Физиолого-биохимические свойства лабораторных мутантов грибов Fusarium, резистентных к флудиоксонилу

Автор: Орина А.С., Гаврилова О.П., Гагкаева Т.Ю., Арабина Е.П., Буркин А.А., Кононенко Г.П.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Технические культуры и секвестрация
Статья в выпуске: 1 т.60, 2025 года.
Бесплатный доступ
Фузариоз зерновых культур - вредоносное заболевание, вызываемое несколькими видами грибов Fusarium . Один из наиболее распространенных патогенов - гриб Fusarium graminearum , поражение которым приводит к загрязнению зерна микотоксинами, в том числе дезоксиниваленолом (ДОН) и зеараленоном (ЗЕН), опасными для здоровья человека и животных. В последнее время все чаще в возделываемом зерне обнаруживают гриб F. proliferatum , продуцирующий фумонизины (ФУМ). Обычно при защите растений от патогенов используются фунгициды, однако они могут приводить к развитию резистентности у грибов Fusarium и изменению их свойств, что увеличивает риск снижения эффективности защитных мероприятий и, как следствие, загрязнения зерна микотоксинами. В представленной работе впервые определена чувствительность генетически охарактеризованных штаммов двух видов F. graminearum и F. proliferatum , выделенных из зерна, к флудиоксонилу (ФДО) и получены лабораторные мутанты грибов, резистентные к этому действующему веществу (д.в.). Также впервые установлены достоверные отличия лабораторных мутантов F. graminearum и F. proliferatum от исходных штаммов по скорости роста, патогенности для пшеницы и токсинопродуцирующей способности. Целью работы был сравнительный анализ физиолого-биохимических свойств коллекционных штаммов грибов Fusarium graminearum и F. pro-liferatum и полученных из них в лабораторных условиях мутантов, резистентных к ФДО. Объектами исследования стали 16 моноконидиальных штаммов грибов Fusarium различного происхождения, идентифицированных по морфологическим признакам как F. graminearum и F. proliferatum , из коллекции Всероссийского НИИ защиты растений (MFG) (ВИЗР, г. Санкт-Петербург, Россия). Для проведения экспериментов штаммы предварительно выращивали на картофельно-сахарозной агаризованной среде (КСА) при 25 °С в темноте. Для молекулярно-генетической идентификации грибов секвенировали фрагмент гена фактора элонгации трансляции EF-1a (tef). В исследовании использовали фунгицид Максим, КС («Syngenta AG», Швейцария), содержащий 25 г/л ФДО, который разводили в стерильной воде до концентрации 5 г/л д.в. Для получения мутантов исходные штаммы Fusarium последовательно культивировали на КСА, содержащем увеличивающиеся концентрации фунгицида. При определении чувствительности грибов к ФДО его концентрации в питательной среде для исходных штаммов составляли 5; 0,5; 0,05 и 0,005 мг/л, для мутантов - 250; 50; 5 и 0,5 мг/л. Из колоний штаммов, выращенных на КСА, вырезали диски диаметром 4 мм и помещали на поверхность питательной среды в центр пластиковой чашки Петри диаметром 85 мм. В контрольном варианте диск помещали на поверхность КСА без добавления фунгицида. Через 3 и 5 сут инкубации соответственно штаммов F. graminearum и F. proliferatum определяли средний диаметр колонии гриба, вычитая диаметр инокуляционного диска. Концентрацию фунгицида, приводящую к 50 % подавлению роста (полумаксимальное ингибирование; IC50) рассчитывали с помощью программы Quest Graph™ IC50 Calculator. Для оценки влияния температуры на скорость роста грибы культивировали при 25 °С и 35 °С в темноте в термостате Innova 44R («Eppendorf», Германия), измеряя диаметр колоний: для F. graminearum соответственно на 3-и и 7-е сут, для F. proliferatum - на 5-е и 7-е сут. Скорость роста определяли как отношение диаметра колонии к числу суток культивирования (мм/сут). Патогенность штаммов оценивали посредством инокуляции отрезков листьев пшеницы (Triticum aestivum L.) сорта Васса. Для изучения токсинообразования штаммы грибов выращивали на автокловированном зерне пшеницы. Количество ДОН и ЗЕН в культурах F. graminearum и ФУМ в культурах F. proliferatum определяли с помощью аттестованных коммерческих иммуноферментных тест-систем (ВНИИВСГЭ, Россия). При культивировании на КСА без фунгицида диаметр колоний исходных штаммов F. graminearum варьировал от 34 до 52 мм на 3-и сут, а исходных штаммов F. proliferatum - от 48 до 64 мм на 5-е сут. Концентрации флудиоксонила (IC50), приводящие к полумаксимальному ингибированию роста мутантов F. graminearum и F. proliferatum , оказались соответственно в 900-60000 и 6-458 раз выше значений IC50 для исходных штаммов. Два штамма F. proliferatum изначально показали высокую резистентностью к ФДО и образовывали ФУМ в количестве, которое в 6 и 22 раза превышало средний показатель для исходных штаммов этого вида. Средняя скорость роста у лабораторных мутантов F. graminearum и F. proliferatum при 25 °С составила соответственно 17,2±1,0 и 12,8±0,2 мм/сут и оказалась на 2,6 и 1,6 мм/сут больше, чем средние показатели у исходных штаммов этих видов грибов. Шесть лабораторных мутантов F. graminearum были патогенными для пшеницы, и при инокуляции на отрезках листьев появлялись некрозы длиной от 14±3 до 44±1 мм, что оказалось в среднем в 1,6 раза меньше, чем у исходных штаммов гриба, тогда как два мутанта F. graminearum не вызывали некрозы в отличие от исходных культур.
