Фитопланктон высокоминерализованных рек Приэльтонья

Автор: Горохова Ольга Геннадьевна, Зинченко Татьяна Дмитриевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 5-5 т.16, 2014 года.
Бесплатный доступ
Представлены результаты исследований фитопланктона семи высокоминерализованных рек Приэльтонья (2008, 2012-14 гг.). Выявлено 116 таксонов водорослей из 7 систематических отделов, дана характеристика таксономической структуры. Диатомовые водоросли (Bacillariophyta) являются ведущими по видовому богатству и количественному развитию. Наибольшая численность и биомасса фитопланктона регистрировались в устьевых участках рек. Выявлены массовые виды ( Chaetoceros muelleri Lemm., Nitzschia closterium Ehr., Amphora coffeaeformis (Ag.) Kütz., Achnanthes brevipes Ag., виды родов Thalassiosira, Cyclotella, Dunaliella ), принадлежащие к широко распространенным эвригалинным таксонам. Изменение численности и биомассы фитопланктона не зависит от уровня минерализации в реках, удельное видовое богатство отрицательно коррелирует с величиной солености. Отмечено изменение структурных характеристик альгоценозов с выявлением монодоминантных сообществ в диапазоне минерализации от 6,6 до 41,4 г/л.
Соленые реки, оз. эльтон, фитопланктон, численность, биомасса
Короткий адрес: https://sciup.org/148203457
IDR: 148203457 | УДК: 582.574.2
Phytoplankton of saline rivers of the lake Elton basin
Is given the results of studies of phytoplankton seven highly mineralized rivers of the lake Elton basin in 2008 and 2012-14. Identified 116 taxa of algae of 7 systematic divisions, characterized taxonomic structure. Species richness and the level of quantitative development determined mainly Bacillariophyta. Maximum values of the abundance of phytoplankton are often formed in the estuaries of rivers. Revealed abundant species ( Chaetoceros muelleri Lemm., Nitzschia closterium Ehr., species of the genera Thalassiosira, Cyclotella, Dunaliella, Amphora coffeaeformis (Ag.) Kütz., Achnanthes brevipes ), belonging to the widespread euryhaline species. The change in the abundance and biomass of phytoplankton does not depend on the level of salinity in rivers, specific species richness was negatively correlated with the magnitude of salinity. A noticeable change in the structural characteristics of the communities identify monodominant communities in the salinity range of 6,6 to 41.4 g / L.
Текст научной статьи Фитопланктон высокоминерализованных рек Приэльтонья
П риродно-территориальный комплекс При-эльтонье в аридной зоне Прикаспийской низменности представляет собой феномен ландшафта солянокупольной тектоники в мало нарушенном массиве опустыненных степей [14]. В настоящее время он имеет статус природного парка «Эльтонский», который решает проблему сохранения ценных комплексов и объектов, обеспечивая возможность оптимизации природопользования в условиях антропогенного воздействия на территории Приэльтонья [6].
Изучение фитопланктона рек, впадающих в оз. Эльтон, осуществляется в рамках комплексных многолетних исследований экологического состояния водных объектов Приэльтонья, оценки биоразнообразия гидробионтов и структуры их сообществ в условиях значительного градиента минерализации [8, 9].
Первые гидробиологические изыскания, проведенные в окрестностях оз. Эльтон, относятся к началу ХХ в. В работе «О микрофауне водоемов окрестностей Эльтона и Баскунчака» [3] содержатся фаунистические данные, однако авторы упоминают и о двух видах фитофлагеллят: Dunaliella salina Teod. и Asteromonas gracilis Artari. Эти водоросли были встречены в самом оз. Эльтон («у устьев рек Хара и Большая Само-рода» [3], стр. 58), D. salina была найдена в реках Малая и Б. Саморода (ранее Б.Сморогда). В дальнейшем (1998 и 2001 гг.) фитопланктон рек Приэльтонья изучали сотрудники Волгоградско-
го отделения ГосНИОРХ [13], которые приводят данные о таксономическом составе, преобладающих видах, а также количественном развитии фитопланктона. Основу видового богатства повсеместно формировали диатомовые водоросли, тогда как представители других отделов значительно уступали им по числу видов. Структурообразующие виды были выделены авторами по вкладу в численность/биомассу фитопланктона и по частоте встречаемости. К их числу в различных реках относились представители отдела Ba-cillariophyta – Cyclotella meneghiniana Kütz., Chaetoceros muelleri Lemm., Achnanthes brevipes Ag., Navicula sp. и отдела Cryptophyta – Cryptomonas erosa Ehr., C. ovata Ehr., Rhodomonas sp. На отдельных участках р. Хара авторы отмечали массовое развитие видов Euglenophyta – Eu-glena texta (Duj.) Hubner, E. polymorpha Dang.
