Физиологическая специализация возбудителя бурой ржавчины пшеницы в Красноярском крае

Автор: Мешкова Л.В., Россеева Л.П., Сидоров А.В., Сабаева О.Б., Зверовская Т.С., Белан И.А.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 1, 2019 года.
Бесплатный доступ
Цель исследования - мониторинг расового со- става, динамика изменения вирулентности возбу- дителя бурой ржавчины и поиск эффективных ис- точников устойчивости. Представлены много- летние результаты анализа вирулентности при- родной популяции возбудителя бурой ржавчины пшеницы Красноярского края. Определен расовый игенотипический состав 716 монопустульных изо- лятов патогена с использованием дифференци- рующих наборов сортов растения-хозяина. Выяв- лено 11 физиологических рас гриба: 10,20, 57, 68,77, 107, 117, 122, 144, 172 и 184, в зависимости отгода и сорта доминируют 10, 20, 77 и 122 расы. Ежегодно в спорообразцах присутствует 77-я расас частотой встречаемости от 11,0 % в 2003 г. до 96,4 % в 2016 г. При определении патотипического состава патогена на 16 изогенных линиях было выявлено 47 патотипов, в период 2000-2006 гг. доминировали патотипы SGFT - 14,3 % и SGKT - 12,3 %, позднее TJTT - 30,5 % и TJPT - 25,9 %, аTJTT и TGTT отмечены во все годы изучения. Раз- личия между патотипами обусловлены в основном генами вирулентности рр 3а, 3kа, 9, 11, 24 и 26...
Регион, пшеница, устойчи вость, ржавчина, монопустульный изолят, раса, патотип
Короткий адрес: https://sciup.org/140243341
IDR: 140243341 | УДК: 632.4:633.11(571.51)
Physiological specialization of brown rust pathogen on the wheat in Krasnoyrsk region
The research objective was monitoring of racial structure, dynamics of change of a virulence of brown rust activator and the search of effective sources of sta- bility. Long-term results of virulence analysis of natural population of causative agent of brown rust of wheat of Krasnoyarsk Region were presented. Racial and geno- typic composition of 716 monopustulan isolates of the pathogen was determined using differentiating sets of varieties of host plant. 11 physiological races of the fun- gus were identified: 10, 20, 57, 68, 77, 107, 117, 122,144, 172 and 184, depending on a year and variety, 10, 20, 77 and 122 race predominated. Every year in spore samples there were 77 races with the frequency of11.0 % in 2003 up to 96.4 % in 2016. When determining pathogenesis of pathogen on 16 isogenic lines, 47 pathotypes were identified, in the period 2000-2006, the patents SGFT-14.3 % and SGKT - 12.3 % dominated, later TJTT - 30.5 % and TJPT - 25.9 %, two - TJTT and TGTT, were noted during all the years of study...
Текст научной статьи Физиологическая специализация возбудителя бурой ржавчины пшеницы в Красноярском крае
Введение. Бурая ржавчина пшеницы, вызываемая урединиальной стадией гриба Puccinia triticina Erikss et Senn, несмотря на определенные успехи при создании устойчивых сортов, и сегодня остается одним из биотических факторов снижения урожайности и негативного воздействия на качество получаемой продукции [1–3]. Наиболее экономически оправданный и экологически обоснованный способ снижения потерь от этого заболевания – создание и внедрение в производство устойчивых сортов.
Успех при этом во многом зависит от правильности выбора генов устойчивости или их комбинаций, перспективных для использования в селекционных программах при создании резистентных сортов в предполагаемом регионе их выращивания, что вызывает необходимость в определении структуры популяции патогена и ее изменчивости.
Сужение генетического разнообразия в пользу немногих генов резистентности, как указывают С.П. Мартынов и Т.В. Добротворская, может вызвать адекватное изменение в популяции возбудителя и массовое размножение патогена на однородном генетическом материале [4].
