытия мохового яруса от 0 до 15 %; лишь на одной пробной площади – 30 %.
Группа ассоциаций Betuleta nemoriher-bosa – березняки неморальнотравные
Средняя сомкнутость древостоя 0,71. Для сообществ группы характерен такой же состав травяного яруса, как и в неморальнотравных группах ассоциаций с другими лесо- образователями – доминантами древостоя (из ели, широколиственных пород). В сообществах группы присутствует подрост ели, а также постоянно, но в небольших количествах – дуба. Наиболее константной является группа неморальных видов: Ajuga reptans, Ranunculus cassubicus, Dryopteris carthusiana, Galeobdolon luteum, Stellaria holostea, Ae-gopodiumpodagraria, Carexpilosa, Pulmonaria obscura. В их числе отмечены виды, имеющие свой оптимум не в широколиственных лесах, а в южной тайге и зоне хвойношироколиственных лесов. Характерна также группа бореальных видов-мезофитов: Oxalis acetosella, Rubus saxatilis, Luzula pilosa, а также виды, приуроченные к более влажным местообитаниям – Geum rivale, Deschampsia cespitosa. Высокая встречаемость щучки дернистой в производных мелколиственных сообществах Московской области отмечена в литературе (Растительность…, 1996).
Ассоциация 1. Betuletum athyriosum – березняк кочедыжниковый. Сообщества ассоциации отличаются низкой сомкнутостью древостоя (0.5). В древостое присутствуют обе березы, как Betula pendula , так и Betula pubescens . Характерно очень малое участие лещины. В подлеске присутствует Salix caprea , а в подросте – ель и дуб. Доминантом травяно-кустарничкового яруса является Athyrium filix-femina . В сообществах ассоциации характерно значительное участие Deschampsia cespitosa. Данная ассоциация является переходной к нитрофильной группе ассоциаций, для которой характерна высокая ценотиче-ская роль кочедыжника женского.
Ассоциация 2. Betuletum galeobdolosum – березняк зеленчуковый. Сообщества ассоциации характеризуются более сомкнутым древостоем (0,73) и развитым кустарниковым ярусом из лещины; ее среднее проективное покрытие составляет 25 %. В древостое, кроме березы, встречаются Picea abies, Populus tremula, Quercus robur, Acer platanoides и Alnus incana. Зеленчуковый вариант в составе асс. Aegopodio-Betuletum (березняк неморальнотравный) был выделен В. И. Василевичем (1996).
Субасс. caricosum pilosae – волосистоосоковая. Субассоциация выделена по высокому обилию осоки волосистой, которая со-доминирует наравне с зеленчуком. По наблюдениям ряда исследователей, сообщества с осокой волосистой приурочены к плоским вершинам всхолмлений (Рысин, Савельева, 2002). Характерно наличие в сообществах подроста ели и дуба, но в небольших количествах.
Var . Geum rivale – вариант с гравилатом речным. Сообщества варианта отличаются бóльшим участием нитрофильных видов в составе травяно-кустарничкового яруса ( Athy-rium filix-femina, Geum rivale, Crepis paludosa ). Они также отличаются от других сообществ по участию Rubus saxatilis и Fragaria vesca.
Субасс. typicum – типичная. В сообществах субассоциации в травяном ярусе из доми-нантов отмечен только Galeobdolon luteum . Подобные сообщества приурочены к верхним частям склонов всхолмлений (Рысин, Савельева, 2002). Характерно значительное участие в подросте ели (табл.).
Субасс. aegopodiosum – снытевая. В сообществах субассоциации содоминируют зеленчук и сныть. Леса со значительным участием сныти встречаются на выположенных участках или склонах с незначительным уклоном (Рысин, Савельева, 2002). Здесь также наблюдается массовый подрост ели.
Var. Oxalis acetosella – вариант кисличный. Выделен по значительному обилию кислицы (покрытие 10-20 %); в сообществах варианта также наблюдается участие бореальных видов – Gymnocarpium dryopteris, Maian-themum bifolium, Trientalis europae.