Грибы fusarium, флудиоксонил, резистентность, патогенность, микотоксины
Короткий адрес: https://sciup.org/142244146
IDR: 142244146 | УДК: 632.4.01/.08::615.015.8 | DOI: 10.15389/agrobiology.2025.1.138rus
Physiological and biochemical characters of laboratory induced Fusarium mutants resistant to fludioxonil
Fusarium head blight is a harmful disease of cereals caused by Fusarium fungi. Fusarium graminearum is among the most widespread fungi in the world and is of great economic importance, which leads to grain contamination with the mycotoxins deoxynivalenol (DON) and zearalenone (ZEN) that poses danger to human and animal health. Recently, F. proliferatum , the fumonisins (FUM) producer, has been increasingly discovered in cultivated cereals. Fungicide treatment is the most popular practice for managing pathogens, but also leads to the emergence of fungicide resistance in Fusarium fungi and changes in their features. As a result, the risk of reducing the effectiveness in disease control and increasing a contamination of grain with mycotoxins exists. In this study, the sensitivity of genetically characterized F. graminearum and for the first time F. proliferatum strains isolated from grain, to fludioxonil has been determined, and the laboratory mutants of fungi resistant to this active substance have been generated. Also, for the first time, significant differences between the laboratory mutants and parent strains of F. graminearum and F. proliferatum in terms of growth rate, pathogenicity to wheat and toxin-producing ability have been established. The aim of study was the comparative analysis of physiological and biochemical characters of the parent F. graminearum and F. proliferatum strains with mutants resistant to fludioxonil obtained in the laboratory conditions. The objects of the study were 16 monoconidial Fusarium strains of different geographical origins from the collection of All-Russian Institute of Plant Protection (MFG) (VIZR, St. Petersburg, Russia), previously morphologically identified as F. graminearum and F. proliferatum . For laboratory experiments, the strains were previously grown on potato sucrose agar (PSA) at 25 °C in the dark. The identification of Fusarium strains was confirmed by sequence analysis of the fragment of gene encoding translation elongation factor EF-1a ( tef ) . The fungicide Maxim (Syngenta AG, Switzerland), containing 25 g/l fludioxonil, was diluted in sterile water to a concentration of 5 g/l of the active ingredient. To obtain F. graminearum and F. proliferatum mutants resistant to fludioxonil, the parent Fusarium strains were sequentially cultured on PSA containing increasing concentrations of the fungicide. When determining the sensitivity of fungi to fungicide, the concentrations of fludioxonil in PSA for the parent strains were 5; 0.5; 0.05 and 0.005 mg/l, and for the mutants 250; 50; 5 and 0.5 mg/l. From colonies of the parent strains and mutants grown on PSA, 4 mm discs were cut out and placed on the nutrient medium in the center of a plastic 85 mm Petri dish. In the control, the disk was placed on the surface of PSA without the fungicide. After 3- and 5-day incubation of the F. graminearum and F. proliferatum strains, respectively, the average diameter of the fungal colony was meaasured excluding the diameter of the inoculation disk. The fungicide concentration resulting in 50 % growth inhibition (half-maximal inhibition concentration; IC50) was calculated using the Quest Graph™ IC50 Calculator program. To assess the effect of temperature on the fungal growth rate, the strains were cultured at 25 °C and 35 °C in the dark in Innova 44R incubator (Eppendorf, Germany), and diameter of F. graminearum and F. proliferatum colonies were measured on days 3 and 7, and on days 5 and 7, respectively. The growth rate was determined as the ratio of the colony diameter to the number of days of culture (mm/day). The pathogenicity of the parent strains and mutants was assessed by the detached leaf assay, according to sizes of necrosis caused by fungal strains after inoculation of the wheat (Triticum aestivum L., cv. Wassa). To assess toxin production, fungal strains were grown on autoclaved wheat grain. The determination of DON and ZEN in F. graminearum cultures and FUM in F. proliferatum cultures was performed using the certified commercial enzyme immunoassay test systems (VNIIVSGE, Russia). When cultured on PSA without fungicide, the colony diameter of the parent F. graminearum strains varied from 34 to 52 mm on day 3, and that of the parent F. proliferatum strains were from 48 to 64 mm on day 5. The IC50 concentrations of fludioxonil resulting in half-maximal growth inhibition for F. graminearum and F. proliferatum mutants were 900-60000 and 6-458 times higher than the IC50 values for the parent strains, respectively. Two F. proliferatum strains initially demonstrated high resistance to fludioxonil and produced FUM 6 and 22 times higher than the average for the other parent strains of this species.
Текст научной статьи Физиолого-биохимические свойства лабораторных мутантов грибов Fusarium, резистентных к флудиоксонилу
Фузариоз зерновых культур — одно из наиболее вредоносных и активно изучаемых грибных заболеваний, которое приводит к значительным потерям урожая зерна и снижению его качества. Среди грибов Fusarium , вызывающих это заболевание, многие виды образуют токсичные вторичные метаболиты (микотоксины), которые имеют выраженный видоспецифичный характер (1-3). К широко распространенным и агрессивным патогенам зерновых культур относится вид F. graminearum Schwabe, образующий микотоксины дезоксиниваленол (ДОН) и зеараленон (ЗЕН) (4-6). Другой вид F. proliferatum (Matsush.) Nirenberg ex Gerlach & Nirenberg, с недавнего времени также часто выявляемый в микобиоте зерна, способен продуцировать фумонизины группы В, представленные преимущественно фумонизином В 1 (ФУМ) (7-9).
Обработку фунгицидами считают наиболее эффективной и распространенной стратегией борьбы с фузариозом, однако известно, что при применении этого элемента технологии в популяциях грибов Fusarium могут появиться изоляты, резистентные к препаратам с действующими веществами из разных классов — бензимидазолов (10), стробилуринов (11), триазолов (12), а также к флудиоксонилу (ФДО) — несистемному фунгициду из класса фенилпирролов (13). Штаммы Fusarium , устойчивые к ФДО, впервые выявлены в популяциях F. oxysporum и F. sambucinum , выделенных из клубней картофеля (14), а позже обнаружены в популяциях F. graminearum (15-17) и F. pseudograminearum (18).
Свойства резистентных к ФДО штаммов Fusarium (скорость роста, чувствительность к осмотическому стрессу, интенсивность спороношения, патогенность и токсинообразование) могут существенно отличаться от свойств чувствительных штаммов того же вида в популяции, что было продемонстрировано ранее на примере F. graminearum (15, 19, 20), тогда как для гриба F. proliferatum такие исследования не проводились.
