Физиолого-биохимическая характеристика растений Aconitum septentrionale в сообществах Южного Тимана

Автор: Головко Т.К., Табаленкова Г.Н., Малышев Р.В., Далькэ И.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 1-4 т.13, 2011 года.
Бесплатный доступ
Дана эколого-физиологическая и биохимическая характеристика растений аконита высокого (Aconitum septentrionale), произрастающих в разных фитоценозах на Южном Тимане в бассейне р. Сойва. Выявлены различия между растениями в удельной поверхностной плотности листьев, содержании пигментов, накоплении сахаров, фотосинтетической активности и продуктивности, обусловленные преимущественно световыми и эдафическими условиями местообитания.
Продуктивность, фотосинтез, пигменты, сахара
Короткий адрес: https://sciup.org/148199744
IDR: 148199744 | УДК: 581.1
Physiological and biochemical characteristic of plants Aconitum septentrionale in South Timan communities
Ecological physiological and biochemical characteristic of Aconitum septentrionale plants, growing in different phytocenoses in South Timan in basin of Soyva river is given. Distinctions between plants in specific superficial density of leaves, the maintenance of pigments, accumulation of sugars, photosynthetic activity and efficiency, caused mainly by light and edaphic habitat conditions are revealed.
Текст научной статьи Физиолого-биохимическая характеристика растений Aconitum septentrionale в сообществах Южного Тимана
Aconitum septentrionale Koelle (аконит высокий) – многолетнее высокотравное растение с крупными пальчато-рассеченными листьями и синими цветками [7]. Вид широко распространен в таежной зоне европейского Северо-Востока, встречается в лесах, на опушках, лугах. Подземные и надземные части растений содержат большое разнообразие алкалоидов. Исследования фитоценотической приуроченности и некоторых характеристик аконита в подзоне средней тайги на территории Республики Коми показали его перспективность как ресурсного вида [4]. Однако для рационального использования ресурсов данного вида важно не только оценить запасы сырья и содержание целевого продукта, необходимо более полное знание об эколого-биологических и физиологических свойствах растений.
Цель работы: выявить вариабельность физиолого-биохимических характеристик растений аконита высокого в ценопопуляциях Южного Тимана.
Материал и методы. Исследования проводили в первой половине июля 2009 г. на территории ботанического заказника «Сойвин-ский» (Южный Тиман). Район исследований относится к Атлантико-Арктической области умеренного климатического пояса и характеризуется избыточно влажным умеренноконтинентальным климатом с продолжительной
зимой, коротким и сравнительно теплым летом [1]. Среднегодовая температура составляет – 1,5оС, средняя температура самого теплого месяца – июля около +15оС. Сумма осадков за вегетационный период варьирует от 315 до 365 мм. По данным Коми республиканского центра по гидрометеорологии и мониторингу окружающей среды последняя декада июня и первая декада июля 2009 г. были прохладней обычного (на 4оС). Осадков выпало вдвое больше нормы. В целом же погодные условия вегетационного периода существенно не отличались от средних многолетних значений. Для района исследований характерны деградированные перегнойно-карбонатные, перегнойноскелетные почвы, формирующиеся на вершинах и склонах гряд и холмов. Пятнами встречаются дерново-карбонатные и дерновые щебенчатые почвы, приуроченные к выходам на поверхность известняков и других плотных пород [2]. Растения аконита отбирали в двух типичных для данного района ценопопуляциях (ЦП), локализованных на левом берегу р. Сой-ва. По сведениям, приведенным в работе [3], ЦП1 характеризуется как сероольшанник аконитовый. Древесный ярус сформирован Alnus incana, с примесью Picea obovata и Betula pu-bescens. Травяно-кустарничковый ярус густой, высота основной массы травостоя 0,9-1,2 м. Преобладают A . septentrionale , Filipendula ulmaria , Thalictrum minus . ЦП2 описана как ельник крупнотравно-злаковый. Древесный ярус представлен Picea obovata , Betula pubescens , единично Pinus sylvestris . Травянокустарничковый ярус густой, преобладают Milium effusum , A. septentrionale , Aegopodium podagraria .
