Физиологические исследования древесных растений

Автор: Молчанов А.Г.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: К 75-летию института лесоведения ран
Статья в выпуске: 4, 2019 года.
Бесплатный доступ
Приведены результаты изучения водного и светового режимов, а также углеродного баланса лесов в разных лесорастительных зонах, засухоустойчивости и теневыносливости разных пород деревьев. Построена математическая модель определения углеродного баланса разных пород в различных климатических условиях. Разработана методика оценки фотосинтетической продуктивности древостоев, основанная на зависимости КПД использования солнечной радиации на фотосинтез охвоенных или облиственных побегов и поглощения пологом древостоя солнечной радиации.
Интенсивность фотосинтеза, дыхание древесных растений, радиационный режим древостоев, солнечная радиация, фотосинтез
Короткий адрес: https://sciup.org/143169946
IDR: 143169946 | УДК: 581.1; | DOI: 10.24419/LHI.2304-3083.2019.4.02
Physiological studies of woody plants
The Laboratory of Plant Physiology was founded at the Forest Institute in 1944 under the supervision of a member of correspondent USSR Academy of Sciences L.A. Ivanova. During this time, the laboratory studied the water, light conditions and carbon balance of the forest in different forest zones. The dependence of the number and spectral composition of PARs under the forest canopy on the throughput of the canopy of the forest stand was investigated. The physiological mechanism of drought tolerance of trees in the steppe zone has been studied. It has been shown that there are various ways of adapting tree species to drought. The joint existence of species with different types of adaptation of trees to arid conditions (for example, oak, fluffy ash and small-leaved elm) reduces the life of plantings. Artificial shading studied the effect of light deficiency on growth, leaf anatomy and carbon metabolism in seedlings of different species and revealed a physiological mechanism shade tolerance...
Текст научной статьи Физиологические исследования древесных растений
Для ссылок:
Молчанов, А. Г. Физиологические исследования древесных растений [Электронный ресурс] / А. Г. Молчанов // Лесохоз. информ.: электрон. сетевой журн. – 2019. – № 4. – С. 23–31. URL:
Л аборатория физиологии и экологии древесных пород в Институте леса АН СССР была создана в 1944 г., ее руководителем стал член-корреспондент АН СССР Л. А. Иванова. Сотрудники лаборатории занимались вопросами определения интенсивности транспирации растений. В 1949 г. был разработан метод быстрого взвешивания [1]. Одновременно с транспирацией в разных климатических зонах изучали фотосинтез и образование органического вещества древостоями, в результате чего была установлена параллельная динамика водного и углеродного режимов [2, 3].
В дальнейшем, для определения предела необратимого обезвоживания листьев у разных древесных пород, был разработан специальный метод [4], с помощью которого установлено, что в засушливом климате могут выживать породы с разным строением корневых систем, например дуб и ясень. У дуба глубокие корни, в его почках до глубокой осени медленно протекает эмбриональный рост побегов, и к концу вегетационного периода полностью формируется побег следующего года. Ясень пушистый обладает густой сетью поверхностных корней и быстро тратит влагу верхнего горизонта почвы. В середине лета в листьях ясеня возникает необратимый водный дефицит, и они засыхают. Однако благодаря быстрому формированию в почках побегов будущего года рост на следующий год проходит успешно. Эти исследования проводили на Деркульской опытной станции.
Деркульскую опытную станцию закрыли в 1957 г., Институт леса перевели в Красноярск, а в Московской обл. (с. Успенское) была организована Лаборатория лесоведения АН СССР под руководством академика В. Н. Сукачева. Экспериментальные работы по физиологии растений стали проводить в Серебряноборском опытном лесничестве. Здесь на пробных площадях изучали радиационный режим под пологом леса и его влияние на рост и процессы газообмена СО2 в нижних ярусах леса. Газообмен определяли методом Иванова-Коссович [5].
