Физиологическое состояние Fucus vesiculosus L. Баренцева моря при длительном нахождении в воздушной среде

Автор: Рыжик Инна Валериевна, Макаров Михаил Владимирович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Гидробиология
Статья в выпуске: 2 т.21, 2018 года.
Бесплатный доступ
Увеличение антропогенной нагрузки на экосистемы обусловило необходимость проведения мероприятий, направленных на очищение окружающей среды и восстановление нарушенных биоценозов. Для очистки морской среды от тяжелых металлов, радионуклидов, нефтепродуктов предложено использовать плантации-биофильтры, биологической составляющей которых являются водоросли-макрофиты. В качестве наиболее подходящих объектов определены фукусовые водоросли, в частности Fucus vesiculosus, поскольку они обладают высокой сорбционной активностью по отношению к тяжелым металлам, радионуклидам, способны включать нефтяные углеводороды в процесс метаболизма. При исследовании физиологических особенностей растений с помощью стандартных методов (определение фотосинтетической активности, содержания фотосинтетических пигментов, относительного содержания воды) выявлена способность F. vesiculosus выдерживать длительное нахождение в воздушной среде. В ходе эксперимента водоросли помещали на сетку, скручивали в рулон, который обматывали брезентом (для предотвращения высыхания растений), и устанавливали в разных экспериментальных условиях при постоянной (8 ºС) и переменной (7-19 ºС) температурах. Водоросли сохраняли фотосинтетическую способность в течение 30 дней; через 20 дней наблюдалось снижение интенсивности фотосинтеза по сравнению с контрольными растениями; после эксперимента F. vesiculosus восстанавливали уровень фотосинтетических процессов в течение одних суток. Общее содержание хлорофиллов и соотношение Chl А / Chl C в течение эксперимента не изменялись, что также свидетельствует об устойчивости фотосинтетического аппарата растений. Показано, что F. vesiculosus при нахождении в условиях, обеспечивающих сохранение влаги в талломах не менее 55 %, в течение 30 дней способны сохранять физиологическую активность. Эта способность расширяет возможности использования F. vesiculosus в качестве объекта санитарной аквакультуры; возможно предварительное строительство модулей плантаций-биофильтров и их длительная транспортировка.
Санитарная плантация-биофильтр, биоремедиация, загрязнение нефтепродуктами, баренцево море
Короткий адрес: https://sciup.org/142215137
IDR: 142215137 | УДК: 57.(022 | DOI: 10.21443/1560-9278-2018-21-2-253-260
The physiological state of the Fucus vesiculosus L. of the Barents Sea in a prolonged air condition
The increase in anthropogenic pressure on ecosystems necessitated the implementation of measures aimed at purifying the environment and restoring disturbed biocoenoses. To remediate the marine environment from heavy metals, radionuclides, oil products, it has been proposed to use plantations-biofilters, the biological components of which are seaweeds. Fucoids, in particular Fucus vesiculosus, have been identified as the most suitable objects as they have a high sorption activity to heavy metals, radionuclides, are able to include petroleum hydrocarbons in own metabolism. By the standard methods of plant physiological research (determination of photosynthetic activity, content of photosynthetic pigments, relative water content) the ability of F. vesiculosus to withstand prolonged exposure to air has been determined. In the experiment, the algae were placed on the netting, twisted into a roll wrapped with a tarp (to prevent drying plants) and set in different experimental conditions (in constant (8 ºC) and variable (7-19 ºC) temperatures). Algae retained the photosynthetic capacity for 30 days, but after 20 days there was a decrease in this process compared to control. After the experiment, algae recovered the level of photosynthetic capacity for one day. The total content of chlorophylls and the ratio of Chl A / Chl C during the experiment did not change, which also indicates the sustainability of the photosynthetic apparatus of F. vesiculosus. It has been shown that F. vesiculosus is able to maintain physiological activity within 30 days if it is under conditions that ensure moisture retention in the thalli is not less than 55 %. This ability expands the potential of using F. vesiculosus as an object of sanitary aquaculture: it is possible to pre-build modules of plantation-biofilter and their long transportation.