Сведения о современном состоянии фитопланктона рр. Хара, Ланцуг, М. Саморода, Солянка, приводятся в публикациях Т.Н. Бурковой, обработавшей предоставленный ей материал, собранный сотрудниками лаборатории экологии малых рек в августе 2006, 2008-2011 гг. и в мае 2011 г. Автор приводит данные о таксономическом составе альгофлоры планктона рек, указывая на ведущую роль диатомовых водорослей (32-42% видового состава), а также цианопрокариот и зеленых водорослей (20-22%). Список аль-гофлоры приводится для р. Хара, он включает 219 видов и внутривидовых таксонов [4,5].
Сезонная, годовая и пространственная изменчивость таксономического состава альгофлоры в условиях значительного градиента минерализации требует проведения дальнейших исследований и детального анализа количественных и структурных характеристик фитопланктона.
МАТЕРИАЛЫ И МЕТОДЫ
Материал представлен пробами фитопланктона, отобранными 12-14 августа 2008 г., 17-19 мая и 13-15 августа 2012 г., 13-15 августа 2013 г. 1217 мая и 13-17 августа 2014 г. на участках среднего течения и в устьях рек Хара, Ланцуг, Чернавка, Солянка, Б. Саморода, М. Саморода и Карантин-ка. На участках верхнего течения рек, в связи с отсутствием постоянного стока, отбор проб осуществлялся в отдельные периоды. Сбор и обработка проб для определения количественных показателей фитопланктона проведены по общепринятым гидробиологическим методикам [10]. Пробы, зафиксированные раствором Люголя, концентрировали фильтрацией через мембранные фильтры с диаметром пор около 1 мкм с применением вакуумного насоса. Определение, подсчет и измерение водорослей проводили в камере типа «Учинская», объемом 0,01 мл. Биомасса фитопланктона вычислена счетно-объемным методом. К доминирующим отнесены виды, численность или биомасса которых составляла не менее 10% от общей величины, к субдоминантам - виды с численностью (биомассой) равной от 5 до 10%. Всего было собрано и обработано более 80 альго-логических проб планктона. Для характеристики сообществ использовали показатели: общее видовое богатство (число видов в альгофлоре), удельное видовое богатство (число видов в пробе), численность (млн кл/л), биомассу (мг/л), индексы видового разнообразия Шеннона (H), и вырав-ненности Пиелу (Е). Под частотой доминирования понимали количество проб, в которых вид доминирует (в процентах от общего для реки числа проб). Рассчитывали коэффициент корреляции Пирсона (р < 0,05).
Питание соленых рек, впадающих в оз. Эльтон происходит за счет подземных вод и атмосферных осадков. Для рек характерны сезонные колебания уровня воды: весной после таяния снега они наполняются, в летний период глубины уменьшаются. Верховья рек обычно пересыхают летом, а наполнение водой отмечается в русловых участках среднего и нижнего течения. Гидрологогидрохимический режим р. Хара отличается тем, что в период весеннего половодья в нее осуществляется попуск волжских вод через систему каналов. Прогрев воды в реках происходит быстро: так, во второй декаде мая 2012 и 2014 гг. температура воды достигала 14,8-22,5 и 19,2-33,4 °С соответственно, и оставалась в пределах указанных величин в летне-осенний период. Скорость течения изменяется от 0,2 до 0,4 м/сек. Величина рН характеризует воды рек как слабощелочные и щелочные (6,9-9,2). Реки Приэльтонья в разной степени загрязняются в результате антропогенного воздействия, а в приустьевой части также по причине концентрации гнездящихся водоплавающих и околоводных птиц [8, 9].