Таблица. Геоботаническая характеристика березняков неморальнотравных
Phytocenological characteristic of nemoral birch forests
|
Группа ассоциаций |
||||||||||||||||
|
Betuleta nemoriherbosa |
Betuleta nemoriherbosa composita |
|||||||||||||||
|
Ассоциация |
||||||||||||||||
|
§ 1 II К) tq |
Betuletum galeobdolosum |
Betuletum quercoso-corylosum |
Betuletum tilioso-caricosum pilosae |
|||||||||||||
|
Субассоциация |
||||||||||||||||
|
1 |
2 |
3 |
4 |
5 |
||||||||||||
|
Вариант |
||||||||||||||||
|
1а 1 |
3а 1 |
4а 1 |
4b 1 |
4c 1 |
5а 1 |
|||||||||||
|
Номера описаний |
||||||||||||||||
|
Ярусы и виды |
oo (S |
о |
co \o |
о о |
J C9 |
Й |
6 |
о |
oo о |
xo n |
3 |
m1 |
s |
2 |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
|
|
Древостой, сомкнутость |
0.5 |
0.8 |
0.7 |
0.9 |
0.85 |
0.6 |
0.6 |
0.7 |
0.8 |
0.7 |
0.4 |
0.7 |
0.7 |
0.5 |
0.9 |
0.9 |
|
Betulapendula, % |
40 |
65 |
65 |
50 |
40 |
55 |
45 |
70 |
70 |
45 |
20 |
25 |
20 |
40 |
||
|
Betulapubescens Picea abies Populus tremula Quercus robur Tilia cordata Acerplatanoides Alnus incana |
20 |
20 2 1 |
5 8 |
40 5 5 |
30 20 10 2 |
40 10 2 |
5 3 1 1 |
30 2 |
3 2 10 3 3 |
10 25 20 4 |
3 30 18 20 + |
5 12 |
20 30 40 10 |
40 15 30 5 |
||
|
Подрост и подлесок, покрытие, % |
40 |
30 |
50 |
15 |
30 |
70 |
45 |
50 |
40 |
50 |
65 |
25 |
50 |
40 |
30 |
40 |
|
Подрост деревьев, % Picea abies Quercus robur Populus tremula |
25 15 10 |
10 4 2 4 |
5 3 3 |
10 7 2 |
5 5 1 1 |
40 40 1 + |
35 32 3 + |
40 35 1 |
30 30 1 |
20 20 1 |
5 1 1 1 |
8 2 2 1 |
10 + 2 10 |
10 5 1 1 |
30 28 |
40 40 |
|
Tilia cordata |
+ |
4 |
2 |
|||||||||||||
|
Acerplatanoides |
2 |
1 |
5 |
1 |
3 |
+ |
||||||||||
|
Betulapendula |
+ |
+ |
8 |
5 |
||||||||||||
|
Подлесок, покрытие, % Corylus avellana Sorbus aucuparia |
15 5 2 |
30 30 1 |
50 50 5 |
10 8 2 |
30 20 + |
45 45 2 |
15 15 2 |
30 20 2 |
15 15 |
30 25 5 |
60 60 1 |
25 25 + |
40 40 + |
30 30 5 |
+ |
5 5 |
|
Viburnum opulus |
2 |
1 |
+ |
+ |
+ |
+ |
1 |
+ |
+ |
+ |
||||||
|
Salix caprea |
8 |
8 |
||||||||||||||
|
Lonicera xylosteum |