Так, в результате широкого распространения в производстве сортов пшеницы с одинаковым генетическим контролем устойчивости Кавказ (Lr 26), Ершовская 32, Куйбышская 1, Юна (Lr 23), Л 503, Юлия, Волгоуральская (Lr 19) в популяциях появились и быстро распространились биотипы патогена с генами вирулентности р 26, р 23 и р 19, что и привело к поражению ранее устойчивых сортов [5–7]. Аналогичная картина наблюдалась и при выращивании сортов (Терция, Соната, Дуэт и др.) в Омской, Челябинской и Новосибирской областях, устойчивость которых детерминирована геном Lr 9 [8–11].
По данным Е.И. Гультяевой, доля сортов пшеницы – носителей гена Lr 9, включенных в ГР РФ и рекомендованных к выращиванию в стране в 2012 г., составила 9 %, в Уральском регионе – 18 %, а в Западносибирском – 17 % [12, 13].
Основным приемом в выявлении изменения в природных популяциях возбудителей заболеваний является постоянный мониторинг их вирулентности. Полученные данные по наличию генов вирулентности патогена, частоте их встречаемости в популяции позволяют осуществлять поиск источников устойчивости среди форм растения-хозяина для дальнейшего включения их в селекционный процесс.
Цель исследования : мониторинг расового состава, динамика изменения вирулентности возбудителя бурой ржавчины и поиск эффективных источников устойчивости.
Материалы и методы исследования . Сбор инфекционного материала возбудителя бурой ржавчины осуществлялся в период массового проявления заболевания на посевах яровой мягкой пшеницы Красноярского края с восприимчивых сортов селекционных учреждений Западной и Восточной Сибири: Алтайская 70, Ветлужанка, Кантегирская 89, Новосибирская 15, Новосибирская 29, Омская 9, Омская 20, Омская 32, Омская 33, Омская 28, Омская 36, Свирель и Тулунская 12, включенных в ГР РФ и допущенных для выращивания в 10-м и 11-м регионах. Также был изучен споровый материал с селекционных линий пшеницы лаборатории селекции яровой пшеницы КрасНИИСХ, которые, предположительно, были получены в результате скрещивания с сортами, несущими ген устойчивости к листовой ржавчине Lr 9.
Гербарные образцы (листья пшеницы с уреде-ниопустулами), начиная с 2000 г., ежегодно изучали в лаборатории иммунитета растений ФГБНУ «Омский АНЦ» (СибНИИСХ) по расовому составу на стандартном наборе сортов-дифференциаторов: Malаkоff (Lr 1), Corina (Lr 2в), Brevit (Lr 2с), Webster (Lr 2а), Loros (Lr 2с), Mediterranean (Lr 2а+3а), Hussar (Lr 11) и Democrat (Lr 3а). Наличие генов вирулентности в популяции определяли на серии моногенных линий сорта Thatcher (Тс): Lr 1, 2а, 2b, 2с, 3а, 3bg, 3ka, 9, 10, 11, 14a, 14b, 15, 16, 17, 18, 19, 20, 21, 23, 24, 25, 26, 27+31, 28, 29, 30, 31, 32, 33, 36, 38, В, – и линии, полученной от И.Г. Одинцовой с геном Lr Sp. С 2007 г. набор был расширен линиями Lr 43, 44, 45, 46, 47, 48, 49, 50, а с 2012г. и Lr 41. Фенотипический состав определяли по реакции 16 изогенных линий по буквенному ключу, предложенному D.L. Long и J.A. Kolmer, основанному на определении вирулентности 16 линий, сгруппированных по четыре линии в четыре блока: I – Lr 1, Lr 2а, Lr 2с, Lr 3а; II – Lr 9, Lr 16, Lr 24, Lr 26; III – Lr 3ka, Lr 11, Lr 17, Lr 30 и IV – Lr В, Lr 10, Lr 14а, Lr 18 [14].
Возобновляли спорообразцы и размножали мо-нопустульные изоляты патогена на универсально- восприимчивом сорте яровой мягкой пшеницы Саратовская 29.
