Физико-химическая и экологическая характеристика амилолитических ферментов колорадского жука

Автор: Цветков Вячеслав Олегович, Шпирная Ирина Андреевна, Валиахметова Карина Ильдаровна, Яруллина Любовь Георгиевна, Ибрагимов Ринат Исмагилович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
В данной статье описано выделение амилаз насекомых, дана их физико-химическая, биохимическая и экологическая характеристика.
Амилолитические ферменты, очистка белков, физико-химическая характеристика белков
Короткий адрес: https://sciup.org/148202029
IDR: 148202029 | УДК: 577.151.01
Physico-chimical and ecological characterization of amylolitic enzymes of Colorado potato beetle
In this article purification of amylases of insects is described, their physic-chemical, biochemical and ecological features is done.
Текст научной статьи Физико-химическая и экологическая характеристика амилолитических ферментов колорадского жука
Из личинок колорадского жука были выделены амилазы, гидролизующие картофельный крахмал. Для очистки ферментов был использован аффинный сорбент на основе полиакриламида с иммобилизованным крахмалом. С помощью двумерного электрофореза показано, что аффинноочищенные амилазы личинок насекомых представлены 4-мя белками с молекулярными массами в диапазоне 3050 кДа. Наибольшую активность исследуемые ферменты проявляли при рН 7 и температуре 30-40 °С. В зависимости от состава пищевого субстрата (картофель, томаты, баклажаны) изменялся уровень активности амилаз личинок. Состав пищи не оказывал влияния на молекулярный состав ферментов.
Колорадский жук Leptinotarsa decemlineata Say является одним из самых опасных вредителей пасленовых культур, в первую очередь картофеля.
Колорадский жук – олигофаг, и фактор влияния кормового растения для вида имеет особое значение. Все активные фазы жизни насекомого проходят непосредственно на растениях.
Одним из основных биохимических инструментов, позволяющих этим насекомым эффективно перерабатывать и усваивать растительную пищу, являются гидролитические ферменты. В соответствии с составом пищевых субстратов, в тканях насекомых присутствуют ферменты для расщепления белков, жиров и углеводов пищи: протеиназы, преимущественно цистеиновые [1], целлюлазы, α-амилазы, эстеразы, представленные множественными формами [2]. Наиболее подробно изученными являются протеазы, экспериментальные сведения о других гидролитических ферментах пока малочисленны. Амилазы являются одними из наиболее активных карбогидраз колорадского жука [3].
В связи с вышесказанным, изучение состава и свойств амилаз этого опасного вредителя является актуальной задачей. Наша работа посвящена выделению и очистке амилаз из личинок колорадского жука, исследованию их состава и свойств.
Для выделения ферментов использовали личинок колорадского жука III стадии развития, собранных с вегетирующих растений картофеля, и баклажана в Уфимском районе РБ. Навеску замороженных личинок гомогенизировали в ступке (5 г) и экстрагировали в трехкратном объеме ацетатного буфера, pH 5,6, при 4 °С в течение 10 мин. Экстракт дважды центрифугировали на центрифуге Eppendorf 5417R при ускорении 8000g в течение 10 мин при 4°С.
Для очистки ферментов использовали аффинный сорбент, синтезированный по оригинальной методике [4]. В качестве лиганда на сорбенте иммобилизовали специфический субстрат амилаз – крахмал.
Выделение ферментов производили на хроматографе BioLogic LP (Biorad, США) при 4°С. На колонку (2х6 см) с иммобилизованным крахмалом, уравновешенную рабочим буфером, наносили 8 мл супернатанта экстракта и промывали рабочим буфером. Элюцию активного белка производили трис-HCl-буфером (рН 8). Скорость тока буфера составляла 2 мл/мин. Ход хроматографии контролировали по оптической плотности элюата при длине волны 280 нм.
Амилолитическую активность белков определяли методом агарозных гелевых пластин [5]. Концентрацию белка определяли по Брэдфорд [6].
Для определения оптимума рН к аликвотам препарата в четырехкратной повторности добавляли соответствующее количество соляной кислоты или гидроксида натрия до достижения требуемого значения рН и выдерживали в течение 10 мин при 4°С. Для определения температурного оптимума раствор выдерживали в течение 10 мин при нужной температуре. Затем определяли ферментативную активность.