3 |
1 |
10 |
1 |
1 |
1 |
1 |
+ |
1 |
|||||||
|
Frangula alnus Padus avium Sambucus racemosa Malus sylvestris |
1 |
+ |
+ |
3 1 |
1 |
+ + |
+ 1 |
|||||||||
|
Травяной ярус, проективное покрытие, % |
70 |
85 |
60 |
50 |
85 |
80 |
40 |
70 |
60 |
90 |
60 |
65 |
80 |
50 |
85 |
60 |
|
Ajuga reptans Luzulapilosa Ranunculus cassubicus Deschampsia cespitosa |
1 8 10 |
30 1 10 |
2 1 2 |
2 1 3 |
2 1 3 + |
5 3 3 |
1 1 + 1 |
1 10 |
6 1 1 |
+ + + 1 |
5 1 |
2 1 2 + |
+ 2 + |
+ 10 1 |
1 + |
+ 1 1 |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
|
|
Dryopteris carthusiana |
5 |
5 |
+ |
7 |
5 |
2 |
3 |
3 |
+ |
1 |
+ |
1 |
||||
|
Athyrium filix-femina |
30 |
30 |
2 |
20 |
5 |
5 |
3 |
25 |
10 |
20 |
||||||
|
Galeobdolon luteum |
20 |
10 |
15 |
55 |
50 |
15 |
10 |
10 |
10 |
5 |
15 |
20 |
10 |
40 |
||
|
Stellaria holostea |
8 |
20 |
20 |
3 |
8 |
1 |
3 |
1 |
1 |
+ |
1 |
+ |
1 |
|||
|
Aegopodiumpodagraria |
5 |
1 |
3 |
15 |
50 |
1 |
5 |
5 |
5 |
65 |
7 |
|||||
|
Carexpilosa |
3 |
25 |
20 |
10 |
+ |
1 |
20 |
50 |
20 |
|||||||
|
Geum rivale |
2 |
20 |
3 |
1 |
+ |
3 |
1 |
1 |
1 |
1 |
||||||
|
Oxalis acetosella |
4 |
10 |
20 |
2 |
||||||||||||
|
Rubus saxatilis |
10 |
3 |
+ |
1 |
+ |
+ |
1 |
+ |
+ |
|||||||
|
Pulmonaria obscura |
1 |
7 |
5 |
1 |
20 |
5 |
10 |
1 |
7 |
|||||||
|
Asarum europaeum |
2 |
8 |
+ |
7 |
10 |
5 |
1 |
1 |
2 |
10 |
2 |
|||||
|
Convallaria majalis |
+ |
5 |
2 |
1 |
+ |
1 |
+ |
|||||||||
|
Crepispaludosa |
10 |
5 |
2 |
5 |
7 |
2 |
||||||||||
|
Dryopteris filix-mas |
2 |
2 |
1 |
1 |
1 |
+ |
1 |
3 |
1 |
|||||||
|
Equisetumpratense |
2 |
5 |
3 |
2 |
+ |
1 |
||||||||||
|
Equisetum sylvaticum |
2 |
1 |
1 |
1 |
2 |
3 |
+ |
3 |
||||||||
|
Filipendula ulmaria |
3 |
1 |
+ |
1 |
+ |
|||||||||||
|
Fragaria vesca |
3 |
10 |
1 |
1 |
2 |
5 |
2 |
+ |
1 |
+ |
+ |
|||||
|
Gymnocarpium dryopteris |
1 |
2 |
2 |
|||||||||||||
|
Lathyrus vernus |
1 |
1 |
+ |
1 |
1 |
3 |
1 |
|||||||||
|
Lysimachia nummularia |
+ |
1 |
+ |
+ |
||||||||||||
|
Lysimachia vulgaris |
1 |
2 |
1 |
+ |
+ |
1 |
1 |
+ |
+ |
|||||||
|
Urtica dioica |