Текст научной статьи Физиологическое состояние Fucus vesiculosus L. Баренцева моря при длительном нахождении в воздушной среде
В настоящее время увеличивающаяся антропогенная нагрузка на экосистемы обусловила необходимость проведения мероприятий, направленных на очищение окружающей среды и восстановление нарушенных биоценозов. Актуальной проблемой, предваряющей проведение подобных мероприятий, является подбор средств и методов, безопасных для окружающей среды. В ходе проведенных исследований установлен наиболее подходящий способ – биоремедиация, т. е. очистка среды с помощью биологических объектов, в основном микроорганизмов [1–3]. Также возможно применение сорбентов, изготовленных из природных материалов [4–6]. Для очистки морской среды от тяжелых металлов, радионуклидов, нефтепродуктов предложено использовать водоросли-макрофиты в качестве основных компонентов плантации-биофильтра [7–10]. Наиболее подходящими объектами были определены бурые водоросли, поскольку они обладают высокой сорбционной активностью по отношению к тяжелым металлам, радионуклидам, способны включать нефтяные углеводороды в процесс метаболизма [11–13]. Самым устойчивым из фукусовых водорослей является F. vesiculosus [14].
При разработке плантаций-биофильтров одной из задач является создание определенного пула отдельных мобильных секций с возможностью их транспортировки к местам постановки. Для решения данной задачи необходимо определение оптимальных и возможных условий, а также длительности периода, в течение которого водоросли могут находиться вне водной среды без потери жизнеспособности и функциональной активности.
Целью данной работы было определение максимальной продолжительности нахождения талломов F. vesiculosus в безводных условиях посредством оценки физиологического состояния растений (функционального состояния модулей плантации-биофильтра).
Материалы и методы
Экспериментальные работы проводили в июле – августе 2015 и 2017 гг.; использовали талломы Fucus vesiculosus длиной 20–25 см, имеющие 6–8 дихотомических ветвлений; при отборе водорослей не учитывали наличие рецептакулов. Растения закрепляли на сетчатом полотне, которое применяли в качестве основы для отдельной секции плантации [15], скручивали и заворачивали в брезентовую ткань для снижения потерь влаги, формируя рулон. Рулоны находились в термостатированном помещении при температуре 8 ºС (регулируемые условия) и на улице под навесом (для защиты от прямых солнечных лучей), где наблюдались перепады температур от 7 до 19 ºС (нерегулируемые условия). Рулоны периодически
(раз в два дня) поливали морской водой. Поскольку водоросли были завернуты в плотную ткань, они также оказались в условиях отсутствия освещения, необходимого для функционирования фотосинтетического аппарата.
Длительность эксперимента составила 30 суток, в течение которых каждые 10 дней определяли содержание воды в талломах и оценивали физиологическое состояние водорослей по интенсивности фотосинтеза (ИФ) растений скляночным методом в модификации Винклера. Для этого несколько растений изымали из рулона и помещали в сосуды со свежей морской водой. Измерения проводили в течение часа после изъятия водорослей из рулонов в лабораторных условиях при постоянном освещении люминесцентными лампами интенсивностью 100 Вт/м2 и температуре 10 ºС. Для измерения ИФ использовали не менее трех растений из каждого варианта. После проведения измерений растения в экспериментальные рулоны не возвращали.
После 30 суток нахождения водорослей в условиях эксперимента их поместили в естественные условия для оценки возможности и скорости восстановления физиологических показателей. Через 1, 2 и 24 часа определяли содержание воды в талломе и интенсивность фотосинтеза. Для оценки состояния фотосинтетического аппарата в начале и в конце эксперимента, а также через сутки после восстановления растений определяли концентрацию и соотношение хлорофиллов (Хл А, Хл С (С1 + С2)).
Содержание воды в талломах рассчитывали по соотношению сырой и сухой массы высечек таллома (^ = 20). Массу высечек определяли на весах ВЛТЭ-210 с точностью до 0,001 г: сырую – после удаления с поверхности высечек капельно-жидкой влаги фильтровальной бумагой; сухую – после высушивания в сушильном шкафу (Т = 105 °С) в течение 24 часов.
Показатели растений, находившихся в условиях эксперимента, сравнивали с показателями растений из природных зарослей (контроль).