О ростовой функции растений судили по накоплению и распределению сырой и сухой массы. В каждой популяции отбирали по 10-15 растений, разделяли на органы, фиксировали сухим жаром при температуре 105оС, затем высушивали при температуре 70оС и взвешивали. Морфометрические показатели листьев рассчитывали, исходя из их массы и площади. Фотосинтетические пигменты экстрагировали из листьев кипящим 100% ацетоном и определяли спектрофотометрически при длинах волн 662, 644 нм (хлорофиллы) и 470 нм (каротиноиды). О фотосинтетической активности листьев судили по показателям СО2-газообмена, измеренным при помощи ИК-газоанализатора LI-7000 (LICOR, США) при температуре +20оС в диапазоне ФАР от 0 до 1500 мкмоль/м2с. Элементный анализ осуществляли с применением метода оптической эмиссионной спектрометрии с индуктивно связанной плазмой на приборе «SPECTRO CIROS - CCD». Определения содержания азота проводили с помощью элементного CHNS-O анализатора «EA-111». Растворимые углеводы определяли методом нормально-фазовой ВЭЖХ. Газометрические измерения проводили в 6-10-кратной биологической повторности. Для аналитических определений использовали не менее трех независимых проб. Результаты обрабатывали с помощью стандартных статистических методов и компьютерных программ. В таблицах и на рисунке представлены средние арифметические величины показателей со стандартной ошибкой.
Результаты и их обсуждение. Эколого-ценотические условия оказали существенное влияние на накопление и распределение биомассы по органам растения. Общая сухая биомасса растения ЦП1 была почти в 4 раза больше по сравнению с растениями из ЦП2 (табл. 1). По данным [3] количество взрослых особей составляло в среднем 1,2 и 2,2 растений на м2 в ельнике и сероольшаннике соответственно. Хотя растения ЦП1 накапливали втрое больше подземной биомассы, чем растения ЦП2, доля корней и корневищ в общей биомассе составляла менее 25%. У растений ЦП2 этот показатель был в почти в 1,5 раза выше. Известно, что растения аконита накапливают алкалоиды не только в подземной части, но и во всех надземных органах, особенно листьях [4]. На долю листьев приходится около 30% биомассы растений. Листья аконита сильно оводнены, содержание сухих веществ в них составляет 13-15%. Существенная часть надземной биомассы растений приходится на стебли и черешки листьев. Массовая доля стеблей была в 1,5 раза выше у растений, обитающих в ЦП1, тогда как по доле листьев в биомассе растения из ЦП1 и ЦП2 достоверно не отличались. На соцветия приходилось 1% общей биомассы.
Таблица 1. Накопление и распределение биомассы по органам у растений аконита высокого
|
Цено-популяция |
Листья |
Стебли |
Черешки |
Соцветия |
Подземная часть |
Целое растение |
|
сырая масса, г/ |
растение |
|||||
|
ЦП1 |
87 |
93 |
63 |
3 |
61 |
307 |
|
ЦП2 |
23 |
24 |
22 |
1 |
21 |
91 |
|
сухая масса, г/растение |
||||||
|
ЦП1 |
13,.6 |
13,2 |
6,6 |
0,5 |
10,1 |
44,0 |
|
ЦП2 |
3,2 |
2,2 |
2,0 |
0,1 |
3,6 |
11,3 |
|
сухая масса, % от целого растения |
||||||
|
ЦП1 |
30,9 |
30,0 |
15,0 |
1,1 |
23,0 |
100 |
|
ЦП2 |
28,3 |
19,5 |
17,7 |
0,9 |
33,6 |
100 |
Обозначения: ЦП1 и ЦП2 - ценопопуляции аконита высокого соответственно в сероольшаннике аконитовом и ельнике крупно-травном злаковом. Стандартная ошибка средних величин не превышает 30%.