1 |
+ |
3 |
1 |
||||||||||||
|
Maianthemum bifolium |
+ |
2 |
2 |
1 |
+ |
|||||||||||
|
Melica nutans |
2 |
2 |
+ |
+ |
||||||||||||
|
Milium effusum |
4 |
1 |
+ |
1 |
||||||||||||
|
Parisquadrifolia |
1 |
1 |
1 |
+ |
1 |
1 |
+ |
+ |
1 |
+ |
||||||
|
Viola mirabilis |
1 |
+ |
1 |
+ |
1 |
|||||||||||
|
Poa nemoralis |
+ |
15 |
1 |
|||||||||||||
|
Prunella vulgaris |
1 |
1 |
+ |
|||||||||||||
|
Ranunculus repens |
2 |
1 |
1 |
+ |
||||||||||||
|
Rubus idaeus |
1 |
8 |
+ |
|||||||||||||
|
Scrophularia nodosa |
+ |
+ |
+ |
+ |
||||||||||||
|
Solidago virgaurea |
+ |
+ |
+ |
+ |
||||||||||||
|
Stachys officinalis |
1 |
1 |
+ |
3 |
+ |
+ |
||||||||||
|
Angelica sylvestris |
1 |
+ |
+ |
2 |
1 |
|||||||||||
|
Thalictrum aquilegifolium |
+ |
1 |
+ |
+ |
||||||||||||
|
Euonymus verrucosa |
+ |
1 |
+ |
|||||||||||||
|
Geranium sylvaticum |
+ |
1 |
+ |
|||||||||||||
|
Trientalis europaea Brachypodium sylvaticum Veronica chamaedrys |
1 |
1 |
+ |
+ |
1 |
+ |
+ |
|||||||||
|
Pyrola rotundifolia Taraxacum officinale Solanum dulcamara Mercurialisperennis |
+ |
+ |
1 |
+ |
+ |
+ |
3 |
15 |
||||||||
|
Galium odoratum |
+ |
7 |
||||||||||||||
|
Aconitum septentrionale |
1 |
1 |
||||||||||||||
|
Моховой ярус, проективное покрытие, % |
10 |
10 |
5 |
30 |
7 |
10 |
10 |
5 |
6 |
4 |
7 |
3 |
7 |
15 |
0 |
10 |
|
Atrichum undulatum |
2 |
2 |
3 |
10 |
5 |
3 |
3 |
3 |
2 |
4 |
7 |
5 |
||||
|
Cirriphyllumpiliferum |
1 |
1 |
1 |
2 |
1 |
1 |
2 |
Окончание таблицы
1 2
Rhytidiadelphus triquetrus Brachythecium rutabulum Brachythecium salebrosum Callicladium haldanianum Eurhynchium angustirete Oxyrrhynchium hians Plagiochilaporelloides Plagiomnium affine Plagiomnium cuspidatum Plagiomnium undulatum Pleurozium schreberi Sciuro-hypnum reflexum Stereodonpallescens
+ 2
+
1 +
Прим. Субассоциации: 1 – caricosumpilosae ; 2 – typicum ; 3 – aegopodiosum ; 4 – athyriosum ; 5 – typicum . Варианты: 1а – Geum rivale ; 3а – Oxalis acetosella ; 4а – Pulmonaria obscura ; 4b – Carex pilosa ; 4с – Galeobdolon luteum ; 5а – Aegopodiumpodagraria .