Эксперимент проводили в двукратной повторности; статистическую обработку данных осуществляли с использованием программного пакета Excel. На рис. 1–4 представлены средние арифметические значения и доверительные интервалы.
Результаты и обсуждение
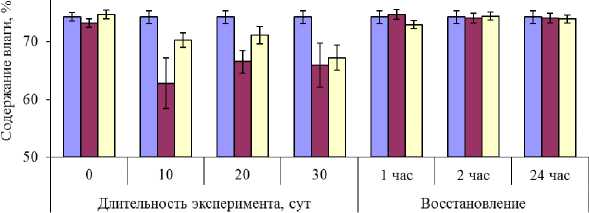
Содержание влаги в талломах растений при естественных условиях (контрольных) составляло 74–75 %. У растений, находившихся в условиях эксперимента, содержание влаги со временем снижалось. Минимальное содержание влаги составляло 63 % и наблюдалось у растений, помещенных в термостатируемые (регулируемые) условия (рис. 1), возможно, вследствие того, что постоянство температуры в помещении обеспечивалось работой кондиционера, который создавал постоянное движение воздуха и осушал его. В результате высыхание рулонов с водорослями в помещении происходило быстрее, чем на улице.
80 -i
□ контроль ■регулируемые условия □ нерегулируемые условия
Рис. 1. Содержание влаги в талломах F. vesiculosus в течение эксперимента
Fig. 1. The moisture content in the F. vesiculosus thallus during the experiment
Содержание влаги в тканях растений, находившихся в условиях эксперимента, уменьшалось намного меньше по сравнению с растениями из природных зарослей, у которых во время отлива содержание влаги может снижаться до 30 %, а близкородственный вид F. spiralis может терять до 90 % имеющейся влаги [16], но при наступлении прилива растения быстро восстанавливают физиологическую активность.
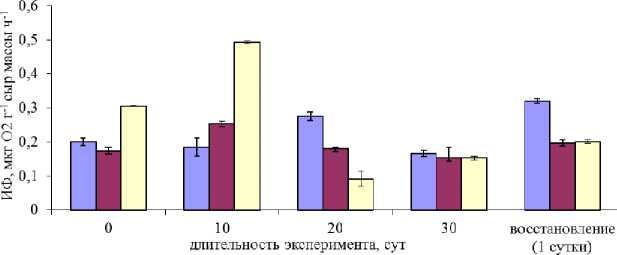
Водоросли при нахождении в воздушной среде сохраняли фотосинтетическую активность в течение всего периода исследований (30 суток), их ИФ была сравнима с показателями растений из природных зарослей (рис. 2).
В течение первых 10 суток у растений, находившихся в воздушной среде, после переноса их в морскую воду отмечалась более высокая ИФ как по сравнению с начальным уровнем, так и по сравнению с контрольными растениями. Возможно, это связано с потерей влаги в период нахождения растений в воздушной среде и активным восстановлением фотосинтетических процессов при переносе в водную среду и достаточном освещении. На 20-е сутки ИФ экспериментальных растений снижалась, что может быть обусловлено индивидуальными особенностями талломов, отобранных из рулонов для определения показателей, поскольку через 30 суток их ИФ была вновь сравнима с показателями контрольных растений.
□ контроль □ регулируемые условия □ нерегулируемые условия
Рис. 2. Интенсивность фотосинтеза F. vesiculosus в течение эксперимента
Fig. 2. The intensity of photosynthesis of F. vesiculosus during the experiment
Общее содержание хлорофиллов (рис. 3, А) и соотношение Хл А / Хл С (рис. 3, Б) в течение эксперимента не менялось, что также свидетельствует о сохранности фотосинтетического аппарата F. vesiculosus при длительном нахождении растений в воздушной среде и при отсутствии освещения.