Радиационный режим леса изучали совместно с сотрудниками Института светотехники, которые разработали и изготовили оригинальные приборы для измерений спектрального состава радиации в лесу, а также помогали в составлении методики измерений [6, 7]. В результате изучения роста и интенсивности фотосинтеза подроста на участках с разной сомкнутостью крон установлено, что при слабой освещенности уровень плато фотосинтеза снижается [8]. Однако морфологическая и функциональная перестройка растений при затенении имеет адаптивный характер. Так, у растений, произрастающих в условиях недостаточности света в толще листа, количество хлоропластов уменьшается; они становятся более крупными, возрастает так называемое сопротивление диффузии СО2, зависящее от снижения активности фотосинтетического фермента рибулезодифосфат-карбоксилазы. Это и является причиной сниженного уровня плато фотосинтеза у теневыносливых растений по сравнению со светолюбивыми, а также затененных растений по сравнению с растущими при хорошем освещении [9, 10].
В 1962 г., когда лабораторию возглавила Ю. Л. Цельникер, начали проводить исследования по углеродному газообмену и балансу органического вещества в разных органах деревьев. В процессе работ изучена сезонная динамика образования и расходования углерода у листвы, ветвей и баланс углерода для целого дерева, проведены исследования физиологических механизмов, влияющих на сезонную динамику роста побегов. В течение года определяли оводненность точек роста побегов (на верхушках молодых побегов, а затем и в почках, после их формирования), способность поглощать воду, интенсивность их дыхания и содержание нуклеиновых кислот (ДНК и РНК).
Результаты исследования показали, что точки роста побегов осенью и зимой (в состоянии глубокого покоя) не способны поглощать воду. Во время глубокого покоя нуклеиновые кислоты в почках не синтезируются. При распускании листьев и быстром видимом росте идет ускоренное образование нуклеиновых кислот, интенсивность дыхания остается высокой, но способность к фотосинтезу развивается лишь постепенно, по мере образования хлорофилла. Рост листьев и хвои происходит за счет импорта органического вещества из запасов ветвей. По мере нарастания площади листьев и хвои их функция меняется – они становятся экспортерами. Так, например, лист березы становится экспортером органического вещества при достижении 20% его конечной площади, а лист дуба – около 90% [11].
Время распускания почек у древесных пород различается и зависит от сроков наступления среднесуточной положительной температуры. Этот показатель имеет различные значения у разных древесных пород (ранораспускающиеся с низкой стартовой температурой и позднораспу-скающиеся – с высокой), но для каждой породы эта температура постоянна [12].
Кроме того, в процессе исследований изучалось поглощение углерода ассимилирующими органами, которое складывается из затрат на ночное дыхание, на дыхание скелетных частей и на прирост. Каждая ветка тратит только тот углерод, который сама ассимилировала; продолжительность её жизни определяется углеродным балансом, а избыток углерода, не истраченный веткой, экспортируется и служит источником углеродного питания остальных частей дерева. [13, 14]. Проведены расчеты углеродного баланса ветвей разного возраста. Установлено, что ветви погибают, если баланс углерода становится ниже, чем необходимые затраты на рост ветвей следующего года. На этом основании был установлен предельный возраст жизни ветвей. Определена зависимость интенсивности дыхания ствола от его диаметра, высоты, температуры ствола и воздуха. Установлено, что температурный коэффициент дыхания Q10 для всех пород равен 2,2.
Разработанная климатическая модель роста и длительности жизни леса разного породного состава позволила установить, в каких климатических условиях может произрастать лес определенного породного состава, и сделать прогноз трансформации растительности при изменении климата [16].
С 1969 г. в с. Городище Ярославской обл. был создан Опорный пункт Северной ЛОС (под руководством Ю. Л. Цельникер и В. В. Осипова).