На пробных площадях единично встречены виды: (приведен порядковый номер описания в таблице и проективное покрытие вида): Pinus sylvestris – № 9 (5); Salix caprea – № 10 (25); Fraxinus excelsior – № 15 (1); Euonymus verrucosa – № 16 (1); Juncus effusus – № 10 (1); Agrostis canina –№ 1 (5); Moehringia trinervia – № 8 (3); Carexpallescens – № 10 (+); Scirpus sylvaticus – № 8 (1); Alchemilla filicaulis – № 1 (4); Veronica officinalis – № 6 (+); Galium uliginosum – № 5 (+); Vicia sepium – № 6 (1); Viola canina – № 10 (+); Stellaria nemorum – № 9 (2); Myosotispalustris № 1 – (+); Festuca gigantea – № 8 (2); Valeriana officinalis № 1 (1); Galeopsis bifida – № 8 (1); Platanthera bifolia – № 1 (+); Hieracium umbellatum – № 1 (+); Hypericumperforatum – № 1 (1); Impatiens noli-tangere – № 8 (+); Cirsium heterophyllum – № 2 (3); Coccyganthe flos-cuculi – № 10 (+); Equisetum arvense – № 10 (+); Carex loliacea – № 9 (1); Orthilia secunda – № 1 (1); Anthriscus sylvestris – № 1 (1); Succisapratensis – № 6 (+); Dactylisglomerata – № 10 (1); Dryopteris expansa – № 11 (1); Glechoma hederacea – № 15 (10); Neottia nidus-avis – № 12 (+); Calamagrostis arundinacea – № 14 (1); Stachys sylvatica – № 16 (1); Ranunculus acris – № 14 (+); Carex digitata – № 16 (1); Carex sylvatica – № 13 (+); Polygonatum odoratum – № 15 (+); Pylaisiapolyantha – № 2 (+); Rhytidiadelphus squarrosus – № 10 (+); Brachythecium rivulare – № 16 (+); Dicranella heteromala – № 1 (1); Dicranumpolysetum – № 8 (3); Dicranum scoparium – № 1 (1); Ditrichum flexicaule – № 6 (+); Sanionia uncinata – № 1 (1); Fissidens bryoides – № 14 (1); Fissidens exilis – № 7 (1); Plagiomnium ellipticum – № 10 (+); Plagiomnium medium – № 10 (+); Lophocolea heterophylla – № 1 (1); Rhytidiadelphus subpinnatus – № 10 (+)
Группа ассоциаций Betuleta nemoriher-bosa compositae – березняки неморальнотравные сложные
Сообщества группы ассоциаций отличаются от сообществ предыдущей группы присутствием в древостое и подросте широколиственных пород (дуба и липы). Отметим, что значительную роль здесь начинает играть осина, как порода более требовательная к богатству почв. Средняя сомкнутость древостоя 0,7. Флористический состав травяного яруса схож с сообществами простых березняков неморальнотравных. В.И. Василевич (1996) для Северо-Запада Европейской России также приводит ассоциацию «березняк сложный» (под названием Corylo-Betuletum) и отмечает, что «высокое обилие широколиственных древесных пород не приводит к появ- лению травяного яруса, заметно отличающегося от того, что имеется при отсутствии этих видов» (Василевич, 1996: 11). Мы подразделяем группу ассоциаций березняки неморальнотравные сложные на 2 ассоциации по наличию в древостое одной из двух широколиственных пород – дуба или липы. Но различия между ассоциациями этим не ограничиваются.
Ассоциация 3. Betuletum quercoso-corylosum – березняк с дубом лещиновый. Средняя сомкнутость древостоя составляет 0,58. В древостое, помимо березы и дуба, часто встречаются Picea abies, Populus tremula, Tilia cordata, Acer platanoides. Сообщества ассоциации характеризуются наличием дуба в древостое, хорошо выраженным подлеском из лещины и крайне слабым участием ели в подросте. Сообщества ассоциации отличаются также постоянным участием и высоким (до 25%) проективным покрытием кочедыжника женского. С высоким постоянством встречаются также Equisetum pratense, Crepis palu-dosa. В пределах этой ассоциации мы выделяем одну субассоциацию и три варианта.
Субасс. athyriosum – кочедыжниковая. В травяно-кустарничковом ярусе помимо Athy-rium filix-femina встречаются те же виды, что и в других сообществах группы неморальнотравных березняков.
Var. Pulmonaria obscura – вариант с медуницей. У Н.А. Коновалова (1929) для Подмосковных опытных лесничеств также выделена ассоциация с медуницей ( Betuletum pulmonariosum ), однако она значительно отличается от нашего. В сообществах ассоциации, описанной Н.А. Коноваловым (1929) в древесном ярусе отсутствует примесь широколиственных пород, нет густого подлеска из лещины, зато присутствуют другие виды-доминанты, относящиеся к неморальным элементам флоры.
Var . Carex pilosa – вариант волосистоосоковый – выделен по содоминированию кочедыжника и осоки.