По минерализации, в соответствии с классификации О.А. Алекина [2], реки-притоки оз. Эльтон можно разделить на 2 группы: солоноватые (с минерализацией от 1 до 25 г/л), к ним относятся рр. Хара, Ланцуг и Б. Саморода и реки с морской соленостью (25-50 г/л) - Чернавка и Солянка. В первой группе рек минерализация увеличивается от верховьев к устью, во второй диапазон изменения солености по длине реки меньше (табл. 1). Преобладающими анионами в реках Чернавка, Солянка, М. Саморода являются ионы хлора; в реках Хара, Ланцуг и Б. Саморода содержание ионов хлора и сульфатных ионов изменяется по длине реки [12]. По преобладающим катионам вода в реках относится к натриевой, натриймагниевой или магниевой группе. Концентрации основных биогенных элементов (минерального азота и общего фосфора) в реках высокие, отмечена обратная зависимость содержания общего фосфора от уровня минерализации [12]. Концентрации кремния были типичными для поверхностных вод Волжского бассейна [12]. В табл. 1 представлены некоторые гидрологогидрохимические характеристики рек Приэльто-нья в местах отбора проб.
Таблица 1. Гидролого-гидрохимические характеристики рек в местах отбора проб
|
Река |
Длина, км** |
Глубина, м* |
Температура, °С* |
Минерализация, г/л*** |
||
|
верхнее течение |
среднее течение |
нижнее течение (устье) |
||||
|
Чернавка |
5,2 |
0,15-0,75 |
18,5-31,5 |
29,6 28,1-31,0 |
28,5 27,5-30,0 |
28,1 17,2-32,0 |
|
Солянка |
6,7 |
0,05-0,80 |
19,2-30,2 |
27,10 25,7-28,6 |
26,10 24,0-27,6 |
27,20 25,2-29,0 |
|
Хара |
46,4 |
0,03-0,65 |
16,1-33,4 |
11,5 |
8,4 6,6-10,0 |
18,7 11,7-41,4 |
|
Ланцуг |
19,9 |
0,10-0,45 |
14,8-30,1 |
- |
6,3 5,9-7,0 |
15,8 10,6-30,0 |
|
Б. Саморода |
24,3 |
0,02-0,60 |
19,2-26,5 |
- |
9,0 7,6-9,8 |
11,5 10,0-15,0 |
|
М. Саморода |
10,3* |
0,05-0,25 |
24,3-25,8 |
- |
- |
180,3 |
|
Карантинка |
2,4* |
0,01-0,05 |
17,6-28,1 |
- |
9,3 |
- |
* измерения проведены в период сбора альгологических проб; **расчетные данные выполнены Е.М. Белозеровой (каф. гидрологии суши МГУ); рр. М. Саморода и Карантинка – по [6]; *** данные аккредитованной гидрохимической лаборатории ООО «Центр мониторинга водной и геологической среды» г. Самара; над чертой – средние значения, под чертой – диапазон.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
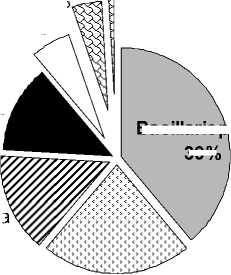
Таксономический состав. Фитопланктон соленых рек представлен 116 таксонами водорослей, относящихся к 7 систематическим отделам (табл. 2). Основу альгофлоры формировали виды и разновидности Bacillariophyta (диатомовые) – 39% от общего видового списка (табл. 2, рис. 1); их доля в отдельных реках составляла от 27 (М. Саморода) до 61% (Ланцуг). В таксономическом отношении диатомовые водоросли – одна из основных групп планктона речных систем. Их значимая роль в соленых реках была сопоставима с долей в альгофлоре континентальных минерализованных водоемов [17, 20, 22].
Cyanoprokaryota (цианопрокариоты) составляли 22% альгофлоры планктона рек, доля их в разных водотоках меняется от 14 до 27% (рис. 1). Видовое богатство этой группы связано с толерантностью некоторых представителей к условиям повышенной минерализации [17, 19]. Кроме того, обилие видов этих двух отделов в неглубоких соленых реках объясняется особенностями проникновения в водную толщу типичных обитателей придонного горизонта в условиях высокой плотности соленых вод.