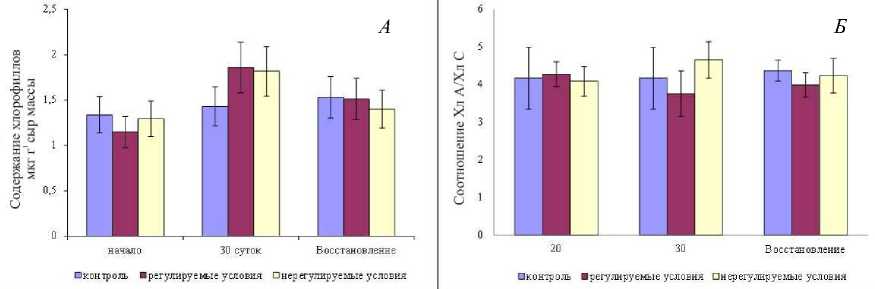
Рис. 3. Содержание хлорофиллов (А) и соотношение Хл А / Хл С (Б) в ткани F. vesiculosus в течение эксперимента и в период восстановления
Fig. 3. Chlorophyll content (A) and Chl A / Chl C ratio (Б) in the F. vesiculosus tissue during the experiment and the period of recovery
После переноса растений из условий эксперимента в нормальные условия (свежая морская вода и естественное освещение) происходило быстрое восстановление фотосинтетической активности: уже через час ИФ экспериментальных растений была всего в 1,5 раза ниже, чем контрольных, через два часа показатели достигли уровня контрольных растений. Через сутки ИФ была в два раза выше по сравнению с начальным уровнем (рис. 4). Неизменность содержания и соотношения фотосинтетических пигментов и быстрое восстановление функциональных показателей свидетельствуют о сохранности фотосинтетического аппарата в активном состоянии в течение всего периода эксперимента.
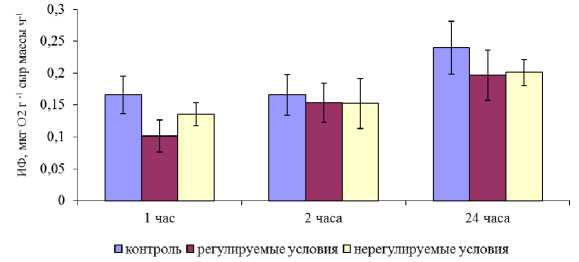
Рис. 4. Интенсивность фотосинтеза экспериментальных растений F. vesiculosus в период восстановления
Fig. 4. The intensity of photosynthesis of experimental plants of F. vesiculosus during the recovery period
В ряде работ по изучению влияния осушения на водоросли было показано, что скорость восстановления физиологических показателей зависит от количества потерянной влаги: если достигнута критическая потеря воды, то восстановления ИФ не происходит. Для каждого вида водоросли максимальная степень осушения, при которой еще возможно восстановление физиологической активности, индивидуальна [17; 18]. F. vesiculosus может выдерживать более длительное осушение по сравнению с другими фукоидами и быстро восстанавливать потерянную влагу: растения способны терять до 91 % воды [19] и до 32 часов находиться в воздушной среде без потери жизнеспособности [20; 21]. За сохранение жизнеспособности водорослей отвечают дегидрин-подобные белки (члены группы белков тканей наземных растений), устойчивые к высыханию и способствующие стабилизации мембран и ферментов при значительной потере влаги [22; 23]. Способность литоральных водорослей находиться продолжительное время вне водной среды может также определяться возможностью переключения механизмов карбоксилирования и использования атмосферной двуокиси углерода в качестве источника неорганического углерода, что было обнаружено еще в середине XX в. известным физиологом растений R. Bidwell [24].
Заключение
По результатам проведенных исследований [14; 25–28], F. vesiculosus является уникальным видом водорослей, способным существовать в широком диапазоне условий окружающей среды и имеющим высокую степень толерантности к воздействию загрязнителей. В ходе эксперимента показано, что этот вид водорослей может выдерживать отсутствие освещения и находиться вне воды при положительных температурах окружающей среды в течение 30 суток.
Практический результат данной работы состоит в установлении возможности хранения или длительной транспортировки указанного вида фукоидов или готовых модулей плантации-биофильтра, изготовленных на его основе, без организации специальных условий. Главным требованием сохранности растительного материала является его содержание в условиях постоянной влажности, доступа воздуха и при отсутствии прямых солнечных лучей.
Работа выполнена в рамках государственного задания ММБИ КНЦ РАН по теме "Механизмы адаптации, регуляции роста, размножения и рационального использования водорослей-макрофитов арктических морей", № 0228-2018-0004.
Список литературы Физиологическое состояние Fucus vesiculosus L. Баренцева моря при длительном нахождении в воздушной среде
- Baker K. H., Herson D. S. Bioremediation. New York: McGraw-Hill, 1994. 375 p.