В представленной работе впервые определена чувствительность генетически охарактеризованных штаммов двух видов — F. graminearum и F. proliferatum , выделенных из зерна, к ФДО и получены лабораторные мутанты грибов, резистентные к этому действующему веществу (д.в.). Также впервые установлены достоверные отличия лабораторных мутантов F. graminearum и F. proliferatum от исходных штаммов по скорости роста, патогенности для растений пшеницы и токсинопродуцирующей способности.
Целью работы был сравнительный анализ физиолого-биохимических свойств коллекционных штаммов грибов Fusarium graminearum и F. proliferatum и полученных из них в лабораторных условиях мутантов, резистентных к флудиоксонилу.
Методика. Объектами исследования стали 16 моноконидиальных штаммов грибов Fusarium различного происхождения, предварительно идентифицированных по морфологическим признакам как F. graminearum и F. proliferatum, из коллекции лаборатории микологии и фитопатологии (MFG) Всероссийского НИИ защиты растений (ВИЗР, г. Санкт-Петербург, Россия). Для последующих экспериментов штаммы предварительно выращи- вали на картофельно-сахарозной агаризованной среде (КСА) при 25 °C в темноте.
Для молекулярно-генетической идентификации грибов из 10-50 мг мицелия, собранного с поверхностей колоний на КСА, выделяли ДНК по адаптированной методике с помощью 2 % раствора бромида цетилтриметиламмония и хлороформа. Фрагмент гена фактора элонгации трансляции EF-1a ( tef) амплифицировали с использованием праймеров EF1 (5 ‘ -ATG-GGTAAGGARGACAAGAC-3 ’ ) и EF2 (5 ‘ -GGARGTACCAGTSATCATGTT-3 ‘ ) (21). Реакцию проводили в объеме смеси 25 мкл, содержащей 20,5 мкл dH 2 O, 2,5 мкл 10х Taq ПЦР-буфер («Qiagen N.V.», Нидерланды), 0,5 мкл dNTP 10 мМ (ЗАО «Евроген», Россия), по 0,3 мкл каждого праймера в концентрации 25 мкМ (ООО «Компания Алкор Био», Россия), 0,1 ед. акт. TaqAB полимеразы (ООО «Компания Алкор Био», Россия), 1 мкл ДНК. Амплификацию осуществляли по следующему протоколу: 3 мин при 95 ° C; 15 с при 95 ° C, 15 с при 62 ° C, 1 мин при 72 ° C (35 циклов); 3 мин при 72 ° C (C1000 Touch Thermal Cycler, «Bio-Rad», CША).
Нуклеотидные последовательности амплифицированных фрагментов ДНК определяли на секвенаторе ABIPrism 3500 («Applied Biosystems, Hitachi», Япония) с использованием набора реактивов BigDye Terminator v3.1 Cycle Sequencing Kit («Applied Biosystems», CША). Консенсусные нуклеотидные последовательности выравнивали в программе Vector NTI Advance 10 («Thermo Fisher Scientific», CША) и депонировали в базу данных NCBI GenBank. Видовую принадлежность штаммов подтверждали посредством поиска наибольшего сходства среди гомологичных последовательностей репрезентативных штаммов грибов Fusarium из коллекции Northern Regional Research Laboratory (NRRL, CША) и коллекции культур CBS-KNAW (CBS, Нидерланды), представленных в базе данных NCBI GenBank.
В исследовании использовали фунгицид Максим, КC («Syngenta AG», Швейцария), содержащий 25 г/л ФДО, который разводили в стерильной воде до концентрации 5 г/л д.в., соответствующей концентрации рабочего раствора для протравливания семян с максимальной рекомендованной нормой расхода. Рабочий раствор далее последовательно 10-кратно разводили стерильной водой и вносили в KCA, охлажденный до 50 ° C. После тщательного перемешивания питательную среду разливали по 20 мл в чашки Петри диаметром 85 мм.
Для получения мутантов исходные штаммы Fusarium культивировали на КCА, содержащем 0,5 мг/л ФДО, через 7 сут отмечали наличие роста и переносили диск диаметром 4 мм, вырезанный стерильным пробочным сверлом из активно растущей части колонии, на КCА с 5 мг/л ФДО. Затем выросшую культуру вновь пересевали таким же способом на КCА с 25 мг/л ФДО. Через 7-14 сут инкубации отмечали наличие роста и повторяли пересев в тех же условиях. Лабораторные мутанты, полученные в результате пассажей на среде с возрастающей концентрацией фунгицида, проверяли на стабильность, сохраняли и далее использовали для сравнения свойств с исходными коллекционными штаммами.
При определении чувствительности грибов к ФДО его концентрации в питательной среде для исходных штаммов составляли 5; 0,5; 0,05 и 0,005 мг/л, для мутантов — 250; 50; 5 и 0,5 мг/л. Диски с культурами грибов помещали мицелием вниз на поверхность питательной среды в центр каждой чашки. В контрольном варианте диск помещали на поверхность КCА без добавления фунгицида. Через 3 и 5 сут инкубации соответственно штаммов F. graminearum и F. proliferatum определяли средний диаметр колонии гриба, вычитая диаметр инокуляционного диска. Действие фунгицида на линейный рост штамма оценивали по уменьшению диаметра колонии в каждом варианте по сравнению с контролем, выраженному в процентах. Концентрацию фунгицида, приводящую к 50 % подавлению роста (полу-максимальное ингибирование; IC50), рассчитывали с помощью программы Quest Graph™ IC50 Calculator («AAT Bioquest», США).
Для оценки влияния температуры на скорость роста из колоний исходных штаммов и мутантов, выращенных на КСА, вырезали диски диаметром 4 мм, которые помещали на поверхность КСА в центр пластиковой чашки Петри диаметром 85 мм. Культивировали при 25 ° С и 35 ° С в темноте в термостате Innova 44R («Eppendorf», Германия), измеряя диаметр колоний в двух взаимно перпендикулярных направлениях и вычитая диаметр иноку-ляционного диска: для F. graminearum соответственно на 3-и и 7-е сут, для F. proliferatum — на 5-е и 7-е сут. Скорость роста определяли как отношение диаметра колонии к числу суток культивирования (мм/сут).