Таблица 2. Таксономический состав водорослей фитопланктона соленых рек, впадающих в оз. Эльтон (n=80)
|
Отделы |
Хара |
Большая Саморода |
Ланцуг |
Чернавка |
Солянка |
Карантинка |
Малая Саморода |
Всего |
|
Cyanoprokaryota |
14 |
13 |
7 |
7 |
9 |
4 |
3 |
26 |
|
Chrysophyta |
1 |
1 |
||||||
|
Bacillariophyta |
27 |
30 |
32 |
16 |
16 |
6 |
3 |
45 |
|
Dinophyta |
4 |
1 |
1 |
5 |
||||
|
Cryptophyta |
7 |
5 |
6 |
5 |
4 |
1 |
7 |
|
|
Euglenophyta |
12 |
5 |
2 |
3 |
3 |
3 |
15 |
|
|
Chlorophyta |
13 |
6 |
3 |
5 |
3 |
1 |
5 |
17 |
|
Всего |
78 |
60 |
51 |
36 |
35 |
15 |
11 |
116 |
Cryptophyta 6%
Dinophyta 4%
й
Chrysophyta 1%
Chlorophyta 15%
Euglenophyta 13%
39%
Cyanoprokaryota 22%
Bacillariophyta
А
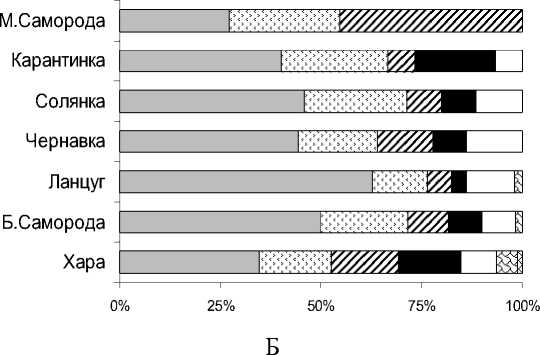
Рис. 1. Соотношение видов водорослей разных отделов в альгофлоре планктона соленых рек: А - общая характеристика; Б – в реках
Виды отдела Chlorophyta занимают третье-четвертое место по значению в формировании альгопланктона соленых рек Приэльтонья (рис. 1, табл. 2). В составе зеленых водорослей встречается не более 3-13 видов в реках, независимо от уровня их минерализации. Они представлены в первую очередь формами, способными выдерживать значительные концентрации солей: фитоф-лагеллятами из родов Dunaliella, Tetraselmis, Asteromonas. Более 50% видов зеленых водорослей (порядка Chlorococcales) характерны для планктона солоноватой р. Хара.
Эвгленовые водоросли (15 видов) в планктоне отдельных рек представлены единичными видами (табл. 2). В отличие от пресных водоемов, где условия развития для этой группы водорослей обычно благоприятны, в высокоминерализованных реках Euglenophyta наиболее чувствительны к воздействию экстремальных факторов.
Криптофитовые водоросли в реках-притоках оз. Эльтон формируют около 6% видового состава, выделяясь относительным разнообразием в планктоне солоноватых рр. Ланцуг, Хара и Б. Саморода. В отличие от других групп водорос- лей, представители отдела динофитовых встречены эпизодически в трех реках (табл. 2), а золотистые (Mallomonas sp.) единичны в среднем течение р. Хара. Наибольшим видовым богатством в период исследований (2008, 2012-2014 гг.) характеризовался фитопланктон солоноватой р. Хара (табл. 2).
Структурные и количественные характеристики альгоценозов планктона.
В структуре альгоценозов высокоминерализованных рек количественное развитие фитопланктона определяли Bacillariophyta с преобладанием их типично планктонных форм: Chaetoceros muel-leri Lemm., Nitzschia closterium Ehr., Thalassiosira weissflogii (Grun.) G.Fryxell & Hasle, Thalassiosira sp., Cyclotella meneghiniana Kütz., Cyclotella sp. В тоже время в планктоне регистрировались и представители бентоса, такие как: Amphora cof-feaeformis (Ag.) Kütz., Navicula pygmaea Kütz.,
Navicula sp., Achnanthes brevipes Ag. и некоторые другие. Постоянное преобладание диатомовых отмечено в рр. Ланцуг, Чернавка и Солянка. При близком составе массовых форм, комплексы доминирующих видов Bacillariophyta в каждой из рек имели некоторые индивидуальные черты. Например, частота доминирования некоторых из названных видов выше в соленых реках по сравнению с солоноватыми (табл. 3). Кроме того, отдельные виды Bacillariophyta, не всегда доминируют в каждой из соленых рек. Например, вид Nitzschia closterium в рр. Хара и Ланцуг развивался в небольшом количестве. Вид Chaetoceros muelleri в р. Б. Саморода отмечен единично, а в р. Солянка нами не зарегистрирован. Подробная характеристика массовых видов Bacillariophyta в структуре планктоценозов солоноватых и соленых рек представлена в табл. 3.