- Atlas R. M. Bioremediation of petroleum pollutants//International Biodeterioration & Biodegradation. 1995. V. 35, N 1-3. P. 317-327.
- Alexander M. Biodegradation and bioremediation. 2nd Edition. Academic Press, 1999. 453 p.
- Choi H. M., Cloud R. M. Natural sorbents in oil spill cleanup//Environmental Science & Technology. 1992. V. 26, N 4. P. 772-776 DOI: 10.1021/es00028a016
- Thomson B. M. Removal of metals and radionuclides using apatite and other natural sorbents//Journal of Environmental Engineering. 2003. V. 129, N 6. P. 492-499. (ASCE)0733-9372(2003)129:6(492) DOI: https://doi.org/10.1061/
- Hlihor R. M., Gavrilescu M. Removal of some environmentally relevant heavy metals using low-cost natural sorbents//Environmental Engineering and Management Journal. 2009. V. 8, N. 2. P. 353-372.
- Способ очистки морских прибрежных вод от пленочных и диспергированных в поверхностном слое воды нефтепродуктов: пат. 2375315 Рос. Федерация/Воскобойников Г. М., Коробков В. А., Макаров М. В. № 2007106573/13; заявл. 21.02.07; опубл. 10.12.2009, Бюл. № 34.
- Воскобойников Г. М., Макаров М. В., Рыжик И. В. Теоретические основы и перспективы использования плантации-биофильтра для очистки морской акватории от нефтяного загрязнения//Комплексные исследования больших морских экосистем России = Integrated investigations of the Russian large marine ecosystems. Апатиты: КНЦ РАН, 2011. С. 202-234.
- Воскобойников Г. М., Ильинский В. В., Лопушанская Е. М., Макаров М. В., Пуговкин Д. В., Рыжик И. В. . Санитарная водорослевая плантация для очистки прибрежных акваторий от нефтепродуктов: от теории к практике//Вопросы современной альгологии. Специальный выпуск. Доклады Сабининских чтений (2012-2016). М.: Перо, 2017. С. 160-186.
- Воскобойников Г. М., Зубова Е. Ю., Макаров М. В., Пуговкин Д. В., Рыжик И. В. Санитарная водорослевая плантация (СВП): варианты//Биоразнообразие и устойчивое развитие: материалы III междунар. науч.-практ. конф., Симферополь, 15-19 сент. 2014 г. Симферополь: Крымский научный центр, 2014. С. 66.
- Золотухина Е. Ю., Тропин И. В., Кононенко Р. В. Распределение тяжелых металлов в талломах бурой водоросли Laminaria sacharina (Ag.) Kjellm//Вестник Московского университета. Сер. 16: Биология. 1992. № 1. С. 72-76.
- Степаньян О. В. Морфо-функциональные перестройки у водорослей-макрофитов Баренцева моря под воздействием нефти и нефтепродуктов: дис.. канд. биол. наук: 25.00.28. Мурманск, 2003. 146 с.
- Усягина И. С., Воскобойников Г. М., Макаров М. В., Ильин Г. В., Салахов Д. О. Биосорбция радионуклидов водорослями-макрофитами из морской воды (на примере Fucus vesiculosus)//Материалы научных мероприятий, приуроченных к 15-летию Южного научного центра РАН: Междунар. науч. форума "Достижения академической науки на Юге России", Междунар. молодежной науч. конф. "Океанология в XXI в.: современные факты, модели, методы и средства" памяти члена-корреспондента РАН Д. Г. Матишова, Всерос. науч. конф. "Аквакультура: мировой опыт и российские разработки", г. Ростов-на-Дону, 13-16 декабря 2017 г. Ростов н/Д: ЮНЦ РАН, 2017. С. 290-293.
- Макаров М. В., Рыжик И. В., Воскобойников Г. М. Выбор объектов для санитарной аквакультуры, или Fucus vesiculosus vs Терминатор: шансы на выживание//Материалы научных мероприятий, приуроченных к 15-летию Южного научного центра РАН: Междунар. науч. форума "Достижения академической науки на Юге России"; Междунар. молодежной науч. конф. "Океанология в XXI в.: современные факты, модели, методы и средства" памяти члена-корреспондента РАН Д. Г. Матишова; Всерос. науч. конференции "Аквакультура: мировой опыт и российские разработки" г. Ростов-на-Дону, 13-16 декабря 2017 г. Ростов н/Д: ЮНЦ РАН, 2017. С. 406-408.