Здесь проведены исследования по сравнению продуктивности двух основных лесообразующих светолюбивых пород – сосны и березы, произрастающих в оптимальных лесорастительных условиях, двумя методами:
-
1) лесоводственным (по методике А. А. Молчанова и В. В. Смирнова), который предполагал взятие модельных деревьев по ступеням толщины (взвешиванием всех частей дерева в абсолютно сухом состоянии), с последующим пересчетом биомассы на 1 га в зависимости от распределения деревьев по ступеням толщины и высоты;
-
2) эколого-физиологическим – оригинальный метод, основанный на связи КПД использования поглощенной солнечной радиации на фотосинтез и поглощенной пологом древостоя солнечной радиации.
Экофизиологическая методика была представлена на международной конференции в Таллинне, затем в Монреале и опубликована в журнале Tree Physiology [17]. Она основана на оценке поглощенной солнечной радиации пологом древостоя, которую легко можно определить инструментальным способом и оценить поглощенную охвоенным побегом радиацию в фотоинтегрирующей сфере. По этой методике определяли поглощение ФАР побегами сосны, находящимися в разных слоях полога соснового древостоя (световые-теневые). Эти данные использовали для расчета фотосинтетической продуктивности соснового древостоя [18]. Результаты оценки продуктивности древостоев сосны и березы по двум методам получились довольно близкими. Так же мало различается лесоводственная продуктивность 40-летних насаждений исследуемых пород (сосны и березы), произрастающих в одинаковых оптимальных лесорастительных условиях. В процессе работы была изучена зависимость интенсивности фотосинтеза сосны и березы от окружающих условий [19]. Одновременно с изучением биопродуктивности надземной части сосны и березы лесоводственным методом была определена биопродуктивность подземной части исследуемых древостоев, изучен суточный ход и сезонная интенсивность дыхания корневых мочек и скелетной части корней сосны и березы [20].
Проведено сравнение интенсивности фотосинтеза сосны, произрастающей в крайних по продуктивности типах леса, в заболоченном низкопродуктивном пушицево-сфагновом сосняке и высокопродуктивном сосняке чернично-кисличном. Установлено, что в заболоченном типе леса при уровне грунтовых вод ниже 40 см интенсивность фотосинтеза практически такая же, как в чернично-кисличном сосняке, а при уровне грунтовых вод 5–10 см в заболоченном сосняке интенсивность фотосинтеза была в 2 раза ниже, чем в высокопродуктивном [21]. В результате изучения эмиссии СО2 с поверхности почвы в этих типах леса получена нетто-продуктивность по газообмену СО2 сосновых древостоев [22, 23], а также установлена зависимость интенсивности газообмена сфагнума в болотном сосняке пушицево-сфагновом от уровня почвенно-грунтовых вод.
В 1997 г., в связи с реорганизацией института, исследования по физиологии древесных растений в Серебряноборском лесничестве были прекращены. Однако с 2005 г. экофизиологиче-ские исследования были возобновлены благодаря А. И. Гурцеву.
В рамках этих исследований разработана методика оценки эмиссии СО2 из почвы методом беспрерывной регистрации по открытой схеме [24] и методика оценки газообмена одновременно на 5 объектах. Для этого создано устройство, состоящее из газоанализатора «КЕДР» (Москва) или LiCor 820, логера Mini32, 10 пневмопереключателей, расходомера и 5 спаренных вибрационных побудителей расхода воздуха SONIC 388. При помощи этой установки можно проводить мониторинг эколого-физиологических показателей в экосистемах (интенсивности фотосинтеза, дыхания стволов, эмиссии СО2 с поверхности почвы с корнями и почвы без корней) с одновременной регистрацией параметров окружающей среды. По разработанной методике исследовано влияние сплошной рубки леса на эмиссию СО2 с поверхности почвы [25].
В Серебряноборском опытном лесничестве на основе этой методики определена интенсивность фотосинтеза ели и сосны, эмиссии СО2 с поверхности стволов, проведено сравнение эмиссии СО2 с поверхности стволов и с поверхности почвы, а также сравнение эмиссии с поверхности почвы на участках с корнями и без корней.
