Физиолого-биохимические ответы растений сахарной свеклы (Beta vulgaris L.), зараженных грибом Alternaria alternata, на применение микробных препаратов на основе Bacillus subtilis
, зараженных грибом Alternaria alternata, на применение микробных препаратов на основе Bacillus subtilis")
Автор: Ласточкина О.В., Пусенкова Л.И., Юлдашев Р.А., Ильясова Е.Ю., Aliniaeifard S.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Иммунитет и болезни растений
Статья в выпуске: 5 т.53, 2018 года.
Бесплатный доступ
Болезни листьев, вызванные патогенными грибами рода Alternaria, значительно снижают качество и продуктивность корнеплодов сахарной свеклы ( Beta vulgaris L.). К перспективным агентам биоконтроля болезней растений относятся биопрепараты на основе бактерий Bacillus subtilis в связи с их способностью стимулировать рост и активировать иммунитет растений к биотическим стрессорам. К началу исследования мы не располагали сведениями о влиянии B. subtilis на физиологические характеристики растений сахарной свеклы, пораженных альтернариозом. Нами впервые представлены данные, которые свидетельствуют, что такие биопрепараты препятствуют снижению фотосинтеза, индуцированному возбудителем альтернариоза, и инициируют защитные реакции у растений сахарной свеклы, включая синтез ингибиторов гидролитических ферментов и при этом уменьшая количество пролина. Высокую эффективность проявил испытанный впервые новый штамм B. subtilis 10-4, выделенный из почв в Республике Башкортостан. Цель работы заключалась в оценке влияния биопрепаратов фитоспорин-М, витаплан и штамма B...
Фотосинтетические пигменты, гидролазы, сахар, пролин, сахароза, alternaria аlternata, устойчивость, продуктивность, сахарная свекла
Короткий адрес: https://sciup.org/142216602
IDR: 142216602 | УДК: 633.41:632.4:578.1:58.071 | DOI: 10.15389/agrobiology.2018.5.958rus
Effect of bacillus subtilis based micrrobials on physiological and biochemical parameters of sugar beet (Beta vulgaris L.) plants infected with Alternaria alternata
Phytopathogenic Alternaria fungi are economically important causative agents of sugar beet ( Beta vulgaris L.) leaf diseases which significantly reduce root yield and quality. Promising agents for plant disease biocontrol are Bacillus subtilis based biologicals due to the ability to stimulate plant growth and immunity to many biotic stressors. Starting our experiments, we could not find publications on B. subtilis effects towards physiological parameters of sugar beet plants affected by Alternaria. This paper is the first to report that B. subtilis -based biologicals including novel Bashkirian isolate B. subtilis 10-4 prevent a decrease in leaf photosynthetic activity in sugar beet plants affected by A. alternata, activate hydrolytic enzyme inhibitors, suppress proline production, and increase sugar content in roots. Our objective was to estimate effects of Fitosporin-M, Vitaplan, and endophytic strain B. subtilis 10-4 on leaf photosynthetic pigments (chlorophyll a, b and carotenoids), leaf area index, activity of hydrolases (proteases and amylases) and their inhibitors, as well as proline and sugar levels in leaves, root level of sucrose, and productivity in healthy plants as compared to those artificially infected with A...
Текст научной статьи Физиолого-биохимические ответы растений сахарной свеклы (Beta vulgaris L.), зараженных грибом Alternaria alternata, на применение микробных препаратов на основе Bacillus subtilis
Болезни листьев, вызванные патогенными грибами рода Alternaria , значительно снижают продуктивность и качество растений сахарной свеклы ( Beta vulgaris L.) — одной из наиболее важных сахароносных культур, которая служит источником сырья для сахарной, пищевой, кондитерской, 958
спиртовой промышленности, производства биоэтанола, удобрений, кормов для животных (1). У пораженных растений развивается альтернариоз, для которого характерно образование пятнистостей на поверхности листьев (2). Нарушаются физиологические функции, изменяются анатомо-морфологические показатели, снижается продуктивность, отмирают отдельные части растений, происходит их полная гибель (3). Преждевременные потери ассимиляционной площади листового аппарата, вызванные альтер-нариозом, приводят к утрате пластических веществ корней, затраченных на новообразование листьев, замедляется нарастание массы корнеплодов, в них снижается содержание сахарозы (2).
Преимущества использования для оздоровления растений биологических препаратов в сравнении с химическими средствами защиты заключаются в экологической безопасности и системном иммуномодулирующем действии (3, 4). К перспективным агентам биоконтроля и активаторам иммунитета растений относятся микробные препараты на основе бактерий Bacillus subtilis , обладающих антагонизмом к патогенным микроорганизмам и положительно влияющих на продуктивность культур (4-6). Ростостимулирующий и протекторный эффекты этих препаратов показаны на многих видах растений (7-9) и в отношении различных стрессовых факторов биотической и абиотической природы (10-12). Считается, что такое действие обусловлено способностью B. subtilis продуцировать биологически активные вещества (инсектицидные и антимикробные компоненты, фитогормоны, сидерофоры и хелаторы) (13-15), снижать содержание этилена в растениях, улучшать фиксацию азота, усвояемость макро- и микроэлементов (16), запускать механизмы системной устойчивости растений в ответ на стрессы (17) посредством активации салицилат- и жасмонат-зависимых сигнальных путей (18-20).