Таблица 3. Характеристики основных видов доминантов и субдоминантов (d, sd) фитопланктона в солоноватых (рр. Хара, Ланцуг) и соленых реках (Чернавка, Солянка)
|
Виды |
Реки |
Численность |
Биомасса |
частота доминирования |
||
|
млн кл./л |
% от общей |
мг/л |
% от общей |
|||
|
Chaetoceros muelleri (d, sd) |
I* |
0,6-46,5 |
12-99 |
1,0-14,7 |
6-98 |
33 (28)*** |
|
II |
2,6-19,5 |
30-98 |
0,6-3,15 |
30-85 |
33 (33) |
|
|
III |
0,5-96,0 |
5-59 |
0,24-22,31 |
7-72 |
38 (38) |
|
|
Thalassiosira weissflogii (d) |
I |
0,01-0,49 |
20-43 |
0,01-1,38 |
11-74 |
20 (28) |
|
II |
–** |
– |
0,07-0,18 |
10-13 |
- (22) |
|
|
III |
0,68 |
11 |
2,33-4,41 |
30-63 |
10 (20) |
|
|
IV |
0,3-5,5 |
15-28 |
0,11-19,31 |
35-72 |
33 (67) |
|
|
Nitzschia closterium (d, sd) |
III |
1,1-63,7 |
6-67 |
0,14-8,09 |
8-53 |
56 (56) |
|
IV |
3,3-28,8 |
17-64 |
0,54-4,82 |
6-31 |
38 (38) |
|
|
Cyclotella sp. (d, sd) |
I |
2,92 |
28 |
0,36-1,01 |
6-10 |
7 (13) |
|
II |
0,6-239,4 |
55-100 |
0,07-27,53 |
30-100 |
50 (42) |
|
|
Cyclotella meneghiniana (d, sd) |
I |
– |
– |
0,07-0,99 |
18-25 |
- (13) |
|
II |
– |
– |
0,06-0,26 |
7-25 |
- (17) |
|
Из состава цианопрокариот к доминантам относятся в основном виды родов Oscillatoria и Lyngbya, попадающие в воду из цианобактериальных матов, где они обильно развиваются. Доминирование планктонных видов родов Merismopedia, Anabaenopsis, Planktothrix отмечено локально в верхнем и среднем течении и в застойных участках р. Хара при солености до 6-9 г/л. К числу массовых форм планктона в большинстве исследованных рек принадлежат и Cryptophyta (Cryptomonas salina Wisl., Cryptomonas cf. erosa и др.). Например, в р. Солянка криптофитовые являются постоянным структурным компонентом альгоценозов, входя в состав доминантов и субдоминантов (доминирование в 60% проб). Виды отделов Euglenophyta и Dinophyta доминировали в планктоне отдельных рек эпизодически. Несмотря на то, что присутствие представителей криптофитовых и эвгленовых водорослей в составе преобладающих форм планктона пресных водоемов имеет индикаторное значение и нередко указывает на загрязнение, причины их массового развития в соленых реках могут быть иными и требуют дальнейшего анализа. Для некоторых рек характерно локальное доминирование водорослей отдела Chlorophyta. В устьевых участках рр. Хара, Б. Саморода, а также в устье и в среднем течение р. М. Саморода было отмечено формирование олигодоминантных сообществ фитопланктона с преобладанием жгутиковых форм зеленых водорослей рода Dunaliella, разнообразие которых характерно для рек При-эльтонья [18]. Кроме того, в устьях этих рек были зарегистрированы и другие представители
Chlorophyta, относящиеся к пикопланктонной фракции с размерами клеток 1,4-2,3 мкм. Эта группа, при численности 73-80% от суммарной фитопланктона, создавала до 6-13% его биомассы. Отметим, что изучению автотрофного пикопланктона уделяется в настоящее время большое внимание: имея высокую фотосинтетическую активность, он является одним из существенных производителей первичной продукции и важнейшим компонентом пищевой цепи в водоемах [11, 21, 24]. В целом в состав массовых форм, опреде- ляющих структуру альгоценозов соленых рек, входят виды характерные для высокоминерализованных континентальных вод, эвригалобы, которые способны к осморегуляции [15, 17, 18, 22, 23].