Продолжены исследования по оценке предрассветного водного потенциала хвои (ПВПХ) в различных условиях недостатка влаги. В связи с климатическими изменениями, необходимо оценить выживаемость саженцев древесных растений на открытом месте, на вырубках и под пологом леса. В настоящее время ведутся исследования по оценке выживаемости подроста сеянцев под пологом леса при недостатке ПВПХ и освещенности. Освещенность под пологом леса будет определяться методом фотографирования крон цифровым фотоаппаратом с широкоугольным объективом, с дальнейшей обработкой по программе LAI – hemisfer122 (Швеция).
Установлена высокая чувствительность древесных растений к изменению радиационного и температурного режимов в разных условиях водообеспеченности. Однако до сих пор неизвестно, при каком значении ПВПХ у деревьев ели происходит уменьшение продуктивности, а при каком – снижение интенсивности фотосинтеза.
Подрост ели нормально функционирует при ПВПХ от -0,4 до -0,9 МПа, при этом фотосинтез выходит на плато насыщения солнечной радиации при 100–150 Вт/м2, достигая 6 мкмоль СО2/м2с, а при довольно значительном недостатке влаги (ПВПХ = -1,4 МПа) фотосинтез ели снижается почти в 2 раза. Интенсивность фотосинтеза подроста при световом насыщении составила почти 4 мкмоль СО2/м2с, а при ПВПХ = -2,5 МПа фотосинтез ели фактически сменился на дыхание (рисунок).
В таблице представлены уравнения зависимости интенсивности фотосинтеза от солнечной радиации. Коэффициент детерминации при ПВПХ = -2,5 МПа показывает, что интенсивность фотосинтеза в таких условиях водообеспеченно-сти не зависит от солнечной радиации.
Таким образом, у саженцев ели под пологом леса при ПВПХ ниже -1,4 МПа интенсивность фотосинтеза уменьшается в 2 раза, а при -2,5 МПа и ниже газообмен становится отрицательным.
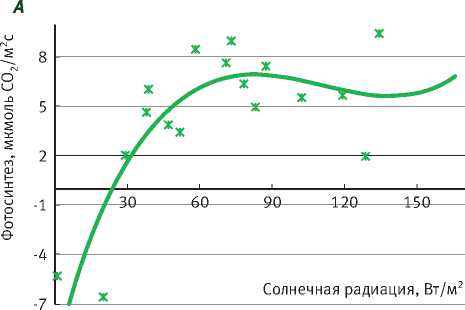
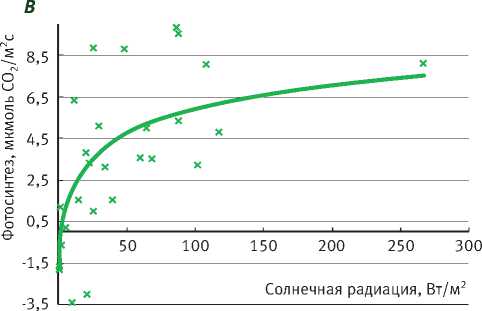
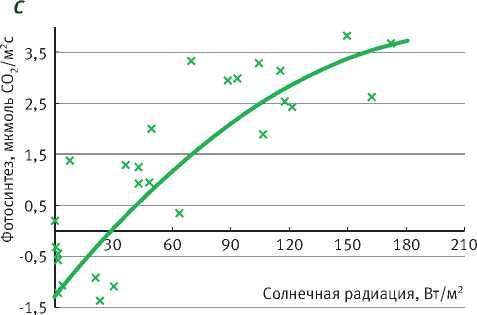
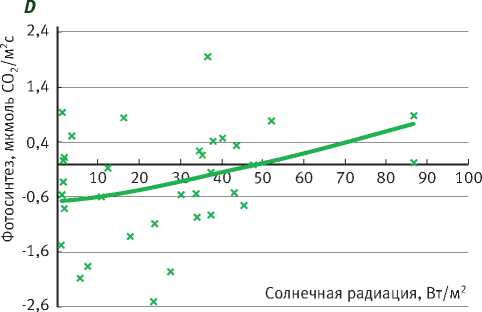
Зависимость фотосинтеза ели от солнечной радиации в разных условиях предрассветного водного потенциала (А = – 0,4 МПа; B = – 0,9 МПа; С = – 1,4 МПа; D= – 2,5 МПа)
Уравнения зависимости фотосинтеза ели от солнечной радиации
|
Значение ПВПХ, Мпа |
Уравнение регрессии |
Коэффициент детерминации |
|
- 0,4 |
y= 2E-05x3 – 0,0051x2 + 0,5165x – 9,8133 |
R2 = 0,75 |
|
- 0,9 |
y = 1,1,631509ln(x) – 1,6114 |
R2 = 0,45 |
|
- 1,4 |
y = -0,0001x2 + 0,0473x – 1,2787 |
R2 = 0,64 |
|
- 2,5 |
y = 7E-05x2 + 0,0106x – 0,6845 |
R2 = 0,09 |
Условные обозначения: y – фотосинтез, мкмоль CO2, x – солнечная радиация, Вт/м2.