Важную роль в индукции устойчивости растений к патогенам играют гидролитические ферменты (амилазы, пектиназы, протеазы) и их ингибиторы (21-23). На модельных растениях картофеля и сахарной свеклы показано, что внесение биопрепаратов на основе B. subtilis способствует активации синтеза ингибиторов протеаз и защищает растения от проникновения и развития патогенных микроорганизмов (6). О развитии защитных реакций растений к стрессам разной природы можно также судить по степени накопления в них пролина и сахаров, служащих маркерами формирования устойчивости в экстремальных ситуациях (24-26). В то же время, несмотря на значительный объем экспериментальных данных, последовательность развития защитных механизмов, индуцируемых B. subtilis , до конца не ясна. Начиная свои исследования, мы не располагали сведениями о влиянии B. subtilis на фотосинтетическую активность листьев сахарной свеклы как интегральную характеристику физиологического состояния всего растения и о характере изменений в содержании пролина и сахаров в листьях в условиях заражения возбудителями альтернариоза.
В результате проведенных исследований мы впервые выявили, что внесение биологических препаратов на основе Bacillus subtilis препятствует снижению фотосинтетической активности листового аппарата сахарной свеклы, индуцированному возбудителем альтернариоза, и инициирует защитные реакции, включающие активацию ингибиторов гидролитических ферментов, повышение содержания пролина и сахаров. Это снижает повреждающее действие патогенного гриба A. alternata на растения сахарной свеклы и способствует формированию более крупных корнеплодов.
Цель работы заключалась в оценке влияния биопрепаратов фито-спорин-М, витаплан и штамма Bacillus subtilis 10-4 на физиолого-биохи- мические показатели и продуктивность сахарной свеклы при заражении Alternaria alternata.
Методика . Исследования выполняли на растениях сахарной свеклы Beta vulgaris L. сорта Кампаи (ООО «АгроСем-Инвест», г. Краснодар). В опытах использовали биопрепараты фитоспорин-М ( B. subtilis 26D, НВП «БашИнком», г. Уфа) (П, 30 г/10 л), витаплан ( B. subtilis 2604D + B. sub-tilis 2605D, ЗАО «Агробиотехнология», Россия) (СП, 20 г/га) и новый штамм B. subtilis 10-4, выделенный в Башкирском НИИ сельского хозяйства (105 КОЕ/мл) (8). Растения опрыскивали суспензиями биопрепаратов 2-кратно — в фазу 2-3 пар и 4-6 пар настоящих листьев. Расход рабочей жидкости составлял 300 л/га.
Полевые опыты проводили в Предуральской степной зоне Республики Башкортостан (OOO «Чишмы агроинвест») в 2013 году на мелких делянках (5 м2). Почва на опытном поле — чернозем выщелоченный (рН 5,25), Hg — 5,50 мг-экв/100 г почвы, содержание гумуса — 8,69 %, калия и фосфора — соответственно 29,0 и 23,0 мг/100 г. Сахарную свеклу высаживали в сроки, общепринятые для региона, появление всходов отмечали на 12-14-е сут. Всходы искусственно заражали, нанося на листья по 100 мкл суспензии гриба A. alternata (106 КОЕ/мл). Степень развития болезни оценивали визуально в течение вегетации по 5-балльной шкале: 0 баллов — отсутствие симптомов, 1 балл — поражение от 1 до 25 % площади листа, 2 балла — от 26 до 50 %, 3 балла — от 51 до 75 %, 4 балла — более 75 %.
В течение вегетации трижды (фазы 4-6 пар настоящих листьев, смыкания листьев в рядках, технической спелости) отбирали контрольные (необработанные, здоровые) и опытные (обработанные биопрепаратами и зараженные A. alternata ) целые растения или отдельно корнеплоды и побеги для оценки физиолого-биохимических показателей.
Содержание хлорофиллов a и b определяли согласно S.W. Jefferey с соавт. (27). Образцы листьев взвешивали, измельчали, перетирали в ступке с добавлением углекислого кальция и 90 % ацетона (из расчета 0,05 г листьев на 10 мл ацетона), полученный экстракт фильтровали. Оптическую плотность растворов экстрагированных пигментов измеряли при λ = 436 нм и λ = 680 нм на спектрофотометре UV-2401PC («Shimadzu», Япония). Концентрацию каротиноидов в суммарной вытяжке пигментов вычисляли по формуле P. Wettstein (28).
Образование пролина в листьях оценивали по методике L.S. Bates с соавт. (29) в модификации Л.Г. Калинкиной (30). Для этого брали по 2 г исследуемого материала и заливали 2,5 мл кипящей дистиллированной воды. Пробирки доводили до кипения в водяной бане и охлаждали. Затем пробирки с 2 мл холодной пробы, 2 мл нингидринового реактива, 2 мл ледяной уксусной кислоты помещали в водяную баню, кипятили 1 ч и охлаждали. Интенсивность окрашивания комплекса пролина с нингидрином определяли при λ = 522 нм на спектрофотометре СФ-26 («ЛОМО», Россия). Содержание пролина устанавливали по калибровочной кривой, используя стандартные растворы химически чистого L-пролина («Sigma Aldrich», США).