Мониторинг расового и генотипического состава бурой ржавчины пшеницы осуществляли по методике отсеченных листьев в светокультуре с использованием раствора бензимидазола [15]. Тип реакции растения на внедрение патогена определяли по международной шкале в модификации Джонстона и Бровдера [16], где 0, 1, 2 – устойчивость (R), 3, 4 – восприимчивость (S), Х – гетерогенность. По частоте встречаемости фенотипов патогена в популяциях грибов судили об их сходстве или различии, согласно формуле Л.А. Животовского: r = ∑min (p,q), где r – коэффициент сходства; p и q – минимальные частоты фенотипов в одном из двух сравниваемых образцов [17].
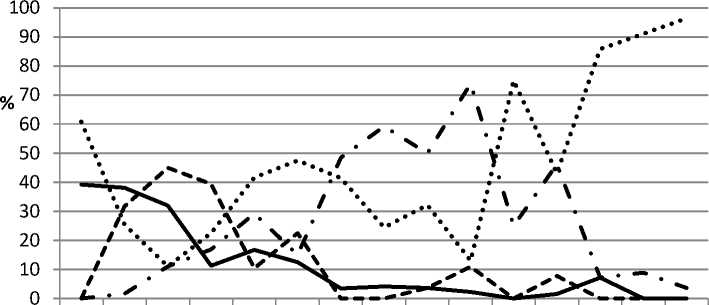
Результаты исследования и их обсуждение . Анализ расового состава 716 монопустульных культур бурой ржавчины за период с 2000 по 2017 г. выявил наличие 11 физиологических рас гриба: 10, 20, 57, 68, 77, 107, 117, 122, 144, 172 и 184, – доминируют в зависимости от года и сорта – 10, 20, 77 и 122. Ежегодно в популяциях присутствует 77 раса в среднем с частотой встречаемости 53,3 % и варьирует от 11,0 % в 2003 г. до 96,4 % в 2016 г. Средняя частота встречаемости 122-й расы составляет 33,3 % и колеблется от 0 до 73,9 % в 2009 г.; 20-я раса зафиксирована в 11 годах (68,7 %), доминирует в 2002 г., с частотой встречаемости 38,1 %; 10-я раса встречается 8 раз (50 %) и преобладает в 2003 г. (45,0 %) (рис. 1).
Физиологические расы 57, 107, 117, 172 и 184 выявлены дважды за период наблюдения, раса 68 – три раза, а 144 – один, с максимальной частотой встречаемости до 10,2 %. Следует отметить увеличение частоты встречаемости 77-й расы и снижение 10, 20 и 122-й рас, которые практически элиминировали в спо-рообразцах последних лет сборов (табл. 1).
Патотипический состав возбудителя бурой ржавчины определяли на 16 изогенных линиях растения-хозяина. Было установлено, что популяция Красноярского края представлена 47 патотипами, наибольшее разнообразие отмечено в 2005 г. – выявлено 27 патотипов; наименьшее в 2012 г. – 3. Основные: TJTT, TGTT, TKTT, THTT, TJTS, TSTT, TTTT, PJTT, THTS, KHTT, KJTT и PGTT, – в период с 2000 по 2006 г. доминировали патотипы SGFT – 14,3 % и SGKT– 12,3 %, позднее (2007–2016 гг.) TJTT – 30,5 % и TJPT – 25,9 %, третье месте в оба периода занимал патотип TGTT с частотой встречаемости 11,9 и 13,8 % соответственно.