Var . Galeobdolon luteum – вариант зе-ленчуковый.
Ассоциация 4. Betuletum tilioso-caricosum pilosae – березняк с липой волосистоосоковый. Средняя сомкнутость древостоя в этой ассоциации гораздо выше, чем в предыдущей, достигая 0,93. По нашим данным, верхний ярус образован исключительно Betula pubescens. Betula pendula в сообществах ассоциации не отмечена. Также в сложении древостоя участвуют Picea abies, Populus tremula, Acer platanoides. В сообществах ассоциации практически полностью отсутствует ярус из лещины, которая в лесах с участием липы в древостое обычно встречается редкими кустами (Курнаев, 1968). При этом, в подросте отмечено значительное участие ели. Таким образом, вполне вероятно, что подобные сообщества без вмешательства человека в скором времени сменятся липово-еловыми лесами. В сообществах ассоциации отсутствует Athyrium filix-femina, что происходит, вероятно, вследствие затенения сомкнутым древостоем. Также интересно, что только в березняках с липой присутствует ясменник душистый (Galium odoratum), в других неморальнотравных березняках он не был встречен. В.И. Василевич (1996) также отмечает, что ясменник встречается только в тех березняках, где имеется липовый ярус. Пролесник (Mercurialis perennis) и борец северный (Aconitum septentrionale) также встречены только в березняках с липой.
Субасс. typicum – типичная.
Var. Aegopodium podagraria – вариант снытевый. Выделяется по преобладанию сныти в травяном ярусе.
С помощью неметрического многомерного шкалирования (NMDS) проанализировано положение сообществ выделенных ассоциаций в многомерном пространстве переменных. Выяснилось, что сообщества каждой ассоциации создают неперекрывающиеся кластеры и, таким образом, хорошо дифференцируются в экологическом пространстве (рис. 2).
Можно предположить, что комплексная ось 2 представляет собой степень неморали-зации сообществ, поскольку в нижней половине ординационной диаграммы представлены сообщества со значительных участием широколиственных пород, а в верхней – они практически отсутствуют. По оси 1 наблюдается изменение доли участия нитрофильных видов. Так, в левой части диаграммы располагаются сообщества с бóльшим участием нит-рофильных видов.
При преобразовании многомерного пространства в двумерное относительное расположение точек изменяется. Чтобы оценить, значимы ли эти изменения по отношению к исходному расположению описаний, применяли показатель стресса. В данном случае, при сведении многомерного пространства к двумерному уровень стресса составил 14%. Обычно считается, что при значениях стресса в пределах от 10 до 20% результаты ордина- ции можно достоверно интерпретировать, хотя некоторые объекты и могут располагаться случайно (Clarke, 1993). Таким образом, в данном случае вполне можно доверять полученному результату. Как показывает практика, чем больше описаний и, соответственно, видов используется при ординации, тем выше показатель стресса. Следует отметить, что низкий показатель стресса в результате проведенной ординации, возможно, является также следствием небольшой выборки (Clarke, 1993; Новаковский, 2008; McCune, Grace, 2002).
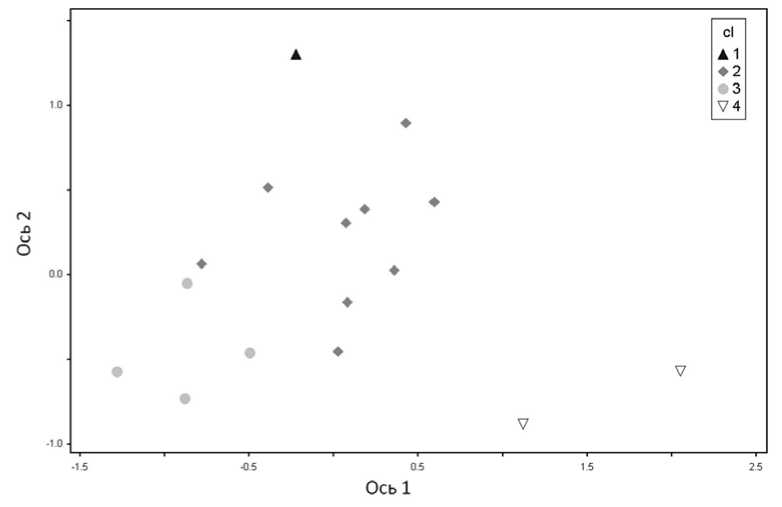
Рис. 2. Ординация геоботанических описаний лесных сообществ методом NMDS. Ассоциации: 1 – Betuletum athyriosum ; 2 – Betuletum galeobdolosum ; 3 – Betuletum quercoso-corylosum ;
4 – Betuletum tilioso- caricosum pilosae .