Ферментативную активность протеиназ, амилаз и их ингибиторов определяли спектрофотометрическим методом (21, 31), накопление сахара в листьях — фотометрическим методом с применением 2,4-динитрофенола по ГОСТ Р 51636-2000, количество сахарозы в корнеплодах — методом холодного водного дигерирования с использованием поляриметра П161-М (Россия) (32). Площадь поверхности листьев измеряли с помощью фотопланиметра, массу надземной части растений и массу корнеплодов — ве- совым методом (33).
Все опыты проводили в 3-4 биологических и 4-5 аналитических повторах. Статистическую обработку выполняли с использованием компьютерной программы STATISTICA 6.0 («StatSoft, Inc.», США). На рисунках и в таблицах представлены средние значения ( M ) и их стандартные отклонения (±SD) при Р = 0,95.
Результаты . Альтернариоз поражает листовую поверхность растений, образуя пятнистости, и приводит к уменьшению фотосинтетической поверхности листьев (2). Фотосинтез — главный процесс при формировании продуктивности растений, от его интенсивности зависит общая биологическая урожайность культур (34). В свою очередь, содержание основных фотосинтетических пигментов (хлорофиллов а, b и каротиноидов) служит одним из косвенных индексов фотосинтетической активности и важнейшим биохимическим показателем растения, определяющим интенсивность фотосинтеза (24, 34). В наших опытах инфицирование сахарной свеклы A. alternata приводило к снижению в листьях содержания хлорофилла а (до 1,5-кратного) и b (до 1,2-кратного) по сравнению со здоровыми растениями (рис. 1), что свидетельствует о нарушении процесса фотосинтеза и уменьшении фотосинтетической деятельности растений.
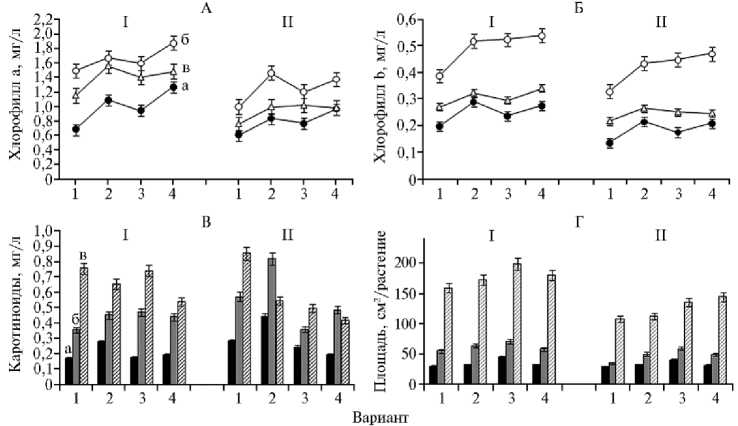
Рис. 1. Содержание хлорофилла а (А) , хлорофилла b (Б) , каротиноидов (В) и площадь листовой пластинки (Г) у здоровых (I) и инфицированных Alternaria alternata (II) растений сахарной свеклы Beta vulgaris L. сорта Кампаи при использовании биопрепаратов: 1 — контроль, 2 — фитоспорин-М, 3 — витаплан, 4 — штамм Bacillus subtilis 10-4; а — первая обработка, б — вторая обработка, в — уборка урожая (OOO «Чишмы агроинвест», Республика Башкортостан, 2013 год).
Инокуляция штаммом B. subtilis 10-4 и обработка биопрепаратами фитоспорин-М и витаплан восстанавливала фотосинтетическую деятельность растений. Так, 2-кратная обработка биопрепаратами предотвращала индуцированное стрессом снижение содержания хлорофиллов а и b во всех вариантах опыта (см. рис. 1, А, Б). Содержание каротиноидов в листьях при заражении A. alternata увеличивалось (см. рис. 1, В). Обработка витапланом и штаммом B. subtilis 10-4 способствовала снижению их количества, тогда как после двух обработок фитоспорином-М наблюдалось значительное накопление каротиноидов, превышавшее контрольные значения. Однако к уборке содержание каротиноидов снижалось и было сопоставимо с таковым в вариантах, в которых применялись витаплан и штамм 10-4 (см. рис. 1, В). Следует отметить, что в незараженных растениях 2-кратная обработка биопрепаратами хоть и приводила к незначительному повышению количества каротиноидов, но к уборке этот показатель был ниже, чем в контрольном варианте (см. рис. 1, В).
Внесение биопрепаратов в нормальных условиях произрастания стимулировало фотосинтетическую деятельность растений, вероятно, за счет увеличения содержания физиологически активного хлорофилла a. Действительно, результаты, полученные при оценке площади поверхности листьев (см. рис. 1, Г), коррелировали с данными о влиянии исследуемых препаратов на содержание хлорофиллов a и b. Растения, обработанные препаратами, во всех вариантах в течение всего вегетационного периода характеризовались значительно большей площадью листовой пластинки как в норме, так при заражении A. alternata (см. рис. 1, Г).