В табл. 4 приведены основные характеристики фитопланктонных сообществ рек Приэльтонья. Значительный диапазон величин численности, биомассы, числа видов и показателей разнообразия был характерен для всех рек вне зависимости от их уровня минерализации.
Таблица 4. Основные структурные показатели фитопланктона рек Приэльтонья
|
Реки |
численность, млн кл./л |
биомасса, мг/л |
число видов в пробе |
индекс Шеннона, бит/экз. |
индекс Пиелу |
|
Хара |
0,16-161,15 |
0,128-26,41 |
3-29 |
0,07-2,38 |
0,02-0,56 |
|
Ланцуг |
0,45-239,98 |
0,23-28,22 |
1-19 |
0,001-3,09 |
0,001-0,82 |
|
Солянка |
0,49-45,09 |
0,263-26,70 |
5-20 |
1,54-3,22 |
0,45-0,92 |
|
Чернавка |
0,68-163,09 |
0,45-31,25 |
8-16 |
1,00-2,89 |
0,33-0,87 |
|
Б. Саморода |
0,88-13,24 |
0,43-16,31 |
4-26 |
1,14-3,77 |
0,15-0,76 |
|
М. Саморода |
7,94-165,22 |
2,71-122,50 |
3-6 |
0,33-1,26 |
0,17-0,84 |
|
Карантинка |
0,23-12,58 |
0,03-2,99 |
3-14 |
0,83-1,24 |
0,32-0,52 |
Сообщества фитопланктона соленых рек Чернавка и Солянка, как правило, характеризуются полидоминантностью и более высокой выравнен-ностью обилий видов, то есть диапазон величин биоценотических индексов невелик (табл. 4). Высокое видовое разнообразие отмечено в р. Б. Са-морода. В солоноватых реках Хара и Ланцуг, где формируются монодоминантные сообщества, значения индексов разнообразия и выравненности минимальны (табл. 4). Вклад одного-двух видов в суммарную численность фитопланктона составляет 85-99%.
Зависимость количественных показателей фитопланктона от солености рек. Многочисленные исследования фитопланктона континентальных водоемов свидетельствуют об уменьшении видового богатства и упрощении структуры альгофлоры в высокоминерализованных водах. Нередко отмечается массовое развитие отдельных видов в планктоценозах при увеличении солености в условиях, когда она является экстремальным фактором [1, 15, 16, 19, 23]. В тоже время приводятся сведения об обеднении видового состава организмов высших трофических уровней (зоопланктона, макрозообентоса) при воздействии экстремальных величин солености и отсутствии зависимости числа видов фитопланктона от уровня минерализации [7].
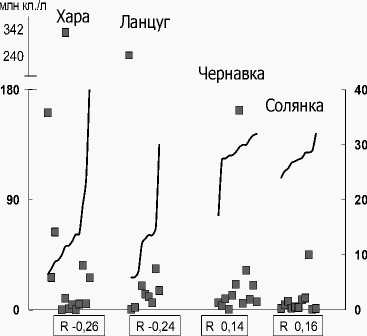
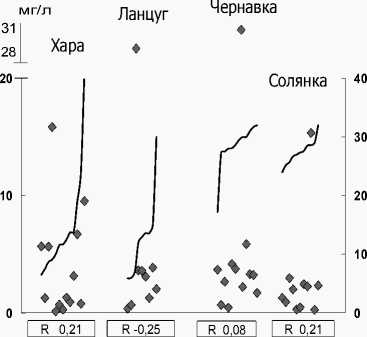
Нами проведена сравнительная оценка зависимости числа видов, численности и биомассы фитопланктона в солоноватых и соленых реках от уровня минерализации. Для удельного числа видов характерна слабая отрицательная корреляционная связь с ней в солоноватых реках с большим градиентом минерализации (6-41 г/л) Хара и Лан-цуг. В соленых реках Чернавка и Солянка с постоянно высокой минерализацией (26-32 г/л) вы- явлена значимая корреляционная связь (R -0,56, R -0.56) видового богатства с соленостью (рис. 1А). Что касается численности и биомассы, то зависимость их величин от уровня минерализации не выражена (рис. 2 Б, В). Например, в соленых реках Чернавка и Солянка минимум (0,39 и 0,09 млн кл./л) и максимум (160,9 и 32,6 соответственно) обилия доминирующих планктонных диатомей зарегистрированы при близких значениях солености (29 и 28 г/л). В р. М. Саморода, при максимальной величине минерализации (180 г/л), отмечено массовое развитие пикопланктонной фракции Chlorophyta, обусловившей максимальную численность фитопланктона (табл. 4). Для устьевого участка р. Хара самому высокому уровню солености (41‰) соответствовали средние для этого водотока численность и биомасса, но удельное число видов было низким (5 видов), а альгоценоз устья характеризовался доминированием Dunaliella sp., который определил 98% суммарной численности и биомассы и минимум видового разнообразия для реки (табл. 4).