Патогенность штаммов оценивали при инокуляции отрезков листьев пшеницы ( Triticum aestivum L.) сорта Васса, которую выращивали в вегетационных сосудах с почвой в течение 10 сут при искусственном освещении. Отрезки листьев длиной 5-7 см помещали в кюветы на фильтровальную бумагу, увлажненную 0,004 % водным раствором бензимидазола (22). В каждом варианте опыта использовали по 10 отрезков листьев. В центре каждого отрезка листа делали укол стерильной иглой и помещали диск диаметром 5 мм с культурой каждого штамма, располагая его мицелием вниз. В контрольном варианте на отрезки листьев раскладывали диски, вырезанные из среды КСА без культуры гриба. Кюветы накрывали стеклом, выдерживали при 20 ° С, влажности 60 % и переменном освещении в режиме свет/темнота 16 ч/8 ч в климатической камере MLR-352H («Panasonic», Япония). Через 5 сут измеряли длину образовавшегося некроза (мм) для каждого отрезка листа. Штамм считали непатогенным, если средняя длина некроза был меньше диаметра инокуляционного диска.
Для оценки токсинообразования штаммов в колбы, содержащие по 20 г зерна пшеницы сорта Васса, добавляли по 12 мл воды, стерилизовали при 1 атм. в течение 60 мин в автоклаве (MLS-3751L, «PHCBI», Япония) и инокулировали остывший субстрат двумя дисками диаметром 5 мм с культурой гриба. В качестве контроля использовали неинокулированный стерилизованный субстрат. Все образцы инкубировали в течение 2 нед при 25 ° С, ежесуточно встряхивая, после чего высушивали при 50 ° С в течение 24 ч и измельчали на лабораторной мельнице («IKA-Werke», Германия). Далее к 1 г муки добавляли 10 мл водного раствора ацетонитрила (84:16, v/v) и выдерживали 14-16 ч на шейкере S-3M («ELMI», Латвия) при 300 об/мин с 2-кратным интенсивным перемешиванием в начале и конце экстракции.
Количество ДОН и ЗЕН в культурах F. graminearum и ФУМ в культурах F. proliferatum определяли с помощью аттестованных коммерческих иммуноферментных тест-систем Дезоксиниваленол-ИФА, Зеараленон-ИФА и Фумонизин В 1 -ИФА (ВНИИВСГЭ, Россия). При 10-кратном разведении экстрактов фосфатно-солевым буферным раствором (рН 7,4) с Tween 20 предел определения ДОН, ЗЕН и ФУМ был равен 0,1 мкг/г.
Для расчета средних значений ( M) показателей и их доверительных интервалов при уровне значимости 95 % ( 1 0,05 х SEM) использовали программы Microsoft Excel 2010 и Statistica 10.0 («StatSoft, Inc.», США). В зависимости от типа распределения сравнение средних значений выборок проводили с помощью параметрического критерия Тьюки или непараметрического критерия Краскела-Уоллиса. Различия считали достоверными при p < 0,05.
Резуёьтаты . Штаммы грибов Fusarium, использованные в работе, приведены в таблице 1.
1. Штаммы грибов Fusarium , включенные в исследование
|
Штамм |
Вид |
Происхождение, год |
Растение-хозяин |
Номер последователь ности tef в GenBank |
|
MFG 58922 |
F. graminearum |
Краснодарский край, 2016 |
Ячмень |
PP449338* |
|
MFG 58954 |
F. graminearum |
Курская область, 2016 |
Пшеница |
PP449339* |
|
MFG 59151 |
F. graminearum |
Краснодарский край, 2016 |
Рис |
PP449340* |
|
MFG 59156 |
F. graminearum |
Воронежская область, 2016 |
Пшеница |
PP449341* |
|
MFG 60025 |
F. graminearum |
Алтайский край, 2016 |
Пшеница |
PP449342* |
|
MFG 60327 |
F. graminearum |
Псковская область, 2018 |
Ячмень |
PP449344* |
|
MFG 60782 |
F. graminearum |
Амурская область, 2019 |
Ячмень |
MW273166 |
|
MFG 60789 |
F. graminearum |
Амурская область, 2019 |
Пшеница |
MW273174 |
|
MFG 58471 |
F. proliferatum |
Краснодарский край, 2012 |
Пшеница |
MW811115* |
|
MFG 58589 |
F. proliferatum |
Ленинградская область, 2013 |
Овес |
MW811118* |
|
MFG 58590 |
F. proliferatum |
Приморский край, 2012 |
Овес |
MW811119* |
|
MFG 58603 |
F. proliferatum |
Липецкая область, 2014 |
Кукуруза |
MW811120* |
|
MFG 58666 |
F. proliferatum |
Нижегородская область, 2014 |
Овес |
PP449337* |
|
MFG 60308 |
F. proliferatum |
Воронежская область, 2017 |
Ячмень |
PP449343* |
|
MFG 60309 |
F. proliferatum |
Краснодарский край, 2018 |
Пшеница |
MW811125* |
|
MFG 60412 |
F. proliferatum |
Краснодарский край, 2012 |
Пшеница |
PP449345* |
Примечание. Звездочками обозначены последовательности, полученные в настоящем исследовании.
Предварительную идентификацию штаммов подтвердили при анализе последовательности фрагмента гена tef , наиболее информативного для дифференциации видов Fusarium (23). Установлено наибольшее сходство (99,51-99,85 %) последовательностей tef анализируемых штаммов F. graminearum с гомологичной последовательностью репрезентативного штамма F. gra-minearum NRRL 31084 (GenBank acc. MW233103). Последовательности tef анализируемых штаммов F. proliferatum сравнивали с гомологичными последовательностями трех штаммов этого вида — NRRL 31071 (AF291058), NRRL 32155 (FJ538242), CBS 131570 (JX118976), сходство составило 98,99100 %.