Очевидно, что выявленное увеличение содержания фотосинтетических пигментов при использовании биопрепаратов в условиях инфицирования A. alternata (см. рис. 1, А, Б, В), помимо их непосредственной роли в процессе фотосинтеза и увеличении размеров листьев (см. рис. 1, Г), может вносить вклад в развитие защитных реакций растений (34). В частности, каротиноиды выполняют фотопротекторную и антиоксидантную функции (35-38), предотвращая повреждения, вызываемые образованием синглетного кислорода и триплетного хлорофилла (37). К тому же они могут принимать энергию возбуждения триплетного хлорофилла, а затем рассеивать ее в виде тепла или гасить молекулы синглетного кислорода (38, 39). Однако следует отметить, что, несмотря на очевидную роль каротиноидов в антиоксидантной защите растений, данные об изменении их содержания при действии стрессовых воздействий весьма противоречивы (22, 24, 36). Подобные эффекты могут объясняться, с одной стороны, индукцией образования каротиноидов под действием стресс-фактора, с другой — его усиленной деградацией при сильном стрессе (22).
Вместе с тем при формировании защитных реакций растений к патогенам значительную роль играют гидролитические ферменты (протеазы, амилазы) и их ингибиторы (21). При инфицировании A. alternata происходило повышение активности протеиназ и амилаз (рис. 2, Б, Г) в листьях сахарной свеклы, что свидетельствует об интенсивном развитии патогена в тканях растений. Вероятно, этот процесс был обусловлен изменениями метаболизма растения-хозяина под воздействием патогена, а также секрецией самим патогеном гидролитических ферментов, способных мацерировать ткани и разрушать компоненты клеточной стенки, что позволяет патогену преодолевать естественную резистентность растения-хозяина. Растения, обработанные штаммом B. subtilis 10-4, фитоспорином-М и вита-планом характеризовались снижением активности гидролаз как в зараженных, так и в здоровых листьях (см. рис. 2, А-Г), причем наибольшее снижение гидролитической активности наблюдалось в вариантах с использованием препарата фитоспорин-М и штамма B. subtilis 10-4.
Значимый вклад в регуляцию активности гидролитических ферментов вносят белковые ингибиторы растения, подавляющие активность собственных и чужеродных ферментов, в частности патогенных грибов и бактерий (23, 40). В ответ на инфицирование A. alternata в листьях сахарной свеклы активность ингибиторов гидролаз снижалась (см. рис. 2, Е, З), вследствие чего, вероятно, уменьшалась сопротивляемость растений к действию ферментов патогена и его распространению в тканях. Обработка исследуемыми препаратами, напротив, способствовала увеличению активности ингибиторов гидролаз (см. рис. 2, Е, З), что указывает на индуциро-962
вание у растений под их влиянием защитных реакций против А. alternatа . Следует отметить, что максимальную активацию защитных белков вызывало применение штамма B. subtilis 10-4 и фитоспорина-М.
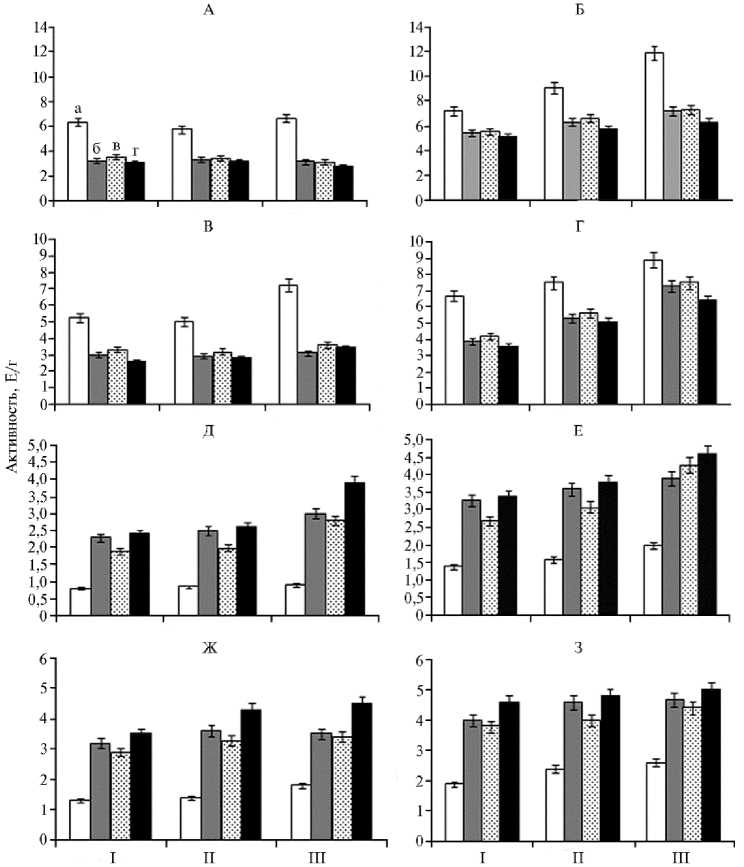
Рис. 2. Активность протеиназ (A, Б) , амилаз (В, Г) , ингибиторов протеиназ (Д, Е ) и ингибиторов амилаз (Ж, З) в листьях у здоровых (А, В, Д, Ж) и инфицированных Alternaria alternata (Б, Г, Е, З) растений сахарной свеклы Beta vulgaris L. сорта Кампаи при использовании биопрепаратов: а — контроль, б — фитоспорин-М, в — витаплан, г — штамм Bacillus subtilis 10-4; I — первая обработка, II — вторая обработка, III — уборка урожая (OOO «Чишмы агроинвест», Республика Башкортостан, 2013 год).