Ordination of phytocenological descriptions of forest communities of the method of NMDS
Заключение
Рассмотрено ценотическое разнообразие неморальнотравных березняков в западной части Наро-Фоминского района Московской области. На основании эколого-фитоценотической классификации, проведенной методом табличного анализа, выделено 2 группы ассоциаций, 4 ассоциации, 5 субассоциаций и 6 вариантов. Проанализированы особенности видового состава и структуры выделенных синтаксонов.
В группе ассоциаций березняков неморальнотравных выделено 2 ассоциации – Betuletum athyriosum и Betuletum galeob-dolosum. Первая ассоциация (березняк кочедыжниковый) отличается менее сомкнутым древостоем, отсутствием выраженного подлеска, доминированием в травяном ярусе Athyrium filix-femina и отсутствием содоми- нантов. Во второй ассоциации (березняк зе-ленчуковый) наблюдается густой подлесок из лещины, более сомкнутый древесный полог, в травяном ярусе характерно высокое постоянство и высокое покрытие зеленчука. В группе ассоциаций березняков неморальнотравных сложных выделено также 2 ассоциации – Betuletum quercoso-corylosum и Betuletum tilioso-caricosum pilosae. Первая (березняк с дубом лещиновый) отличается хорошо выраженным подлеском из лещины, менее сомкнутым древостоем, высоким обилием и постоянством Athyrium filix-femina. Вторая (березняк с липой волосистоосоковый) характеризуется высокой сомкнутостью древостоя, отсутствием подлеска и значительным участием Picea abies в подросте. Кроме того, выявлены виды, встречающиеся только в сообществах с липой – Mercurialis perennis, Galium odoratum и Aconitum septentrionale.
Анализ расположения геоботанических описаний в ординационном пространстве показал, что сообщества хорошо дифференцируются и имеют неперекрывающиеся экологические ареалы. Таким образом, выделенные ассоциации действительно различаются по экологии. Выявлены следующие закономерности в расположении сообществ в орди-национном пространстве: по оси 2 – измене-
Список литературы Фитоценотическое разнообразие березовых неморальнотравных лесов юго-западного Подмосковья
- Абатуров Ю.Д., Зворыкина К.В., Ильюшенко А.Ф. Типы березовых лесов центральной части южной тайги. М.: Наука, 1982, 155 с.
- Абесадзе Т.К. Видовой состав березовых лесов Подмосковья (на примере севера московской Мещеры). Вопр. лесовыращивания и рационального лесоиспользования. Науч. тр. Московск. Лесотехнич. ин-та, вып. 148, 1983, c. 26-29.
- Анненская Г.Н., Жучкова В.К., Калинина В.Р., Мамай И.И., Низовцев В.А., Хрусталева М.А., Цесельчук Ю.Н. Ландшафты Московской области и их современное состояние. Смоленск: Изд-во Смоленск. гуманитар. ун-та, 1997, 296 с.
- Беляева Н.Г., Попов С.Ю. Изменение лесистости бывшего Верейского уезда Московской области за последние 200 лет. Лесоведение, 2016, № 1, c. 44-54.
- Биологическая флора Московской области. Вып. 7. Под ред. Т.А. Работнова. М.: Изд-во Моск. ун-та, 1983, 263 с.