При культивировании на КСА в контроле без фунгицида диаметр колоний у исходных штаммов F. graminearum варьировал от 34 до 52 мм на 3-и сут культивирования, у исходных штаммов F. proliferatum — от 48 до 64 мм на 5-е сут (рис. 1).
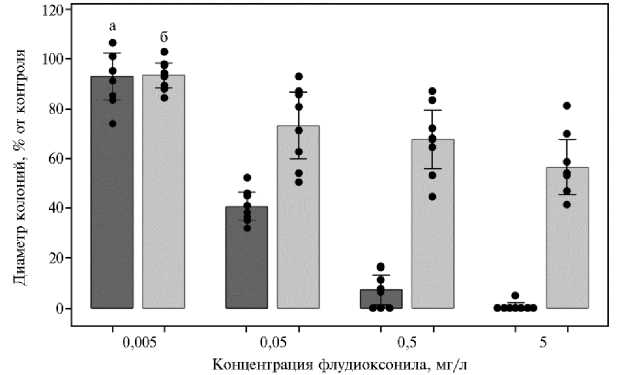
Рис. 1. Рост исходных штаммов Fusarium graminearum (а) на 3-и сут культивирования и F. proliferatum (б) на 5-е сут культивирования под влиянием флудиоксонила в различных концентрациях. Точками указаны значения для индивидуальных штаммов, отрезками — доверительные интервалы средних значений для выборки штаммов одного вида при уровне значимости 95 %, M± ( t 0,05 * SEM).
При минимальной концентрации фунгицида в среде 0,005 мг/л по- давление роста пяти штаммов F. graminearum в сравнении с контролем составляло 4-26 % (р < 0,02), а у двух штаммов MFG 58922 и 60789 наблюдали стимулирование роста на 7 % (р < 0,003). При этой же концентрации ФДО ограничение роста штаммов MFG 58590 и 58603 F. proliferatum составило 6 и 16 % в сравнении с контролем (р < 0,01), а у шести штаммов не имело статистически значимых отличий от контроля. При концентрации ФДО 5 мг/л только один штамм F. graminearum MFG 60782 образовал колонию небольшого диаметра (2,5±0,3 мм), тогда как рост остальных штаммов этого вида был полностью подавлен. В тех же условиях все штаммы F. proliferatum были способны к росту, подавление которого составило 19-58 % (р < 0,01). Два штамма F. proliferatum MFG 58471 и MFG 58590 продемонстрировали низкую чувствительность к ФДО — подавление их роста при наибольшей концентрации вещества не превышало 30 % (р < 0,01). Очевидно, эти штаммы изначально обладали резистентностью к ФДО, поэтому дальнейшего индуцирования их резистентности к этому действующему веществу не проводили.
При поэтапном культивировании исходных штаммов F. graminearum и F. proliferatum на среде с возрастающей концентраций ФДО были получены лабораторные мутанты, которые на 7-е сут культивирования на питательной среде КСА, содержащей 25 мг/л ФДО, формировали колонии диаметром более 50 мм. У мутантов отмечали нетипичное очертание колоний с появлением более активно растущих секторов из отдельных гиф. После субкультивирования в течение ряда пассажей и хранения при 4 ° C на КСА без добавок на протяжении нескольких месяцев у мутантов сохранялся исходный уровень резистентности.
При культивировании на КСА диаметр колоний мутантов F. graminearum варьировал от 39 до 63 мм на 3-и сут, а у мутантов F. proliferatum составил 54-68 мм на 5-е сут (рис. 2).
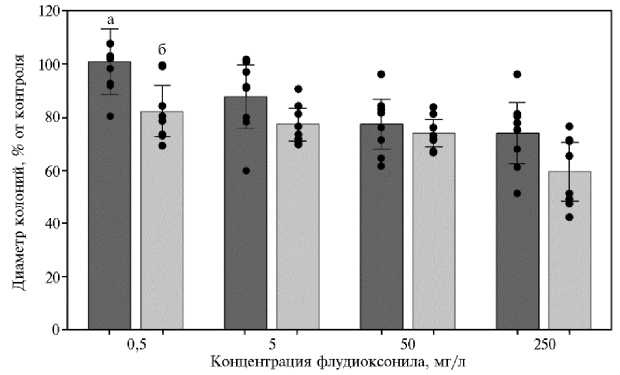
Рис. 2. Рост лабораторных мутантов Fusarium graminearum (а) на 3-и сут культивирования и F. proliferatum (б) на 5-е сут культивирования под влиянием флудиоксонила в различных концентрациях. Точками указаны значения для индивидуальных штаммов, отрезками — доверительные интервалы средних значений для выборки штаммов одного вида при уровне значимости 95 %, M± ( 1 0,05 х SEM).
При концентрации ФДО 5 мг/л рост шести исходных штаммов F. graminearum был подавлен на 3-40 % (р < 0,03) в сравнении с контролем, а два штамма оказались толерантными к такой концентрации действующего вещества. При той же концентрации действующего вещества ингибирование роста мутантов F. proliferatum происходило на 9-30 % (р < 0,009). При максимальной концентрации действующего вещества в среде (250 мг/л) подавление роста мутантов у F. graminearum составило 4-49 % (р < 0,001), у F. proliferatum - 24-58 % (р < 0,0002).
Концентрация ФДО, приводящая к полумаксимальному ингибированию роста, для исходных штаммов F. graminearum варьировала от 1,6 до 5,5 мг/л, для их мутантов — от 4000 до 199000 мг/л (табл. 2).