Важными биохимическими маркерами формирования устойчивости могут выступать накопление в растительной массе сахара и пролина (31). Здоровые растения B. vulgaris постепенно накапливали сахар в листьях в течение всей вегетации (рис. 3, А), что было вполне характерно и согласовывалось с имеющимися в литературе данными (41). Наибольшую скорость накопления сахара наблюдали на первоначальных этапах роста, когда растение энергично формировало листья и корни, и замедлялась к концу образования третьей пары настоящих листьев. По-видимому, это было связано с тем, что в фазу смыкания рядков прирост листьев замедлял- ся, происходило интенсивное утолщение и формирование корнеплодов, сопровождавшееся продолжением накопления сахаров в них.
Инфицирование A. alternata приводило к резкому повышению содержания сахара в листьях по сравнению с показателями у контрольных здоровых растений, что, по-видимому, выполняет протекторную роль и позволяет растениям продолжать рост в стрессовых условиях (см. рис. 3, Б). Этому могут способствовать свойства моносахаридов, связанные с повышением стабильности биомембран, антиденатурационным воздействием на белки и антиоксидантным эффектом (42). Кроме того, накапливающиеся углеводы помогают поддерживать осмотический статус клеток (33).
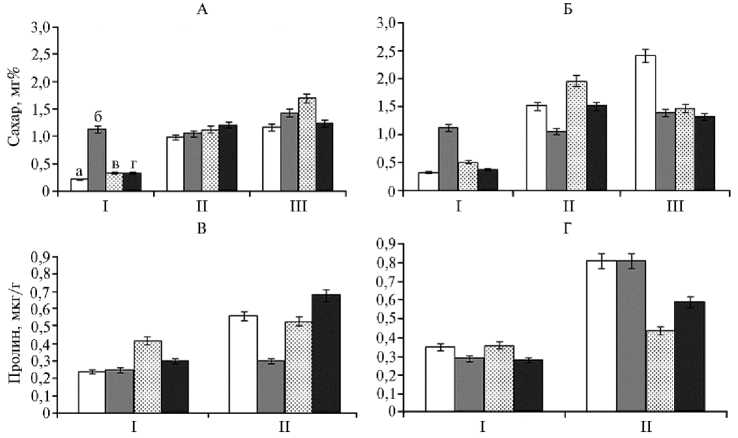
Рис. 3. Содержание сахара (А, Б) и пролина (В, Г) в листьях у здоровых (А, В) и инфицированных Alternaria alternata (Б, Г) растений сахарной свеклы Beta vulgaris L. сорта Кампаи при использовании биопрепаратов: а — контроль, б — фитоспорин-М, в — витаплан, г — штамм Bacillus subtilis 10-4; I — первая обработка, II — вторая обработка, III — уборка урожая (OOO «Чишмы агроинвест», Республика Башкортостан, 2013 год).
Один из наиболее многофункциональных стрессовых метаболитов растений — аминокислота пролин, выполняющая роль не только осмолита и антиоксиданта (24, 43), но и низкомолекулярного шаперона (40), участвующего в поддержании нативной структуры ферментов (24). Во многих исследованиях сообщалось о повышении содержания пролина в растениях в ответ на воздействие стрессов различной природы и о его значении как фактора, важного для выживания растений в экстремальных ситуациях (24-26). Однако данных об изменениях содержания пролина в растениях сахарной свеклы в условиях инфицирования A. alternata и использования препаратов на основе B. subtilis мы в доступной литературе не обнаружили.
Инфицирование сахарной свеклы A. alternata приводило к значительному повышению содержания в растениях пролина (см. рис. 3, Г). В то же время обработка фитоспорином-М, витапланом и штаммом B. sub-tilis 10-4 способствовала предотвращению его накопления, индуцированного стрессом (см. рис. 3, Г). Следует отметить, что под влиянием биопрепаратов в здоровых растениях также наблюдалось незначительное повышение количества пролина, что дополнительно указывает на важную роль этого агента в формировании индуцированной устойчивости к возбудителю альтернариоза (см. рис. 3, В).