20002002200320052006200720082009201020112012 2013201420152016
Год
Раса 10
Раса 20
Раса 77
Раса 122
Рис. 1. Частота встречаемости основных рас возбудителя бурой ржавчины пшеницы Красноярского края
Таблица 1
Расовый состав возбудителя бурой ржавчины пшеницы Красноярского края, 2000–2016 гг., %
|
Год |
Раса |
||||||||||
|
10 |
20 |
57 |
68 |
77 |
107 |
117 |
122 |
144 |
172 |
184 |
|
|
2000 |
39,2 |
60,8 |
|||||||||
|
2002 |
31,7 |
38,1 |
1,6 |
25,4 |
1,6 |
1,6 |
|||||
|
2003 |
45,0 |
32,0 |
1,0 |
11,0 |
11,0 |
||||||
|
2005 |
39,4 |
11,3 |
9,9 |
22,5 |
16,9 |
||||||
|
2006 |
10,4 |
16,7 |
41,6 |
2,1 |
29,2 |
||||||
|
2007 |
22,5 |
12,5 |
2,5 |
47,5 |
15,0 |
||||||
|
2008 |
3,4 |
41,4 |
48,4 |
3,4 |
3,4 |
||||||
|
2009 |
4,1 |
24,5 |
2,0 |
59,2 |
10,2 |
||||||
|
2010 |
3,6 |
3,6 |
3,6 |
32,1 |
8,8 |
50,0 |
7,1 |
||||
|
2011 |
10,9 |
2,2 |
13,0 |
73,9 |
|||||||
|
2012 |
75,0 |
25,0 |
|||||||||
|
2013 |
7,7 |
1,5 |
43,1 |
46,2 |
1,5 |
||||||
|
2014 |
7,1 |
85,8 |
7,1 |
||||||||
|
2015 |
91,1 |
8,9 |
|||||||||
|
2016 |
96,4 |
3,6 |
|||||||||
Анализ монопустульных изолятов бурой ржавчины на изогенных линиях сорта Thatcher (Tc) показал, что большинство линий используемого набора проявили 100 %-ю восприимчивость, более 80 % поражения имели линии Lr: 1, 2в, 2с, 10, 18, 23 и 29, – что говорит о низкой дифференцирующей способности этих линий. Существенные различия выявлены только по генам Lr 26, 45 и 47. Не поразились или проявили высокую устойчивость к монопустульным изолятам линии с генами Lr 9, Lr 19 и Lr 41, независимо от генотипа растения-хозяина и года сбора инокулюма. Эти гены переданы в мягкую пшеницу от других видов: Lr 9 – Aegilops umbellulata, Lr 19 – Thynopyrum elongatum, Lr 41 – Aegilops tauschii, Lr 26 – Secale cerealе, Lr 47 – Aegilops speltoides [18]. Проявляют иммунитет линии мягкой пшеницы c генами устойчивости Lr Sp, которые были привнесены в геном мягкой пшеницы от амфидиплоида Triticum dicoccum – Aegilops speltoides [19].
Таким образом, для изучения изменений в структуре красноярской популяции патогена считаем более приемлемым использование Lr линий с генами устойчивости 3а, 3bg, 3ka, 9, 11, 19, 24, 26, 36, 45, 47 и Sp (табл. 2).
Таблица 2
Частота встречаемости генов вирулентности в популяции возбудителя бурой ржавчины пшеницы, 2000–2016 гг.
|
Год |
Гены вирулентности, % |
||||||||||
|
3а |
3bg |
3ka |
9 |
11 |
19 |
24 |
26 |
36 |
45 |
47 |
|
|
2000 |
- |
86,9 |
- |
0 |
50,0 |
0 |
39,1 |
4,3 |
- |
- |
- |
|
2002 |
38,1 |
30,2 |
38,1 |
0 |
65,1 |
0 |
28,6 |
3,2 |
3,9 |
- |
- |
|
2003 |
25,0 |
20,0 |
2,01 |
0 |
45,0 |
2,0 |
24,0 |
17,0 |
34,0 |
- |
- |
|
2005 |
43,7 |
43,7 |
48,0 |
0 |
30,9 |
0 |
73,2 |
15,5 |
90,1 |
7,1 |
- |
|
2006 |
59,1 |
50,7 |
83,3 |
0 |
58,3 |
2,1 |
20,8 |
8,3 |
83,3 |
0 |
- |
|
2007 |
95,0 |
65,0 |
60,0 |
0 |