Для определения молекулярного состава полученных ферментов проводили SDS-электрофорез в 12%-ном ПААГ (0.75 мм) по Лэммли в камере для вертикального электрофореза (Biorad, США). Электрофорез вели при постоянном напряжении 90 В и силе тока 70 мА.
Для определения изоэлектрической точки полученных ферментов проводили изоэлектрическое фокусирование в 12%-ном ПААГ (1 мм). Изоэлектрическое фокусирование вели при напряжении 1000 В и силе тока 2 мА.
После электрофореза гели фиксировали в 15%-ной ТХУ в течение 10 ч, затем окрашивали раствором Кумасси R-250 в 8%-ной уксусной кислоте и
25%-ном этаноле в течение 20 мин. Окрашивание проводили при комнатной температуре.
Выделение белков. В таблице 1 приведены результаты выделения препарата амилаз из личинок колорадского жука на колонке с полиакриламидным гелем с иммобилизованным картофельным крахмалом. Для получения препарата белка с ферментативной активностью объединяли хромотографические фракции, обладающие высокими значениями амилазной активности; конечный объем раствора составил 12 мл.
Таблица 1 . Аффинная очистка амилаз личинок колорадского жука на колонке с иммобилизованным крахмалом
|
О |
s' Ю 5 CJ о Ы |
tR Ц S 5 я о S m О H V ce |
tR « й Л ® g H § Щ S cd |
я -^ Й M Я g Л я й н Ц g и и Я о « У я |
|
|
Исходный экстракт |
3 |
600 |
50 |
150 |
0,08 |
|
Элюат |
12 |
20 |
42 |
500 |
2,10 |
Амилолитическую активность во фракциях контролировали с помощью метода агарозных гелевых пластин. Как видно, с иммобилизованным субстратом полиакриламидной колонки связывалось около 240 мг белка с ферментативной активностью около 500 Е. Удельная активность ферментов в процессе очистки повысилось в 26 раз, с 0,08 в исходном экстракте до 2,1 Е/мкг белка в элюате.
рН-оптимум и температурная стабильность амилаз. Амилазы колорадского жука проявляют активность в относительно широком диапазоне кислотности среды. Как видно, гидролиз крахмала осуществляется ферментами в интервале от 6 до 8 рН, максимальная активность отмечена при нейтральном значении рН (рис. 1а). По этому показателю амилазы жука существенно не отличаются от описанных в литературе амилолитических ферментов микроороганизмов и различных животных, в т.ч. насекомых и млекопитающих [7]. Исследования альфа-амилазы колорадского жука показали ее способность функционировать в широком диапазоне значений рН (от 6 до 10) с оптимумом около 6,5. Максимальная амилолитическая активность отмечена в переднем отделе кишечника колорадского жука, тогда как в среднем она пренебрежимо мала, а в заднем отсутствует вовсе [8].
Амилолитические ферменты колорадского жука проявляют наибольшую активность в диапазоне от 30 до 40°С (рис. 1б). При воздействии температуры выше 60°С ферментативная активность практиче- ски полностью утрачивается. Содержание и активность амилаз в тканях насекомых существенно зависит от природы пищевого субстрата [9]. Одной из причин такой зависимости может быть влияние пищи на внутреннюю среду насекомого, в частности, на значение рН. В то же время, известно, что оптимум рН для многих ферментов насекомых зависит от их филогенетического положения [8]. По нашим данным, диапазон температуры, в котором функционирует амилазы личинок, также весьма широк (от 25 до 45°С), хотя оптимум составляет 37°С.
Молекулярный состав амилаз. Для более детального исследования молекулярного состава выделенных ферментов, определения молекулярной массы и изоэлектрической точки белков полученные ферменты разделяли методом двумерного электрофореза в 10%-ном ПААГ в денатурирующих условиях.