Растения аконита в разных ЦП отличались по основным показателям листовой поверхности (табл. 2). В ЦП1 они формировали листовую поверхность большей площади. Листья отличались более высокой удельной поверхностной плотностью (УПП). Согласно представлениям о световой адаптации, УПП листьев является параметром наиболее чувствительным к условиям произрастания. Изменения этого показателя в зависимости от поступления ФАР обеспечивает максимальный КПД газообмена листьев [6]. Судя по величине УППЛ условия освещенности растений в ЦП отличались, растения ЦП2 получали меньше света. При этом величина отношения массы листьев к массе растения, характеризующая долю биомассы растения, активно участвующую в фотосинтезе, мало зависела от местообитания растений (табл. 2).
Таблица 2 . Показатели листовой поверхности ценопопуляций аконита высокого
|
Показатели |
ЦП1 |
ЦП2 |
|
Площадь листьев, дм2/растение |
28,9±5,0* |
11,8±1,5 |
|
Удельная поверхностная плотность листьев, г/дм2 |
0,47±0,04* |
0,27±0,03 |
|
Отношение площади листьев к массе растения, дм2/г |
0,66 |
1,04 |
|
Отношение массы листьев к массе растения, г/г |
0,31 |
0,28 |
Обозначения: как в табл. 1. * различия между ЦП1 и ЦП2 существенны при Р≤0,05.
Адаптация фотосинтетического аппарата растений к недостатку света проявлялась также на уровне пигментного комплекса. В расчете на единицу сухой массы листья растений, произрастающих в ЦП2, накапливали на 30% больше хлорофиллов, чем растения в ЦП1 (табл. 3). Однако разница практически нивелировалась при пересчете данных на единицу площади листьев, так как величина УППЛ у растений ЦП1 была в 1,5 раза больше по сравнению с ЦП2. Такие же закономерности были выявлены и при анализе фонда желтых пигментов. Величина соотношения хлорофиллов и каротиноидов в пигментном комплексе растений из разных ЦП составляла около 5, что характерно для большинства видов бореальной флоры.
Таблица 3. Содержание фотосинтетических пигментов в листьях аконита высокого
|
Ценопопу-ляция |
Хлорофиллы |
Каротиноиды |
Хл/кар |
|
|
а+б \ |
а/б |
|||
|
мг/г сухой массы |
||||
|
ЦП1 |
8,48 ± 1,01 |
2,96 ± 0,11 |
1,80 ± 0,20 |
4,71 ± 0,14 |
|
ЦП2 |
13,00 ± 1,75 |
2,81 ± 0,47 |
2,42 ± 0,26 |
5,36 ± 0,17 |
|
Мг/дм2 |
||||
|
ЦП1 |
2,93±0,25 |
2,97±0,06 |
0,62±0,05 |
4,71 ± 0,14 |
|
ЦП2 |
2,47±0,30 |
2,79±0,24 |
0,46±0,06 |
5,36 ± 0,17 |
Обозначения: как в табл. 1.
Анализ содержания и состава растворимых сахаров (табл. 4) показал, что концентрация глюкозы в сухой биомассе листьев аконита существенно выше, чем фруктозы. Среди дисахаров преобладала сахароза. По сумме растворимых сахаров в расчете на единицу биомассы листьев растения из ЦП1 вдвое превосходили растения из ЦП2. Различия сохранялись и при пересчете концентрации сахаров на единицу площади листьев. У большинства видов растений умеренной зоны транспортной формой углеводов служит сахароза. Количество сахарозы в листьях растений ЦП1 было в 1,5 раза выше, чем в ЦП2. Эти различия проявлялись сильнее, если концентрацию сахарозы относили к единице площади листьев.