65,0 |
0 |
57,5 |
5,0 |
72,5 |
0 |
0 |
|
2008 |
96,5 |
89,6 |
93,1 |
0 |
69,0 |
3,4 |
93,1 |
0 |
96,5 |
0 |
0 |
|
2009 |
100 |
100 |
83,7 |
4,1 |
26,5 |
0 |
30,6 |
0 |
42,9 |
0 |
2,0 |
|
2010 |
89,3 |
82,3 |
82,3 |
0 |
39,3 |
0 |
89,3 |
3,6 |
89,3 |
0 |
0 |
|
2011 |
86,9 |
86,9 |
91,3 |
0 |
15,2 |
2,2 |
100 |
2,0 |
- |
0 |
0 |
|
2012 |
100 |
100 |
100 |
0 |
75,0 |
0 |
95,8 |
0 |
87,5 |
4,2 |
2,1 |
|
2013 |
94,8 |
94,8 |
94,8 |
0 |
49,3 |
2,6 |
75,3 |
20,8 |
67,5 |
2,1 |
6,7 |
|
2014 |
100 |
96,4 |
92,9 |
12,5* |
85,7 |
7,1 |
64,3 |
3,6 |
64,3 |
8,6 |
5,7 |
|
2015 |
100 |
100 |
100 |
0 |
91,1 |
0 |
67,9 |
14,3 |
17,9 |
6,8 |
7,1 |
|
2016 |
100 |
100 |
100 |
0 |
96,3 |
0 |
57,1 |
25,0 |
57,1 |
4,3 |
3,6 |
|
Среднее |
69,9 |
67,2 |
68,4 |
0,6 |
52,8 |
1,3 |
56,6 |
11,0 |
61,1 |
2,8 |
2,7 |
*С сорта Свирель – 12,5 %; с линий К-459-2; 512-7; 540-10 – 40 %.
В таблице 2 приведены данные (отсутствуют 2001 и 2004 гг.), которые показывают, что, начиная с 2006 г., наблюдается увеличение вирулентности популяций к линиям с генами устойчивости Lr 3а, 3bg и 3ka и вариабельность по генам Lr 11, 24, 26 и 36. Возможно, это связано с изменениями в сортовом разнообразии и, соответственно, площадями под выращиваемыми сортами. До 2006 г. в посевах превалировал сорт Тулунская 12 и сорта селекции
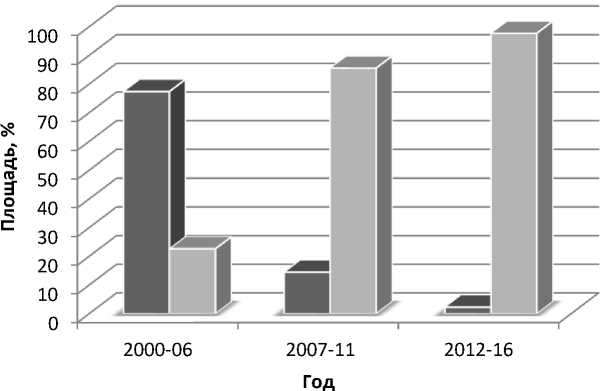
Восточной Сибири (Скала, Тулун 15, Ветлужанка) и незначительные площади занимали сорта Омской и Новосибирской селекции (Омская 9, Омская 20, Омская 32, Омская 33, Кантегирская 89). Начиная с 2007 г., отмечено преобладание сортов западносибирской селекции, к 2016 г. их доля в структуре посевных площадей составила 98,67 %, в основном это сорта селекции СибНИИРСа – Новосибирская 15, Новосибирская 29, Новосибирская 31 и др. (рис. 2).
■ ВС
■ ЗС
Рис. 2. Площадь посева сортов яровой мягкой пшеницы в Красноярском крае, % (ВС – сорта Восточной Сибири; ЗС – сорта Западной Сибири)
Анализ родословных выращиваемых сортов показал, что в сортах, созданных в ВосточноСибирском регионе, преобладали в основном формы местной селекции. Большинство же сортов западносибирской селекции получено с использованием сортов краснодарской и поволжской селекции – Безостая 1, Кавказ, Саратовская 29, Саратовская 36, Безенчукская 98 и др. Проведенная сортосмена с увеличением площади под сортами западносибирской селекции способствовала, на наш взгляд, и изменению популяции патогена. Это подтверждается и расчетами коэффициентов сходства. Расчет сходства по патотипам бурой ржавчины с сортов различного происхождения показал, что между патотипами спорообразцов с сорта Тулунская 12 и патотипами с сортов Омская 28, Омская 20 и Кантегир-ская 89 коэффициент составил 10–15 %, что касается патотипов с сортов Омская 20, Омская 28 и Кан-тегирская 89, то их сходство составило 50–55 %.