2. Концентрации флудиоксонила, вызывающие полумаксимальное ингибирование роста исходных штаммов и лабораторных мутантов Fusarium
|
Штамм |
Вид |
IC 50 , мг/л |
|
|
исходный штамм |
мутант |
||
|
MFG 58922 |
F. graminearum |
4,7 |
66000 |
|
MFG 58954 |
F. graminearum |
3,3 |
199000 |
|
MFG 59151 |
F. graminearum |
4,7 |
4200 |
|
MFG 59156 |
F. graminearum |
2,8 |
52000 |
|
MFG 60025 |
F. graminearum |
4,1 |
36000 |
|
MFG 60327 |
F. graminearum |
4,0 |
148000 |
|
MFG 60782 |
F. graminearum |
1,6 |
24000 |
|
MFG 60789 |
F. graminearum |
5,5 |
62000 |
|
Среднее, M± ( t 0,05 х |
SEM) |
3,8±0,4 |
74000±23000 |
|
MFG 58471 |
F. proliferatum |
20000 |
- |
|
MFG 58589 |
F. proliferatum |
345 |
156000 |
|
MFG 58590 |
F. proliferatum |
34000 |
- |
|
MFG 58603 |
F. proliferatum |
2100 |
102000 |
|
MFG 58666 |
F. proliferatum |
5700 |
36000 |
|
MFG 60308 |
F. proliferatum |
75 |
34000 |
|
MFG 60309 |
F. proliferatum |
721 |
30000 |
|
MFG 60412 |
F. proliferatum |
650 |
49000 |
|
Среднее, M± ( t 0,05 х |
SEM) |
1600±700а |
57000±16000 |
Примечание. Прочерки означают, что у штаммов MFG 58471 и MFG 58590, исходно резистентных к флудиоксонилу (ФДО), мутантов не получали; а — без учета IC 50 ФДО для штаммов MFG 58471 и MFG
В среднем IC 50 ФДО для исходных штаммов и мутантов F. graminearum достоверно различались в 18500 раз (р = 0,008). У шести исходных штаммов F. proliferatum и полученных из них лабораторных мутантов интервалы варьирования IC 50 ФДО составили соответственно 75-5650 и 30000156000 мг/л. В среднем IC 50 ФДО у мутантов F. proliferatum оказалась в 36 раз выше (р = 0,002), чем среднее значение этого показателя для исходных штаммов и в ~ 2 раза выше, чем для двух штаммов F. proliferatum MFG 58471 и 58590, изначально резистентных к ФДО.
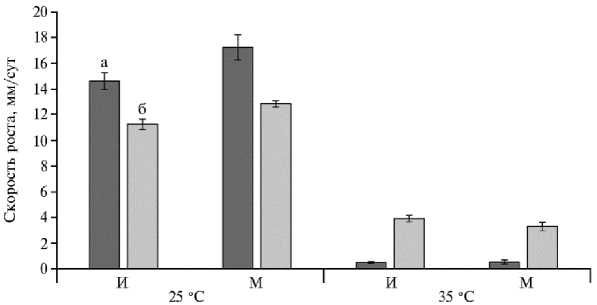
Рис. 3. Скорость роста исходных штаммов (И) и лабораторных мутантов (М) Fusarium graminearum (а) и F. proliferatum (б) при 25 и 35 ° С. Отрезками указаны доверительные интервалы средних значений для выборки штаммов одного вида при уровне значимости 95 %, M± ( t 0,05 х SEM).
При 25 ° C скорость роста исходных штаммов F. graminearum (от 11,4
до 17,3 мм/сут) в среднем была в 1,3 раза выше, чем у F. proliferatum (от 9,6 до 12,9 мм/сут), тогда как при 35 ° C культуры F. proliferatum росли в 7,5 раза активнее, чем F. graminearum (рис. 3).
У шести мутантов F. graminearum при 25 ° C скорость роста увеличилась на 0,6-7,4 мм/сут в сравнении с исходными штаммами, тогда как у MFG 59151 и MFG 60025 оказалась ниже на 0,7 и 3,8 мм/сут. В тех же условиях скорость роста лабораторных мутантов F. proliferatum была больше на 0,3-3,1 мм/сут, чем у исходных штаммов этого вида.
Средние показатели скорости роста мутантов F. graminearum и F. proliferatum (17,2±1,0 и 12,8±0,2 мм/сут) статистически значимо (р = 0,045 и р = 0,02) превышали рассчитанные для исходных штаммов (14,6±0,7 и 11,2±0,4 мм/сут). При 35 ° C достоверных различий по средней скорости роста у исходных штаммов и мутантов F. graminearum и F. proliferatum мы не выявили.
При инокуляции отрезков листьев пшеницы все исходные штаммы F. graminearum были патогенными и вызывали некрозы длиной от 22±2 до 45±1 мм (рис. 4). В среднем агрессивность мутантов F. graminearum оказалась в 1,6 раза ниже (р = 0,03), чем у исходных штаммов. Два мутанта — MFG 60025 и MFG 60782, в отличие от исходных культур, не вызывали появления некрозов, тогда как остальные шесть приводили к их образованию (длина от 14±3 до 44±1 мм). При этом у двух мутантов агрессивность была сопоставима с исходными штаммами, у четырех — снижалась в 1,52,6 раза. Все проанализированные коллекционные штаммы F. proliferatum , а также полученные из них резистентные к ФДО мутанты оказались непатогенными в условиях эксперимента.
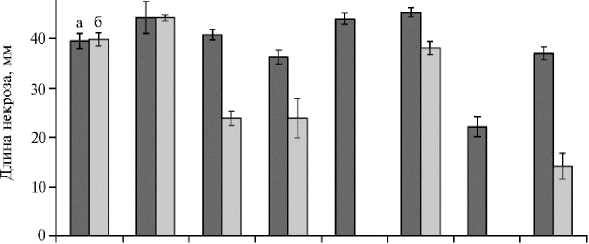
58922 58954 59151 59156 60025 60327 60782 60789
MFG Fusarium graminearum
Рис. 4. Патогенность исходных штаммов (а) и лабораторных мутантов (б) Fusarium graminearum в отношении листьев пшеницы ( Triticum aestivum L.) сорта Васса (5 сут, 20 ° С). Отрезками указаны доверительные интервалы средних значений для выборки штаммов одного вида при уровне значимости 95 %, M± ( 1 0,05 х SEM) ( n = 10).