Совокупным показателем характера физиолого-биохимических про- цессов за весь период вегетации могут служить данные о внешнем состоянии растений и продуктивности корнеплодов. В нашем опыте искусственное заражение растений грибом A. alternata приводило к постепенному увеличению площади пораженной альтернариозом листовой поверхности. Так, к уборке урожая ее доля достигала 75 % и более (4 балла), в то время как у растений, обработанных штаммом B. subtilis 10-4, фитоспорином-М, витапланом, она не превышала 35 % (2 балла). Наилучший эффект отмечали в вариантах с использованием штамма B. subtilis 10-4 и биопрепарата фитоспорина-М. В этих случаях степень развития болезни не превышала соответственно 25 и 30 %. Вместе с тем 2-кратная обработка биопрепаратами приводила к достоверному увеличению средней массы надземной части здоровых растений в 1,8-2,7 раза и средней массы корнеплодов — в 1,6-2,3 раза в зависимости от варианта опыта (табл.). Обработка биопрепаратами предотвращала стресс-индуцированное снижение показателей продуктивности корнеплодов в условиях заражения A. alternata и способствовала сохранению роста массы листьев и корнеплодов, сопоставимого с таковым у здоровых растений.
Накопление массы листьев и корнеплодов у растений сахарной свеклы ( Beta vulgaris L.) сорта Кампаи под влиянием микробных препаратов на основе Bacillus subtilis в норме и при инфицировании Alternaria alternata
|
Вариант |
Масса наземной части, г |
Масса корнеплодов, г |
||||
|
I обработка |
II обработка |
уборка урожая |
I обработка |
II обработка |
уборка урожая |
|
|
Контроль |
4,45±0,19 |
Зд 14,67±0,77 |
оровые 165,20±2,65 |
0,52±0,09 |
4,33±0,30 |
550,80±10,41 |
|
Фитоспорин-М |
6,11±0,30 |
27,96±0,82 |
304,80±2,32 |
0,92±0,19 |
10,42±0,48 |
971,00±11,89 |
|
Витаплан |
7,75±0,51 |
14,25±0,46 |
425,40±3,01 |
1,17±0,28 |
4,94±0,12 |
970,60±13,03 |
|
Bacillus subtilis 10-4 |
11,00±0,42 |
36,70±0,91 |
336,40±4,69 |
1,80±0,12 |
14,12±0,22 |
1142,40±12,62 |
|
A. alternata |
6,86±0,22 |
Зар 14,14±1,13 |
аженные 71,20±1,78 |
1,32±0,09 |
3,37±0,21 |
276,30±4,11 |
|
Фитоспорин-М |
8,84±0,14 |
15,61±0,91 |
182,00±1,99 |
1,81±0,12 |
7,11±0,29 |
463,00±5,33 |
|
Витаплан |
4,68±0,49 |
11,84±0,70 |
127,40±2,06 |
0,64±0,11 |
3,98±0,15 |
502,20±5,09 |
|
Bacillus subtilis 10-4 |
11,21±0,30 |
25,70±1,42 |
209,40±2,57 |
2,72±0,23 |
11,66±0,23 |
607,20±4,95 |
Помимо положительного влияния на интенсивность ростовых процессов и накопление биомассы, 2-кратное внесение биопрепаратов обеспечило получение корнеплодов с более высоким содержанием сахарозы в сравнении с контролем и у здоровых, и у зараженных растений. Так, к моменту уборки урожая в контрольном варианте корнеплоды содержали 16,1 % сахарозы, в опытных вариантах — от 17,9 до 19,0 %. Максимальное количество сахарозы было зафиксировано в вариантах с 2-кратной обработкой посевов фитоспорином-М и штаммом B. subtilis 10-4. В условиях заражения A. alternata корнеплоды растений, обработанных препаратами, во всех вариантах характеризовались повышенным содержанием сахарозы в сравнении с необработанными.
Таким образом, препараты фитоспорин-М, витаплан и штамм Bacillus subtilis 10-4 способствуют усилению синтеза фотосинтетических пигментов (хлорофилла a, b и каротиноидов), увеличению активности ингибиторов гидролаз в листьях и снижению стресс-индуцированного накопления пролина и сахара, оказывая защитный эффект в условиях заражения растений сахарной свеклы грибом Alternaria alternata . При обработке микробными препаратами на основе B. subtilis как у здоровых растений, так и в случае заражения A. alternatа повышалась масса корнеплодов и содержание в них сахарозы . Наиболее эффективными оказались варианты с 2-кратным применением фитоспорина-М и штамма B. subtilis 10-4, в которых негативное влияние фитопатогенного гриба A. alternata на растения
максимально нивелировалось и были получены корнеплоды с наибольшей массой и содержанием сахарозы.
Список литературы Физиолого-биохимические ответы растений сахарной свеклы (Beta vulgaris L.), зараженных грибом Alternaria alternata, на применение микробных препаратов на основе Bacillus subtilis
- Шамилев Р.В., Шамилев С.Р. Оценка и анализ динамики и эффективности производства некоторых растениеводческих культур в РФ. Современные проблемы науки и образования, 2011, 6: 1-7.
- Стогниенко О.И., Селиванова Г.А. Болезни сахарной свеклы, их возбудители. Воронеж, 2008.
- Завалин А.А. Биопрепараты, удобрения и урожай. М., 2005.