Следовательно, при снижении ПВПХ до -1,4 МПа подрост ели, который выжил под пологом леса, в результате ослабления может снова заселиться короедом.
Однако о выживаемости саженцев необходимо судить не только по снижению интенсивности фотосинтеза и водного потенциала, но и по длительности нахождения саженца в экстремальных условиях.
Дальнейшие исследования позволят ответить на следующие вопросы:
-
1. При каком значении предрассветного водного потенциала на открытом месте начинает снижаться интенсивность фотосинтеза у основных лесообразующих пород?
-
2. При каком значении недостатка влаги, предрассветного водного потенциала интенсивность фотосинтеза у основных лесообразующих пород снизится до нуля, смогут ли после такого стресса саженцы восстановиться?
-
3. Какие породы древесных растений наиболее устойчивы к недостатку влаги?
Список литературы Физиологические исследования древесных растений
- Иванов, Л. А. О методе быстрого взвешивания для определения транспирации в естественных условиях / Л. А. Иванов, А. А. Силина, Ю. Л. Цельникер // Ботан. журн. - 1950. - С. 171-185.
- Фотосинтез и транспирация древесных пород в различных климатических зонах / Л. А. Иванов, И. В. Гулидова, Ю. Л. Цельникер, Е. В. Юрина // Водный режим растений в связи с обменом веществ и продуктивность. - М.: изд-во АН СССР, 1963. - С. 121-127.
- Судницын, И. И. О соотношении давления почвенной влаги и сосущей силы листьев древесных пород / И. И. Судницын, Ю. Л. Цельникер // Докл. АН СССР. - 1960. - Т. 151 - № 6. - С. 1457-1459.
- Цельникер, Ю. Л. Скорость потери воды изолированными листьями древесных пород и устойчивость их к обезвоживанию / Ю. Л. Цельникер // Тр. Ин-та леса АН СССР. - 1955. - Т. 27. - С. 6-28.
- Иванов Л.А. Полевой метод определения фотосинтеза в ассимиляционной колбе / Л. А. Иванов, Н. Л. Коссович // Ботан. журн. - 1946. - Т. 31. - № 5. - С. 3-12.
- Пропускание света пологом леса в зависимости от падающей радиации и сомкнутости крон деревьев / Е. А. Акулова, В. С. Хазанов, Ю. Л. Цельникер, Д. М. Шишов // Физиология растений. - 1964. - Т. 11. - Вып. 5. - C. 818-823.
- Цельникер, Ю. Л. Радиационный режим под пологом леса / Ю. Л. Цельникер - М.: Наука. - 1969. - 100 с.
- Малкина, И. С. Фотосинтез и дыхание подроста / И. С. Малкина, Ю. Л. Цельникер, А. М. Якшина. - М.: Наука, 1970. - 183 с.