Можно констатировать, что основные количественные характеристики фитопланктона высокоминерализованных рек Приэльтонья, в их естественном состоянии, не зависят от изменения минерализации. По всей видимости, её величины в пределах от 6 до 180 г/л, в соленых реках- притоках оз. Эльтон не являются критическими для развития фитопланктона. Как было показано выше, структурные особенности альгоценозов проявляются в формировании моно- и олигодоми-нантных сообществ в условиях максимальных для реки значений минерализации: в устьевых участках рек Ланцуг (соленость 30 г/л), Хара (41 г/л), Б. Саморода (180 г/л).
Хара
Чернавка Солянка
Ланцуг
7 89 R 101112 - 131 0 415 , 16 4 1718 2 1920212 2
9 3031 R 323334 - 35 0 3637 , 38 3 3940 3 414243 44
50 5152 R 535455 - 56 0 5758 , 59 5 6061 6 626364 6566676869 70 71 72 R 73 74 75 - 76 0 77 78 , 79 5 80 81 1 82838485 86
о Число видов □ Численность <0} Биомасса
Минерализация, г/л (по дополнительной оси)
Рис. 2. Распределение количественных характеристик фитопланктона солоноватых (Хара, Ланцуг) и соленых (Чернавка, Солянка) рек по градиенту минерализации
ЗАКЛЮЧЕНИЕ
Фитопланктон рек аридного региона Приэль-тонья представлен видами из 7 отделов (116 таксонов рангом ниже рода). Таксономическое бо- гатство определяют водоросли
Bacillariophyta→Cyanoprokaryota→Euglenophyta →Chlorophyta. Наибольшим видовым богатством отличается фитопланктон солоноватых рек Хара, Б. Саморода и Ланцуг. Количественные характе- ристики фитопланктонных сообществ соленых рек экосистемы оз. Эльтон изменяются в диапазоне 0,16-239,98 млн кл./л; 0,03-122,50 мг/л и обусловлены в основном развитием планктонных форм Bacillariophyta. Максимальные показатели численности и биомассы фитопланктона формируются преимущественно в устьевых участках рек. Массовые виды (Chaetoceros muelleri, Nitzschia closterium, Amphora coffeaeformis, Ach-nanthes brevipes, виды родов Thalassiosira, Dunaliella и др.) принадлежат к эвригалобам, способным к осморегуляции в условиях изменения солености. Зависимости количественных характеристик фитопланктона от величины изменения минерализации не выявлено. В тоже время, структурные особенности альгоценозов выражаются в снижении видового разнообразия при формировании монодоминантных сообществ в реках, имеющих значительный градиент минерализации.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (проект № 07-04-96610) и в рамках программы Президиума РАН «Биологическое разнообразие» (раздел «Динамика биоразнообразия и механизмы обеспечения устойчивости биосистем»).
Список литературы Фитопланктон высокоминерализованных рек Приэльтонья
- Алимов А.Ф. Биологическое разнообразие и структура сообществ организмов//Биология внутренних вод. 2010. №3. С.3-10.
- Алекин О.А. Основы гидрохимии. Л.: Гидрометеоиздат, 1970. 444 с.
- Бенинг А.Л., Медведева Н.Б. О микрофауне водоемов окрестностей Эльтона и Баскунчака//Изв. краевед. ин-та изучения Юж.-Волж. обл. Саратов: Сарполиграфпром, 1926. Т. 1. 39 с.