- Berg G. Plant-microbe interactions promoting plant growth and health: perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biot., 2009, 84(1): 11-18 ( ) DOI: 10.1007/s00253-009-2092-7
- Perez-Garcia A., Romero D., de Vicente A. Plant protection and growth stimulation by microorganisms: biotechnological applications of Bacilli in agriculture. Curr. Opin. Biotech., 2011, 22: 187-193 ( ) DOI: 10.1016/j.copbio.2010.12.003
- Пусенкова Л.И., Ильясова Е.Ю., Максимов И.В., Ласточкина О.В. Повышение адаптивного потенциала посевов сахарной свеклы микробными биопрепаратами в условиях биотических и абиотических стрессов. Сельскохозяйственная биология, 2015, 50(1): 115-123 ( ) DOI: 10.15389/agrobiology.2015.1.115rus
- Esitken A., Yildiz H.E., Ercisli S., Donmez M.F., Turan M., Gunes A. Effects of plant growth promoting bacteria (PGPB) on yield, growth and nutrient contents of organically grown strawberry. Scientia Horticulturae, 2010, 124: 62-66 ( ) DOI: 10.1016/j.scienta.2009.12.012
- Lastochkina O., Pusenkova L., Yuldashev R., Babaev M., Garipova S., Blagova D., Khairullin R., Aliniaeifard S. Effects of Bacillus subtilis on some physiological and biochemical parameters of Triticum aestivum L. (wheat) under salinity. Plant Physiol. Bioch., 2017, 121: 80-88 ( ) DOI: 10.1016/j.plaphy.2017.10.020
- Turan M., Ekinci M., Yildirim E., Güneş A., Karagöz K., Kotan R., Dursun A. Plant growth-promoting rhizobacteria improved growth, nutrient, and hormone content of cabbage (Brassica oleracea) seedlings. Turk. J. Agric. For., 2014, 38: 327-333 ( ) DOI: 10.3906/tar-1308-62
- Abeer H., Asma A.H., Allah A., Qarawi A., Shalawi A., Dilfuza E. Impact of plant growth promoting Bacillus subtilis on growth and physiological parameters of Bassia indica (Indian Bassia) grown under salt stress. Pak. J. Bot., 2015, 47(5): 1735-1741.
- Shternshis M.V., Belyaev A.A., Shpatova T.V., Lelyak A.A. Influence of Bacillus spp. on strawberry gray-mold causing agent and host plant resistance to disease. Contemp. Probl. Ecol., 2015, 8(3): 390-396 ( ) DOI: 10.1134/S1995425515030130
- Иванчина Н.В., Гарипова С.Р. Влияние ростстимулирующих бактерий (PGPB) на продуктивность и устойчивость растений. Агрохимия, 2012, 7: 87-95.
- Gutierrez-Manero F.J., Ramos-Solano B., Probanza A., Mehouachi J., Tadeo F.R., Talon M. The plant growth promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiologia Plantarum, 2008, 111(2): 206-211 ( ) DOI: 10.1034/j.1399-3054.2001.1110211.x
- Malfanova N., Franzil L., Lugtenberg B., Chebotar V., Ongena M. Cyclic lipopeptide profile of the plant-beneficial endophytic bacterium Bacillus subtilis HC8. Arch. Microbiol., 2012, 194(11): 893-899 ( ) DOI: 10.1007/s00203-012-0823-0
- Bottini R., Cassan F., Piccoli P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl. Microbiol. Biot., 2004, 65(5): 497-503 ( ) DOI: 10.1007/s00253-004-1696-1
- Grichko V.P., Glick B.R. Amelioration of flooding stress by ACC deaminase-containing plant growth-promoting bacteria. Plant Physiol. Bioch., 2001, 39(1): 11-17 ( ) DOI: 10.1016/S0981-9428(00)01212-2
- Choudhary D.K., Johri B.N. Interactions of Bacillus sp. and plants -with special reference to induced systemic resistance (ISR). Microbiol. Res., 2009, 164: 493-513 ( ) DOI: 10.1016/j.micres.2008.08.007
- Niu D.D., Liu H.X., Jiang C.H., Wang Y.P., Wang Q.Y., Jin H.L., Guo J.H. The plant growth-promoting rhizobacterium Bacillus cereus AR156 induces systemic resistance in Arabidopsis thaliana by simultaneously activating salicylate-and jasmonate/ethylene-dependent signaling pathways. Mol. Plant Microbe. In., 2011, 24(5): 533-542 ( ) DOI: 10.1094/MPMI-09-10-0213
- García-Gutiérrez L., Zeriouh H., Romero D., Cubero J., de Vicente A., Pérez-García A. The antagonistic strain Bacillus subtilis UMAF6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate-and salicylic acid-dependent defense responses. Microb. Biotechnol., 2013, 6: 264-274 ( ) DOI: 10.1111/1751-7915.12028
- González-Gallegos E., Laredo-Alcalá E., Ascacio-Valdés J., Jasso de Rodríguez D., Hernández-Castillo F.D. Changes in the production of salicylic and jasmonic acid in potato plants (Solanum tuberosum) as response to foliar application of biotic and abiotic inductors. American Journal of Plant Sciences, 2015, 6(11): 1785-1791 ( ) DOI: 10.4236/ajps.2015.611179
- Шпирная И.А., Ибрагимов Р.И., Умаров И.А. Подавление активности гидролитических ферментов личинок колорадского жука растительными белками. Вестник БГУ, 2006, 3(11): 49-52.