Учитывая то, что бурая ржавчина может распространяться с воздушными потоками, провели сравнение патотипов патогена Красноярского края и соседних регионов (Омск, Новосибирск), предположительно оказывающих влияние на состав бурой ржавчины на примере структуры популяций 2014 г.
В результате проведенных исследований в популяции 2014 г. было выявлено 14 патотипов, в т. ч. в Омске – 12, Красноярске – 6 и в Новосибирске – 3. В изученных спорообразцах преобладали патотипы TJTT, TКTT и TGTT, различающиеся в основном по частоте встречаемости генов вирулентности рр 9, 24 и 26. Коэффициент сходства между популяциями Красноярск / Омск равнялся 48,8 %, Красноярск / Новосибирск – 40,0 % и Омск / Новосибирск – 74,5 %. Отличие популяции из Красноярска от популяций из Омска и Новосибирска обусловлено генами р 11 и р 26 (табл. 3).
Таблица 3
|
Фенотип |
Формула вирулентности |
Пункт сбора, % |
||
|
Красноярск |
Омск |
Новосибирск |
||
|
TJTT |
9, 26/S |
17,9 |
48,3 |
42,9 |
|
TКTT |
9/S |
10,7 |
20,2 |
45,7 |
|
TGTT |
9, 24, 26/S |
28,6 |
19,1 |
11,4 |
|
THTT |
9, 24/S |
0 |
8,8 |
0 |
|
TSTT |
26/S |
25,0 |
1,1 |
0 |
|
TQTT |
24, 26/S |
7,1 |
0 |
0 |
|
TJTS |
9, 26, 18/S |
0 |
0,5 |
0 |
|
TJPT |
9, 26, 11 /S |
10,7 |
0 |
0 |
|
Другие |
- |
0 |
2,0 |
0 |
Частота встречаемости патотипов бурой ржавчины, % (2014 г.)
Расчет коэффициентов сходства (r) по патотипам показал среднюю связь популяции Восточной Сибири (Красноярск) с Западной Сибирью (Омск, Новосибирск) и высокую связь западносибирских популяций, что говорит о возможности проникновения спорового материала с воздушными массами из Западной Сибири.
Выводы. В результате проведенных исследований (2000–2016 гг.) генофонда популяций возбудителя бурой ржавчины пшеницы Восточной Сибири не выявлено генов, вирулентных к линиям с генами устойчивости Lr 41 и Lr Sp. Несмотря на появление в популяциях патотипов патогена, поражающих сорта и линии с генами Lr 9 и Lr 19, они по-прежнему остаются высокоэффективными (R > 90 %), частичную устойчивость (поражение ≤ 15 %) проявляют гены Lr 26, Lr 45 и Lr 47.
Таким образом, мониторинг вирулентности бурой ржавчины пшеницы Красноярского края показал, что структура популяции патогена обусловлена в основном генотипом выращиваемых сортов и площадями под ними, но не исключается возможность и заноса инфекционного материала, что подтверждается коэффициентом сходства. Полученные данные необходимо учитывать при планировании и создании устойчивых сортов.
Список литературы Физиологическая специализация возбудителя бурой ржавчины пшеницы в Красноярском крае
- Крупнов В.А. Стратегия генетической защиты пшеницы от листовой ржавчины в Поволжье//Вестн. РАСХН. -1997. -№ 6. -С. 12-15
- Санин С.С. Влияние вредных организмов на качество зерна//Защита и карантин растений. -2004. -№ 11. -С. 14-18.
- Лубнин А.Н. Селекция яровой мягкой пшеницы в Сибири. -Новосибирск, 2006. -31 с
- Мартынов С.П., Добротворская Т.В. Генеалогический подход к анализу устойчивости пшеницы к болезням//Фитосанитарное оздоровление экосистемы: мат-лы II Всерос. съезда по защите растений (Санкт-Петербург, 5-10 декабря 2005 г.). -СПб., 2005. -Т. 1. -С. 511-513.