Таблица 4. Содержание растворимых сахаров в листьях аконита высокого
|
Сахара |
ЦП1 |
ЦП2 |
||
|
мг/г сухой массы |
мг/дм2 |
мг/г сухой массы |
мг/дм2 |
|
|
фруктоза |
6,89±2,87 |
3,2±0,3 |
2,47±0,40 |
0,7±0,1 |
|
глюкоза |
63,92±6,16 |
30,1±2,8 |
15,16±1,60 |
4,1±0,4 |
|
моносахара |
*70,81±6,80 |
*33,3±3,2 |
17,63±1,65 |
4,8±0,42 |
|
сахароза |
61,02±2,38 |
28,7±3,0 |
42,01±2,45 |
11,4±1,5 |
|
мальтоза |
7,27±0,80 |
3,4±0,3 |
13,03±1,60 |
3,5±0,4 |
|
дисахара |
**68,29±2,51 |
*32,1±2,1 |
55,04±2,92 |
14,9±0,8 |
|
сумма сахаров |
*139,10±7,24 |
*65,4±3,4 |
72,67±3,36 |
26,3±1,2 |
Обозначения: как в табл. 1. * различия между ЦП1 и ЦП2 существенны при Р≤0.01, при ** Р≤0.05.
Элементный состав растений видоспецифичен и отражает условия почвенного питания растений. Как видно из данных табл. 5, наиболее обогащены минеральными элементами ассимилирующие органы – листья. В них выше, чем других частях растений, концентрация азота, фосфора и особенно кальция. Содержание калия больше в органах, осуществляющих транспортные функции. Сравнение данных показало, что органы аконита, произрастающего в ельнике, накапливали больше калия и фосфора, тогда как различия в содержании азота и других элементов между растениями ЦП1 и ЦП2 были слабо выражены. Концентрация микроэлементов в разных частях растений была неодинаковой. В большинстве случаев их содержание в органах подземной части растений было выше, чем в листьях. Следует отметить, что корни и корневища концентрировали в значительных количествах натрий, железо, марганец, цинк и медь. Что касается влияния эколого-ценотических условий на накопление определенных микроэлементов, то эффекты были разнонаправленными. Так, содержание натрия, марганца и меди было выше в биомассе органов растений ЦП2 по сравнению с ЦП1. В то же время листья растений ЦП1 накапливали больше цинка и железа, чем листья растений ЦП2.
Таблица 5. Содержание основных минеральных элементов в биомассе аконита высокого, мг/г сухой массы
|
Часть растения |
N 1 |
K 1 |
P 1 |
Ca 1 |
Mg |
Fe |
|
ЦП 1 |
||||||
|
листья |
38,0±4,0 |
12±0,5 |
2,8±0,9 |
26±8,0 |
8,5±2,5 |
0,9±0,2 |
|
черешки |
16,6±2,4 |
21±8,0 |
1,9±0,6 |
8,7±2,6 |
6,8±2,0 |
0,2±0,07 |
|
стебли |
14,6±2,2 |
13±5,0 |
1,8±0,5 |
4,8±1,4 |
2,6±0,8 |
0,1±0,03 |
|
подземная часть |
22,0±3,0 |
9±3,0 |
2,3±0,7 |
8,4±2,5 |
3,3±1,0 |
0,4±0,10 |
|
ЦП 2 |
||||||
|
листья |
35,0±4,0 |
34±13 |
6,1±1,8 |
21±1,6 |
5,4±1,6 |
0,2±0,04 |
|
черешки |
12,9±1,9 |
48±19 |
5,7±1,7 |
6,0±1,8 |
3,4±1,0 |
0,1±0,02 |
|
стебли |
15,0±2,0 |
42±17 |
5,4±1,6 |
4,7±1,4 |
3,2±1,0 |
0,2±0,04 |
|
подземная часть |
24,0±3,0 |
17±7 |
11±3,0 |
7,2±2,2 |
5,3±1,6 |
1,8±0,5 |
Обозначения: как в табл. 1.