3. Токсинопродуцирующая способность исходных штаммов и лабораторных мутантов Fusarium graminearum (зерно пшеницы сорта Васса, 25 ° C, 14 сут в темноте)
|
Штамм |
Дезоксиниваленол, мкг/г |
Зеараленон, мкг/г |
||
|
исходный штамм |
мутант |
исходный штамм | |
мутант |
|
|
MFG 58922 |
20 |
0,5 |
631 |
52 |
|
MFG 58954 |
0,2 |
0,2 |
36 |
0,3 |
|
MFG 59151 |
52 |
1,6 |
45 |
0,8 |
|
MFG 59156 |
0,8 |
1,0 |
5 |
0,4 |
|
MFG 60025 |
0,3 |
< LOQ |
39 |
1,0 |
|
MFG 60327 |
133 |
0,2 |
82 |
0,4 |
Ïðîäîëæåíèå òàáëèöû 3 MFG 60782 2,6 3,2 40 1,7
MFG 60789 9,6 1,9 75 1,1
Среднее, M± ( ®,05 x SEM) 27±16 1,1±0,4 119±72 7,2±6,3
Примечание. < LOQ означает количество микотоксина ниже предела обнаружения.
Все исходные штаммы F. graminearum продуцировали ДОН и ЗЕН in vitro (табл. 3). Средние количества микотоксинов, выявленные у мутантов F. graminearum , были ниже соответственно в 25 (р = 0,09) и 16 (р = 0,005) раз, чем у исходных штаммов. Резкое снижение способности продуцировать ДОН, вплоть до полного прекращения его синтеза, наблюдалось у большинства (62,5 %) мутантов F. graminearum , тогда как у остальных не было отличий от исходных штаммов. У всех мутантов F. graminearum способность продуцировать ЗЕН была выражена в 12-205 раз меньше, чем у исходных культур.
Все штаммы F. proliferatum продуцировали ФУМ, количества которого также существенно варьировали (табл. 4). Четыре мутанта F. proliferatum образовывали в 2-25 раз больше этого микотоксина, а два — в 2,0-2,2 раза меньше, чем исходные штаммы. В среднем мутанты F. proliferatum продуцировали в 4 раза больше ФУМ (р = 0,02), чем исходные культуры. Штаммы F. proliferatum MFG 58471 и 58590 с изначально высокой резистентностью к ФДО образовывали ФУМ в 6 и 22 раза выше среднего показателя для шести исходных чувствительных штаммов, а также в 1,4 и 5,4 раза — для мутантов, полученных в лабораторных условиях.
4. Токсинопродуцирующая способность исходных штаммов и лабораторных мутантов Fusarium proliferatum (зерно пшеницы сорта Васса, 25 ° С, 14 сут в темноте)
|
Штамм |
Фумонизин В 1 , мкг/г |
|
|
исходный штамм |
мутант |
|
|
MFG 58471 |
1259 |
- |
|
MFG 58589 |
327 |
164 |
|
MFG 58590 |
4842 |
- |
|
MFG 58603 |
531 |
245 |
|
MFG 58666 |
160 |
1622 |
|
MFG 60308 |
81 |
148 |
|
MFG 60309 |
48 |
1216 |
|
MFG 60412 |
164 |
2000 |
|
Среднее, М± ( / 0,05 x SEM) |
219±63 |
900±284a |
Примечание. Прочерки означают, что у штаммов MFG 58471 и MFG 58590, исходно резистентных к флудиоксонилу (ФДО), мутантов не получали; а — без учета штаммов MFG 58471 и MFG 58590.
Выбранные для исследования коллекционные штаммы F. graminearum и F. proliferatum были выделены из образцов различных зерновых культур и идентифицированы с помощью молекулярно-генетических методов. Коллекционные штаммы F. proliferatum оказались в среднем в 421 раз менее чувствительны к ФДО, чем штаммы F. graminearum. Ранее уже сообщалось о межвидовых различиях по чувствительности грибов Fusarium к этому фунгициду (24-26), в том числе было показано, что штаммы F. proliferatum , выделенные из кукурузы, менее чувствительны к ФДО (при концентрации д.в. 0,5 мг/л подавление роста грибов на 3-и сут составило 4-33 %), чем F. graminearum (подавление роста достигало 100 %) (26).
Поэтапное культивирование штаммов двух видов Fusarium на питательной агаризованной среде с последовательным повышением содержания ФДО привело к получению мутантов с устойчивым признаком резистентности. Неоднородный характер роста колоний у мутантов, который выражался в появлении активно растущих секторов воздушного мицелия из гиф, преодолевших прессинг фунгицида, уже отмечали для некоторых штаммов
Fusarium spp. после культивирования на средах с добавлением ФДО, как и факт сохранения этого нового признака при пересевах на среду с тем же фунгицидом (26, 27).
Мутанты F. graminearum достоверно отличались от исходных штаммов по ряду свойств: средняя скорость их роста на КСА без фунгицида при 25 ° С была выше, тогда как агрессивность в отношении листьев пшеницы и количество продуцируемых микотоксинов ДОН и ЗЕН — значительно ниже. Выявленное нами увеличение скорости роста с приобретением резистентности к ФДО у штаммов F. graminearum отличается от описанных ранее наблюдений, в которых резистентные к ФДО мутанты демонстрировали снижение скорости роста по сравнению с чувствительными изолятами (17, 19). Температура 35 ° С оказалась неблагоприятной для роста исходных штаммов и мутантов F. graminearum , который был одинаково слабым без статистически значимых различий по средним показателям. Многие авторы также отмечали у резистентных изолятов F. graminearum снижение способности поражать листья, колосья, проростки пшеницы и плоды томата, а также пониженную способность продуцировать ДОН (15-17). Известно, что ДОН служит фактором патогенности у F. graminearum и способствует инфицированию зерновых культур (28, 29), снижение его продуцирования приводит к потере агрессивности патогена при взаимодействии с растением.