Результаты показывают, что амилазы колорадского жука представлены четырьмя белками (или группами белков) с молекулярной массой в диапазоне от 30 до 50 кДа и значениями рI от 5 до 6,5 (рис. 2). По-видимому, данные белки являются альфа-амилазами, среди которых известны представители с молекулярной массой 40-60 кДа [10]. Возможно, среди исследованных белков присутствуют также низкомолекулярные формы бета-амилаз или фрагменты их молекул, содержащие центр связывания с субстратом.
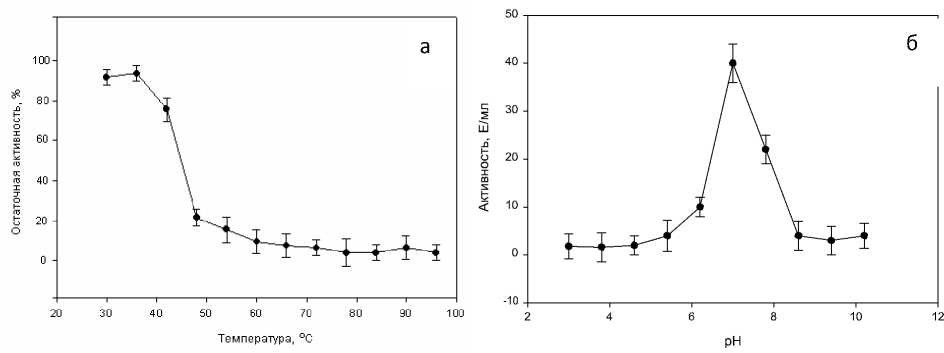
Рис. 1 . Зависимость ферментативной активности амилаз личинок колорадского жука от температуры (а) и величины рН (б). Величину активности определяли при четырехкратной повторности опытной и контрольной проб методом агарозных гелевых пластин. В качестве величины погрешности показан доверительный интервал выборочного среднего
45— 35
25—
3 4 5 6 7 8 9 10
Рис. 2 . Двумерный электрофорез амилаз личинок колорадского жука в 10%-ном SDS-ПААГ. Диапазон рН 3-10. Окраска Кумасси G-250
Влияние кормового субстрата на активность и молекулярный состав амилаз. Наши эксперименты свидетельствуют, что активность амилолитических ферментов насекомых также зависит от состава пищевого субстрата. Как видно из рис. 3, у личинок, выращенных на растениях картофеля сорта Невский, наблюдается наибольший уровень активности амилаз. Личинки, питающиеся листьями картофеля сорта Башкирский и листьями баклажана, имеют более низкие показатели, чем в варианте с сортом Невский. Низкий уровень ферментативной активности в этих случаях может быть связан с частичным ингибированием активности амилаз компонентами пищи [11]. Интересно отметить, что отсутствие пищи течение 48 ч не изменяет активность амилаз личинок (по сравнению с вариантом с сортом Невский). В то же время, активность протеолитических и пектолитических ферментов у личинок значимо снижается при отсутствии корма [12].
На рис. 4 приведены результаты разделения ферментов личинок, питавшихся различными субстратами.
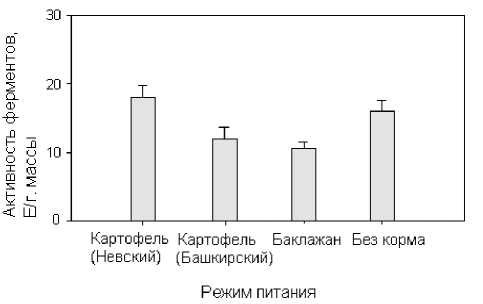
Рис. 3 . Активность гидролитических ферментов в гомогенате личинок колорадского жука в зависимости от пищевого субстрата. «Невский», «Башкирский» – сорта картофеля, «Без корма» – личинки, лишенные пищи в течение 48 ч
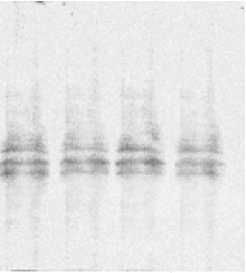
Как показывает разделение очищенных белков методом SDS-электрофореза по Лэммли, во всех исследуемых вариантах спектр белков идентичен. Эти результаты свидетельствуют, что сортовые и видовые особенности растительной пищи могут изменять относительную активность молекулярных компонентов ферментов, но не оказывают влияния на их молекулярный состав. Между тем, некоторыми исследователями было показано, что в зависимости от вида растения, которым питается колорадский жук, способен изменяться количественный и качественный состав протеолитических ферментов [13, 14].