Продуктивность и устойчивость растений в значительной степени зависят от того, насколько успешно растения реализуют функцию фотосинтеза. Наши данные показывают, что скорость фотосинтеза сильно зависит от освещенности (рис. 1). Поглощение СО2 линейно возрастало с повышением ФАР в области от 0 до 100-120 мкмоль/м2с. При освещенности свыше 300-400 мкмоль/м2с происходит насыщение фотосинтеза светом. В области низкой освещенности (на линейном участке световой кривой) различия в скорости фотосинтеза листьев растений ЦП1 и ЦП2 незначительны. В области светового насыщения скорость фотосинтеза единицы массы листьев растений обоих ЦП составляла около 15 мг СО2/(г ч). Фотосинтетическая активность листьев растений, произрастающих в сероольша-нике, выше по сравнению с листьями растений, обитающих в ельнике. Повышение фотосинтетической мощности листьев ЦП1 обусловлено их более высокой УППЛ (табл. 2). По нашим подсчетам при умеренной ФАР (около 500 мкмоль/м2с или 125 Вт/м2) одно растение аконита в ЦП2 за час может ассимилировать до 50 мг СО2, что эквивалентно образованию 70 мг глюкозы. За это же время растение аконита ЦП1 способно ассимилировать почти в 4 раза больше СО2. Данным фактом в значительной степени объясняются различия в продуктивности растений ЦП1 и ЦП2 (табл. 1). С этим также связана и разница в содержании сахаров в листьях (табл. 4).
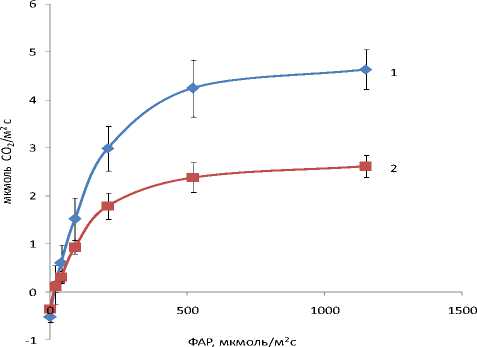
Рис. 1. Световая зависимость фотосинтеза листьев растений Aconitum septentrionale , обитающих в сероольшаннике аконитовом (1) и ельнике крупнотравно-злаковом (2) на Южном Тиммане
Выводы: эколого-ценотические условия оказывают существенное влияние на физиолого-биохимические характеристики и продукционный процесс растений. Установлена положительная связь между фотосинтетической активностью, удельной поверхностной плотностью листьев и накоплением биомассы растениями аконита высокого, обитающими в разных сообществах на Южном Тимане.
Работа выполнена в рамках проекта «Состояние ресурсов полезных растений европейского северо-востока России: мониторинг и разработка биотехнологических подходов по рациональному использованию и воспроизводству» и Программы Отделения биологических наук РАН «Биологические ресурсы России: оценка состояния и фундаментальные основы мониторинга». Исследования поддержаны грантом УрО РАН (№ 09-Т-4-1002)
Список литературы Физиолого-биохимическая характеристика растений Aconitum septentrionale в сообществах Южного Тимана
- Агроклиматические ресурсы Коми АССР. -Л.: Гидрометеоиздат, 1973. 136 с.
- Атлас почв Республики Коми/Под ред. Г.В. Добровольского, А.И. Таскаева, И.В. Забоевой. -Сыктывкар, 2010. 356 с.
- Паршина, Е.И. Биология и ресурсы алкалоидосодержащего вида Aconitum septentrionale Koelle в сообществах таежной зоны Европейского Северо-Востока России. Автореф. дис.... канд. биол. наук. -Сыктывкар. 2009. 18 с.
- Паршина, Е.И. Фитоценотическая приуроченность и ресурсные характеристики Aconitum septentrionale (Ranunculaceae) в подзоне средней тайги на северо-востоке европейской России/Е.И. Паршина, И.Ф. Чадин, С.О. Володина и др.//Растительные ресурсы. 2009. № 3. С. 60-67.
- Растительные ресурсы России: Дикорастущие цветковые растения, их компонентный состав и биологическая активность. Т.1. Семейства Magnoliaceae -Juglandaceae, Ulmaceae, Moraceae, Cannabaceae, Urticaceae/Отв.ред. А.Л. Буданцев. -СПб.-М.: Товарищество научных изданий КМК, 2008. 421 с.
- Тооминг, Х.Г. Экологические принципы максимальной продуктивности посевов. -Л.: Гидрометеоиздат, 1984. 264 с.
- Флора северо-востока Европейской части СССР/Под. ред. А.И.Толмачева. Т.3. Семейства Nymphaeaceae -Hippuridaceae. -Л.: Наука, 1976. 293 с.