- Цельникер, Ю. Л. Физиологические основы теневыносливости древесных растений / Ю. Л. Цельникер. - М.: Наука, 1978. - 212 с.
- Tselniker, Y. L. Effects of irradiance on photosynthesis, ribulese-1, 5-biphosphate carboxelase activity and photosynthetic unit size in Populus tremula L. / Y. L. Tselniker, A. G. Chetverikov, T. F. Andreeva // Photosynthetica. - 1983. - V. 17 - № 4. - P. 550-556.
- Цельникер, Ю. Л. Баланс органического вещества в онтогенезе листа у лиственных деревьев / Ю. Л. Цельникер, И. С. Малкина // Физиология растений. - 1986. - Т. 33. - № 5. - С. 40-51.
- Цельникер, Ю. Л. Влияние температур на сроки распускания и скорость роста листа у лиственных деревьев / Ю. Л. Цельникер. // Проблемы экологического мониторинга и моделирования экосистем. - СПб., 1996. - Т. 16. - С. 164-176.
- Цельникер, Ю. Л. Баланс углерода ветвей ели / Ю. Л. Цельникер, И.С. Малкина // Лесоведение. - 1995. - № 4. - С. 16-25.
- Цельникер, Ю. Л. Структура кроны лиственницы / Ю. Л. Цельникер // Лесоведение. - 1997. - № 3. - С. 40-51.
- Рост и газообмен СО2 у лесных деревьев / Ю. Л. Цельникер, И. С. Малкина, А. Г. Ковалев [и др.]. - М.: Наука, 1993. - 256 с.
- Анализ влияния среды на фотосинтез хвойных Предбайкалья / Ю. Л. Цельникер, М. Д. Корзухин, Г. Л. Суворова [и др.] // Проблемы экологического мониторинга и моделирования экосистем. - СПб., 2007. - Т. ХI. - С. 265-293.
- Molchanov, A. G. Photosynthetic utilization efficiency of absorbed photosynthetically active radiation by Scots pine and birch forest stands in the southern Taiga / A. G. Molchanov // Tree Physiology. - 2000. - V.20. - Р. 1137-1148.
- Молчанов, А. Г. Измерение и расчет поглощения ФАР побегами сосны / А. Г. Молчанов, В. С. Хазанов // Лесоведение. - 1975. - № 2. - С. 75-79.
- Молчанов, А.Г. Экофизиологическое изучение продуктивности древостоев / А. Г. Молчанов. - М.: Наука, 1983. - 136 с.
- Мамаев, В. В. Дыхание древесных корней в сосняке и березняке чернично-кисличных / В. В. Мамаев // Лесоведение. - 1984. - № 6. - С. 53-60.
- Молчанов, А. Г. Сравнение интенсивности фотосинтеза сосны в разных эдафических условиях / А. Г. Молчанов // Лесоведение. - 1993. - № 6. - С. 76-80.
- Молчанов, А. Г. Баланс углекислоты в сосновом насаждении южной тайги / А. Г. Молчанов // Лесоведение. - 1990. - № 1. - С.47-53.
- Molchanov, A. G. Estimation of photosynthetic productivity of a forest stands using the efficiency in the utilization of an absorbed radiation by a stand for photosynthesis / A. G. Molchanov // Monitoring of Energy-Mass Exchange between Atmosphere and Forests Ecosystems. Gotttingen. - 2002. - P. 31-42.
- Татаринов, Ф. А. Оценка и минимизация ошибок при измерении дыхания почв по открытой схеме / Ф. А. Татаринов, А. Г. Молчанов, А. В. Ольчев // Изв. Самарского НЦ РАН. - 2009. - Т. 11. - № 1(7). - С. 1592-1595.
- Молчанов, А. Г. Влияние сплошной вырубки леса на эмиссию СО2 с поверхности почвы / А. Г. Молчанов, Ю. А. Курбатова, А. В. Ольчев // Известия РАН. Сер. биологическая. - 2017. - № 2. - С. 190-196.