- Устройство для биологической очистки морских вод от техногенных загрязнений (варианты): пат. на полезную модель 136037 Рос. Федерация/Воскобойников Г. М., Макаров М. В. № 2013137328/10; заявл. 08.08.2013; опубл. 27.12.2013, Бюл. № 36.
- Schagerl M., Möstl M. Drought stress, rain and recovery of the intertidal seaweed Fucus spiralis//Marine Biology. 2011. V. 158, Iss. 11. P. 2471-2479. DOI: https://doi.org/10.1007/s00227-011-1748-x.
- Dring M. J., Brown F. A. Photosynthesis of intertidal brown algae during and after periods of emersion: a renewed search for physiological causes of zonation//Marine Ecology: Progress Series. 1982. V. 8. Р. 301-308.
- Beer S., Kautsky L. The recovery of net photosynthesis during rehydration of three Fucus species from the Swedish west coast following exposure to air//Botanica Marina. 1992. V. 35, Iss. 6. Р. 487-491. DOI: https://doi.org/10.1515/botm.1992.35.6.487. Published Online: 2009-10-23.
- Kanwisher J. Freezing and drying in intertidal algae//Biological Bulletin. 1957. V. 113, N 2. P. 275-285 DOI: https://doi.org/10.2307/1539085
- Kawamitsu Y., Driscoll T., Boyer J. S. Photosynthesis during desiccation in an intertidal alga and a land plant//Plant and Cell Physiology. 2000. V. 41, Iss. 3. P. 344-353 DOI: https://doi.org/10.1093/pcp/41.3.34
- Андреев В. П., Маслов Ю. И., Сороколетова Е. Ф. Функциональные особенности фотосинтетического аппарата трех видов Fucus в условиях Белого моря. Влияние обезвоживания//Физиология растений. 2012. Т. 59, № 2. С. 244-250.
- Xu D., Duan X., Wang B., Hong B., Ho T. H. D. Expression of a late embryogenesis abundant protein gene, HVA1, from Barley confers tolerance to water deficit and salt stress in transgenic rice//Plant Physiology. 1996. V. 110, N 1. P. 249-257 DOI: https://doi.org/10.1104/pp.110.1.249
- Close T. J. Dehydrins: a commonalty in the response of plants to dehydration and low temperature//Physiologia Plantarum. 1997. V. 100, N 2. P. 291-296. DOI: https://doi.org/10.1111/j.1399-3054.1997.tb04785.x.
- Bidwell R. G. S. Photosynthesis and metabolism of marine algae: II. A survey of rates and products of photosynthesis in C14O2//Canadian Journal of Botany. 1958. V. 36, N 3. P. 337-349. 10.1139/b58-029 DOI: https://doi.org/
- Воскобойников Г. М., Макаров М. В., Рыжик И. В. Изменения в составе фотосинтетических пигментов и структуре клеток у бурых водорослей Fucus vesiculosus L. и F. serratus L. Баренцева моря при длительном нахождении в темноте//Биология моря. 2006. Т. 32, № 1. С. 26-33.
- Макаров М. В., Рыжик И. В., Воскобойников Г. М., Матишов Г. Г. Влияние интенсивности движения воды на морфологические и физиологические показатели Fucus vesiculosus L. Баренцева моря//Доклады академии наук. 2007. Т. 415, № 4. С. 569-570.
- Макаров М. В., Рыжик И. В., Воскобойников Г. М., Матишов Г. Г. Влияние глубины произрастания на морфофизиологические показатели Fucus vesiculosus L. Баренцева моря//Доклады академии наук. 2010. Т. 430, № 3. С. 427-429.
- Makarov M. V. Influence of ultraviolet radiation on the growth of the dominant macroalgae of the Barents Sea//Chemosphere: Global Change Science. Climate Change Effect on Northern Terrestrial and Freshwater Ecosystems. 1999. V. 1, N 4. P. 461-467.