- Буркова Т.Н. Характеристика фитопланктона высокоминерализованной реки Хара//Известия Пензенского госпедуниверситета. Естественные науки. 2011. № 25. С. 493-496.
- Буркова Т.Н. Таксономический состав альгофлоры планктона высокоминерализованной реки Хара//Самарская Лука: проблемы региональной и глобальной экологии. 2012. Т. 21. № 3. С. 25-35.
- Голубков С.М., Балушкина Е.В., Голубков М.С., Литвинчук Л.Ф., Губелит Ю.И. Взаимоотношения между биоразнообразием и функционированием экосистем на примере соленых озер//Тезисы докладов отчетной научной сессии по итогам работ 2006 г. СПб. ЗИН РАН. 2005. С. 11-12.
- Водно-болотные угодья Приэльтонья. Волгоград: Региональный центр по изучению и сохранению биоразнообразия. Издательство: ООО «Видео-Хайтек». 2005. 27 с.
- Зинченко Т.Д., Головатюк Л.В. Биоразнообразие и структура сообществ макробентоса соленых рек аридной зоны юга России (Приэльтонье)//Аридные экосистемы. 2010. Т. 16. № 3 (43). С. 25-33.
- Зинченко Т.Д., Головатюк Л.В., Выхристюк Л.А., Шитиков В.К. Разнообразие и структура сообществ макрозообентоса высокоминерализованной р. Хара (Приэльтонье)//Поволжский экологический журнал. № 1, 2010. С. 14-30.
- Методика изучения биогеоценозов внутренних водоемов. М.: Наука, 1975. 240 с.
- Михеева Т.М., Лукьянова Е.В. Значимость пикопланктонной фракции в фитопланктонных сообществах пресноводных экосистем//Гидробиол. журн. 2000. Т. 36, N. 6. С. 3-14.
- Номоконова В.И., Зинченко Т.Д., Попченко Т.В. Трофическое состояние соленых рек бассейна озера Эльтон//Изв. Самар. НЦ РАН. 2013. № 3. С. 476-483.
- Отчет о проведении полевых работ по изучению водно-болотных угодий природного парка «Эльтонский», организованных в рамках проекта PIN-MATRA «Институциональное обеспечение водно-болотных угодий в Волгоградской области». 2003. Волгоград. 41 с.
- Петрищев В.П. Солянокупольный ландшафтогенез Прикаспийско-Предуральского региона//Вестник Оренбургского госуниверситета. Изд-во ОГУ. 2007, № 3. С. 143-149.
- Сафонова Т.А., Ермолаев В.И. Водоросли водоемов системы озера Чаны. Новосибирск. 1983.153 с.
- Федоров В.Д. Особенности организации биологических систем и гипотеза «вспышки» вида в сообществе//Вестник МГУ. Сер. биол. 1970. № 2. С. 71-81.
- Яценко-Степанова Т.Н., Немцева Н.В, Шабанова С.В. Альгофлора Оренбуржья//Екатеринбург, 2005. 201 с.
- Яценко-Степанова Т.Н., Игнатенко М.Е., Немцева Н.В., Горохова О.Г. Автотрофные микроорганизмы устьевых участков водотоков системы озера Эльтон//Аридные экосистемы. 2015. в печати
- Klymiuk V.N., Barinova S.S., Lyalyuk N.M. Diversity and Ecology of Algal Communities from the Regional Landscape Park «Slavyansky Resort», Ukraine//Research and Reviews: Journal of Botanical Sciences. 2014. V. 3, № 2. P. 9-26.
- Rojo C., Cobelas M.A., Arauzo M. An elementary, structural analysis of river phytoplankton. Hydrobiologia, 1994. 289 (1), 43-55.
- Søndergaard M. Phototrophic picoplankton in temperate lakes: seasonal abundance and importance along a trophic gradient Internat. Rev. Hydrobiologia, 1991. 76: 505-22.
- Taukulis F.E. Diatom communities in lakes and streams of varying salinity from south-west Western Australia: distribution and predictability. 2007.
- Walker K.F. Studies on a saline lake ecosystem//Australian Journal of Marine and Freshwater Research. 1973. Vol. 24(1). P. 21-72.
- Weisse T. Dynamics of autotrophic picoplankton in marine and fresh water ecosystems//In: J.G. Jones (ed.). Advances in Microbial Ecology. 1993. New York. P. 327-370.