Таким образом, наши исследования показывают, что амилолитические ферменты личинок колорадского жука III стадии развития представлены, по крайней мере, 4-мя белками с молекулярными массами в интервале 30-50 кДа и со значеними рI в слабокислой зоне рН. При питании личинок различными видами и сортами растений активность амилаз изменяется, однако молекулярный состав ферментов остается постоянным.
12 3 4
Рис. 4. SDS-электрофорез амилаз личинок колорадского жука при кормлении различным растительными субстратами. 1. Картофель (Невский, контроль); 2. Картофель (Башкирский); 3. Баклажан; 4. Без корма (48 ч). Окраска Кумасси G-250
Исследования поддержаны грантами: госкон-тракт № 01201353578.
-
1. Thie N., Houseman J. Identification of cathepsin B, D and H in the larval midgut of Colorado potato beetle, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae) // Insect Bi-ochem. 1990. V. 20. N 3. P. 313-318.
-
2. Khorram M., Adab R., Yazdaniyan M., Jafarnia S . Digestive alpha-amylase from Leptinotarsa decemlineta (Say) (Co-leoptera: Chrysomelidae): response to pH, temperature and some mineral compounds // Adv. Environ. Biol. 2010. V. 4. N 1. P. 101-107.
-
3. Рябченко Н.А., Никитин Н.И. Влияние пищевого фактора на микро-эволюцию колорадского жука // Вестн. Днепропетровск. ун-та. 2006. Т. 1. С. 165-171.
-
4. Цветков В.О., Шевченко Н.Д., Ибрагимов Р.И., Гарипова М.И. Выделение протеиназ колорадского жука с использованием аффинного сорбента на основе полиакриламида // Мат. 14-й междунар. Пущинской шк.-конф.
-
5. Шпирная И.А., Умаров И.А., Шевченков Н.Д., Ибрагимов Р.И . Определение активности гидролаз и их ингибиторов с использованием субстратов, иммобилизованных в геле агарозы // Прикл. биохимия и микробиология. 2009. Т. 45. № 4. С. 497-501.
-
6. Bradford M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding // Anal. Biochem. 1976. V. 72. P. 248254.
-
7. Horvathova V, Janecek S, Sturdik E. Amylolytic enzymes: their specificities, origins and properties // Biologia, Bratislava. 2000. V. 55. N 6. P. 605-615.
-
8. Zang F., Zhu Y., Cohen A. Molecular cloning and partial characterization of a trupsin-like in salivary gland of Lygus disponsi (Hemiptera: Mirridae) // Insect Biochem. Mol. Biology. 2002. V. 32. P. 455-464.
-
9. Khorram M., Adab R., Yazdaniyan M., Jafarnia S. Digestive alpha-amylase from Leptinotarsa decemlineta ( Say) (Co-leoptera: Chrysomelidae): response to pH, temperature and some mineral compounds // Adv. Environ. Biol. 2010. V. 4. N 1. P. 101-107.
-
10. Matsuura Y., Kusunoki M., Harada W., Kakudo M. Structure and Possible Catalytic Residues of Taka-Amylase A // J. Bi-ochem. 1984. V. 95. N 3. P. 697-702.
-
11. Умаров И.А. Экологические и физиологобиохимические закономерности взаимоотношений в системе "картофель-колорадский жук": Автореф. дис. … канд. биол. наук. Уфа, 2009. 16 с.
-
12. Цветков В.О. Гидролитические ферменты и их белковые ингибиторы как компоненты взаимодействия картофеля с колорадским жуком: Автореф. дис. … канд. биол. наук. Уфа, 2011. 16 с.
-
13. Overney S., Fawe A., Yelle S., Michaud D. Diet-related plasticity of the digestive proteolytic system in larvae of the Colorado potato beetle ( Leptinotarsa decemlineata Say) // Insect Biochem. Physiol. 1997. V. 36. N 4. P. 241-250.