- Demirevska-Kepova K., Simova-Stoilova L., Petrova Stoyanova Z., Feller U. Cadmium stress in barley: growth, leaf pigment, and protein composition and detoxification of reactive oxygen species. J. Plant Nutr., 2006, 29(3): 451-468 ( ) DOI: 10.1080/01904160500524951
- Иевлева Е.В., Ревина Т.А., Кудрявцева Н.Н., Софьин А.В., Валуева Т.А. Внеклеточные протеиназы фитопатогенного гриба Fusarium culmorum. Прикладная биохимия и микробиология, 2006, 42(3): 338-344.
- Колупаев Ю.Е., Ястреб Т.О. Физиологические функции неэнзиматических антиоксидантов растений. Вестник ХНУ. Серия Биология, 2015, 2(35): 6-25.
- Molinari H.B.C., Marur C.J., Daros E., de Campos M.K.F., de Carvalho J.F.R.P., Filho B.J.C., Pereira L.F.P., Vieira L.G.E. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): osmotic adjustment, chlorophyll fluorescence and oxidative stress. Physiologia Plantarum, 2007, 130(2): 218-229 ( ) DOI: 10.1111/j.1399-3054.2007.00909.x
- Shakirova F.M., Avalbaev A.M., Bezrukova M.V., Fatkhutdinova R.A., Maslennikova D.R., Yuldashev R.A., Allagulova Ch.R., Lastochkina O.V. Phytohormones and abiotic stress tolerance in plants/N. Khan, R. Nazar, N. Iqbal, N. Anjum (eds.). Springer, Berlin Heidelberg, 2012 ( ) DOI: 10.1007/978-3-642-25829-9
- Jefferey S.W., Humphrey G.F. New spectrophotometric equations for determining chlorophylls a, b, c1, and c2 in higher plants, algae, and natural phytoplankton. Biochem. Physiol. Pfl., 1975, 167: 191-194.
- Wettstein P. Chrofyll -letal und der submiscopische Form wechsel der Plastiden. Exp. Cell Res., 1957, 12(4): 427-431.
- Bates L.S., Waldern. R.P., Teare D. Rapid determination of free proline for water-stress studies. Plant Soil, 1973, 39: 205-207.
- Калинкина Л.Г. Накопление пролина в клетках морской и пресноводной хлореллы в зависимости от концентрации NaCl в среде и интенсивности роста водорослей. Физиология растений, 1985, 32: 42-52.
- Erlanger B.F., Kokowski N., Cohen W. The preparation and properties of two new chromogenic substrates of trypsin. Arch. Biochem. Biophys, 1961, 95: 271-278.
- Методика определения химического состава и показателей качества сахарной свеклы. Курск, 2001.
- Литвинов С.С. Методика полевого опыта в овощеводстве. М., 2011.
- Андриянова Ю.Е., Тарчевский И.А. Хлорофилл и продуктивность растений. М., 2000.
- Cuttriss A.J., Pogson B.J. Carotenoids. Plant pigments and their manipulation/K.M. Davies (ed.). CRC Press, Boca Raton, 2004.
- Gill S.S., Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch., 2010, 48: 909-930 ( ) DOI: 10.1016/j.plaphy.2010.08.016
- Edge R., Truscott G. Properties of carotenoid radicals and excited states and their potential role in biological systems. In: Carotenoids: physical, chemical, and biological functions and properties/J.T. Landrum (ed.). Kluwer, Dordrecht, 2010.
- Смоликова Г.Н., Медведев С.С. Каротиноиды семян: синтез, разнообразие и функции. Физиология растений, 2015, 62(1): 3-16.
- Jahns P., Holzwarth A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. BBA-Bioenergetics, 2012, 1817(1): 182-193 ( ) DOI: 10.1016/j.bbabio.2011.04.012
- Kim Y., Mosier N.S., Ladisch M.R. Enzymatic digestion of liquid hot water pretreated hybrid poplar. Biotechnol. Progr., 2009, 25(2): 340-348 ( ) DOI: 10.1002/btpr.137
- Пискурева В.A., Павловская Н.Е., Горькова И.В., Житникова B.C. Бактерицидные и комплексообразующие свойства пектинов плодоовощного концентрата. Пищевая промышленность, 2009, 6: 50-51.
- Карпец Ю.В., Колупаев Ю.Е. Ответ растений на гипертермию: молекулярно-клеточные аспекты. Вестник ХНУ. Серия биология, 2009, 1(16): 19-38.
- Szabados L., Savoure A. Proline: a multifunctional amino acid. Trends Plant Sci., 2010, 15(2): 89-97 ( ) DOI: 10.1016/j.tplants.2009.11.009