- Михайлова Л.А., Тырышкин Л.Г., Гусева Н.Н. Исследование популяций возбудителя бурой ржавчины//Защита растений. -1988. -№ 3. -С. 18-19.
- Шаповалова О.Ю. Генетическая структура популяции возбудителя бурой ржавчины пшеницы на Северном Кавказе//Агро XXI. -2003. -№ 1-6. -С. 11.
- Сюков В.В., Вьюшков А.А., Шевченко С.Н. и др. Генетические основы создания сортов яровой мягкой пшеницы, устойчивых к грибным болезням в Среднем Поволжье//Генетика, селекция и семеноводство сельскохозяйственных культур: сб. науч. тр. к 100-летию Самарского НИИСХ. -Самара, 2003. -С. 128-147.
- Мешкова Л.В., Россеева Л.П. Тенденция увеличения вирулентности возбудителя бурой ржавчины пшеницы к эффективным генам устойчивости в Омской области//Современные средства, методы и технологии защиты растений: мат-лы междунар. науч.-практ. конф. -Новосибирск, 2008.-С. 149-153.
- Мешкова Л.В., Россеева Л.П., Шрейдер Е.Р. и др. Вирулентность патотипов возбудителя бурой ржавчины пшеницы к Th Lr9 в регионах Сибири и Урала//Современные проблемы иммунитета растений к вредным организмам: мат-лы II Всерос. конф. (Санкт-Петербург, 29 сентября -2 октября 2008 г.). -СПб., 2008. -С. 70-73.
- Мешкова Л.В., Россеева Л.П., Сидоров А.В. и др. Вирулентность возбудителя бурой ржавчины пшеницы в регионах Сибири и Урала//Иммуногенетическая защита сельскохозяйственных культур от болезней: теория и практика: мат-лы междунар. науч.-практ. конф., посвящ. 125-летию со дня рождения Н.И. Вавилова (Большие Вяземы Московской области, 17-21 июля 2012 г.). -Большие Вязьмы, 2012. -С. 237-241.
- Сочалова Л.П., Христов Ю.А. Влияние генотипа сорта на структуру популяции возбудителя бурой ржавчины пшеницы Puccinia recondite//Сибирский Вестник с-х науки. -2009. -№ 10. -С. 61-67.
- Гультяева Е.И. Генетическое разнообразие российских сортов мягкой пшеницы по устойчивости к возбудителю бурой ржавчины//Докл. Россельхозакадемии. -2012. -№ 2. -С. 29-32.
- Гультяева Е.И., Аристова М.К., Шайданюк Е.Л. и др. Генетическая дифференциация Puccinia triticina Erikss на территории России//Генетика. -2017. -Т. 53, № 9. -С. 1053-1060
- Long D.L., Kolmer J.A. A North American System of Nomenclature for Puccinia triticina//Phytopathology. -1989. -Val. 79. -P. 525-529.
- Михайлова Л.А., Квитко К.В. Лабораторные методы культивирования возбудителя бурой ржавчины пшеницы//Микология и фитопатология. -1970.-Т. 4, № 3.-С. 269-270
- Ghonston C.O., Browder B.E. Seventh revision of physiologic races of Puccinia recondita f. sp.tritici//Plant Dis. Reptr. -1966. -Val. 50. -P. 756-760
- Животовский Л.А. Показатели сходства популяций по полиморфным признакам//Журн. общей биологии. -1979. -Т. 11, № 4. -С. 587-602.
- Каталог мировой коллекции ВИР. Вып. 453. Сорта зерновых культур с известными генами устойчивости к грибным болезням. -Л., 1988. -80 с.
- Богуславский Р.Л., Одинцова И.Г., Пеуша Х.О. Амфидиплоиды и редкие формы пшеницы как источники устойчивости к бурой ржавчине//Проблемы использования генофонда в селекции растений на иммунитет к болезням и вредителям: сб. науч. тр. по прикл. бот., генет. и селекции. -Л., 1987. -Т. 110. -С. 18-23.