-
14. Zhu-Salzman K., Zeng R . Molecular mechanisms of insect adaptation to plant defense: Lessons learned from a Bruchid beetle // Insect Sci. 2008. V. 15. N 6. P. 477-481.
молодых ученых «Биология – наука XXI века». Пущино, 2010 г. С. 66-67.
PHYSICO-CHIMICAL AND ECOLOGICAL CHARACTERIZATION OF AMYLOLITIC ENZYMES OF COLORADO POTATO BEETLE
-
L.G. Yarullina2, R.I. Ibragimov1
-
1Bashkir State University, Ufa
-
2Institute of Biochemistry and Genetics, Ufa Sci. Centre of RAS, Ufa
Список литературы Физико-химическая и экологическая характеристика амилолитических ферментов колорадского жука
- Thie N., Houseman J. Identification of cathepsin B, D and H in the larval midgut of Colorado potato beetle, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae)//Insect Biochem. 1990. V. 20. N 3. P. 313-318.
- Khorram M., Adab R., Yazdaniyan M., Jafarnia S. Digestive alpha-amylase from Leptinotarsa decemlineta (Say) (Coleoptera: Chrysomelidae): response to pH, temperature and some mineral compounds//Adv. Environ. Biol. 2010. V. 4. N 1. P. 101-107.
- Рябченко Н.А., Никитин Н.И. Влияние пищевого фактора на микро-эволюцию колорадского жука//Вестн. Днепропетровск. ун-та. 2006. Т. 1. С. 165-171.
- Цветков В.О., Шевченко Н.Д., Ибрагимов Р.И., Гарипова М.И. Выделение протеиназ колорадского жука с использованием аффинного сорбента на основе полиакриламида//Мат. 14-й междунар. Пущинской шк.-конф. молодых ученых «Биология -наука XXI века». Пущино, 2010 г. С. 66-67.
- Шпирная И.А., Умаров И.А., Шевченков Н.Д., Ибрагимов Р.И. Определение активности гидролаз и их ингибиторов с использованием субстратов, иммобилизованных в геле агарозы//Прикл. биохимия и микробиология. 2009. Т. 45. № 4. С. 497-501.
- Bradford M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding//Anal. Biochem. 1976. V. 72. P. 248-254.
- Horvathova V, Janecek S, Sturdik E. Amylolytic enzymes: their specificities, origins and properties//Biologia, Bratislava. 2000. V. 55. N 6. P. 605-615.
- Zang F., Zhu Y., Cohen A. Molecular cloning and partial characterization of a trupsin-like in salivary gland of Lygus disponsi (Hemiptera: Mirridae)//Insect Biochem. Mol. Biology. 2002. V. 32. P. 455-464.
- Khorram M., Adab R., Yazdaniyan M., Jafarnia S. Digestive alpha-amylase from Leptinotarsa decemlineta (Say) (Coleoptera: Chrysomelidae): response to pH, temperature and some mineral compounds//Adv. Environ. Biol. 2010. V. 4. N 1. P. 101-107.
- Matsuura Y., Kusunoki M., Harada W., Kakudo M. Structure and Possible Catalytic Residues of Taka-Amylase A//J. Biochem. 1984. V. 95. N 3. P. 697-702.
- Умаров И.А. Экологические и физиолого-биохимические закономерности взаимоотношений в системе "картофель-колорадский жук": Автореф. дис. … канд. биол. наук. Уфа, 2009. 16 с.
- Цветков В.О. Гидролитические ферменты и их белковые ингибиторы как компоненты взаимодействия картофеля с колорадским жуком: Автореф. дис. … канд. биол. наук. Уфа, 2011. 16 с.
- Overney S., Fawe A., Yelle S., Michaud D. Diet-related plasticity of the digestive proteolytic system in larvae of the Colorado potato beetle (Leptinotarsa decemlineata Say)//Insect Biochem. Physiol. 1997. V. 36. N 4. P. 241-250.
- Zhu-Salzman K., Zeng R. Molecular mechanisms of insect adaptation to plant defense: Lessons learned from a Bruchid beetle//Insect Sci. 2008. V. 15. N 6. P. 477-481.