В нашем исследовании впервые получены лабораторные мутанты F. proliferatum , резистентные к ФДО, а также проанализированы их свойства в сравнении с исходными штаммами этого вида и природными мутантами — штаммами MFG 58471 и 58590, изначально обладающими резистентностью к этому действующему веществу. Как и в случае с F. graminearum , мутанты F. proliferatum при 25 ° С росли быстрее, чем исходные штаммы. При повышении температуры до 35 ° С скорость роста у исходных штаммов и мутантов F. proliferatum была одинаковой. Следовательно, при повышении температуры в период вегетации встречаемость F. proliferatum в микобиоте зерновых культур будет возрастать, но в то же время при высокой температуре (30-35 ° С) продуцирование ФУМ снижается (30, 31).
Известно, что количества продуцируемого ФУМ зависят от множества факторов, таких как температура, влажность, освещенность, субстрат. Оптимальная температура для роста гриба не всегда совпадает с температурой, при которой гриб продуцирует наибольшее количество микотоксинов. Ранее выявлено, что максимальная скорость роста трех штаммов F. proliferatum , выделенных из зерна пшеницы, наблюдалась при 25 ° C, а два из них продуцировали максимальные количества ФУМ при более низкой температуре — 15 ° C (32). Предположительно, зерно пшеницы служит менее благоприятным субстратом для продуцирования ФУМ грибами, чем зерно кукурузы (33, 34).
Как исходные штаммы, так и мутанты F. proliferatum не вызывали появления некрозов на отрезках листьев пшеницы в лабораторном тесте, что подтверждает слабую агрессивность этого гриба в отношении зерновых культур (35-37). Важным результатом стало отмеченное значительное увеличение количества синтезируемого ФУМ у большинства лабораторно индуцированных мутантов по сравнению с исходными штаммами, а также значительное продуцирование микотоксина двумя природными резистентными мутантами. Роль ФУМ, в частности фумонизина B1, как фактора патогенности грибов до сих пор остается спорной, что, вероятно, связано с конкретными взаимоотношениями, возникающими между различными растениями-хозяевами и продуцентами этих микотоксинов (9, 38, 39). Многочисленные исследования демонстрируют отсутствие связи между способно- стью грибов Fusarium продуцировать ФУМ и их патогенностью в отношении кукурузы и других культур (40-42). Однако, согласно некоторым публикациям, фумонизин B1 способствует подавлению иммунных реакций растения-хозяина за счет ингибирования синтеза церамидов, что приводит к запрограммированной гибели клеток (29, 43).
Неоднородность окружающей среды приводит к сосуществованию в популяциях грибов штаммов, характеризующихся различной чувствительностью к фунгицидам (44). Наряду с глобальными природными процессами, обусловленными изменением климата, агробиоценоз подвергается всем видам воздействия, которые связаны с деятельностью человека, направленной на получение высоких урожаев. Применение фунгицидов может приводить к сдвигу в базовой чувствительности к их действующему веществу у представителей биоты. Распространение резистентности к фунгицидам в популяциях грибов во многом зависит от изменений их адаптационных свойств и динамики конкуренции между устойчивыми и чувствительными штаммами.
В России научное обеспечение защиты зерновых культур от заболеваний, вызванных патогенными микроорганизмами, успешно развивается и остается в числе приоритетных направлений поиска (45). Гетерогенный характер влияния фунгицидных обработок на токсиногенез F. graminearum, а также способность триазолов стимулировать образование ДОН ранее были установлены в серии лабораторных экспериментов (46, 47). Однако системный комплексный подход к изучению многих важных аспектов формирования резистентности популяций пока не реализован. При планировании будущих проектов, несомненно, следует учитывать все многообразие разнонаправленных динамических процессов, приводящих к изменениям адаптационных свойств фитопатогенов, а также особенностей конкурентных отношений между устойчивыми и чувствительными штаммами.
Таким образом, мы определили базовую чувствительность генетически охарактеризованных штаммов двух видов Fusarium graminearum и F. proliferatum , выделенных из зерновых культур на территории России, к фунгициду, содержащему флудиоксонил. Получены лабораторные мутанты грибов F. graminearum и (впервые в мире) F. proliferatum , резистентные к этому действующему веществу, изучены их свойства в сравнении с исходными коллекционными штаммами. Показано, что концентрации флудиок-сонила IC 50 , приводящие к полумаксимальному ингибированию роста мутантов F. graminearum и F. proliferatum , соответственно в 900-60000 и 6-458 раз выше значений для исходных штаммов. Скорость роста лабораторных мутантов F. graminearum и F. proliferatum при 25 ° С выше на 2,6 и 1,6 мм/сут в сравнении со средними показателями исходных культур. При 35 ° С достоверных различий по средней скорости роста у исходных штаммов и мутантов обоих видов грибов не выявлено. Шесть лабораторных мутантов F. graminearum были патогенными для пшеницы, однако вызываемые ими некрозы на отрезках листьев оказались в 1,6 раза меньше, чем в случае исходных штаммов гриба, тогда как два мутанта F. graminearum утратили патогенность. Все лабораторные мутанты F. proliferatum , как и исходные штаммы, оказались неспособны инфицировать листья пшеницы в условиях эксперимента. Средние количества ДОН и ЗЕН, продуцируемые мутантами F. graminearum , были соответственно в 25 и 16 раз ниже, чем у исходных штаммов гриба. При этом мутанты F. proliferatum в среднем синтезировали в 4 раза больше ФУМ, чем исходные штаммы. Полученные нами результаты показывают, что у разных видов Fusarium приобретение резистентности к одному и тому же химическому препарату неодинаково влияет на метаболизм грибов, что
îòðàæàåòñÿ â èçìåíåíèè èõ ñâîéñòâ.
ËÈÒÅÐÀÒÓÐÀ
1 ФГБНУ Всероссийский НИИ защиты растений,
Поступила в редакцию 13 июля 2024 года Принята к публикации 19 августа 2024 года
Sel’skokhozyaistvennaya biologiya [ Agricultural Biology ], 2025, V. 60, № 1, pp. 138-152
PHYSIOLOGICAL AND BIOCHEMICAL CHARACTERS OF LABORATORY INDUCED Fusarium MUTANTS RESISTANT TO FLUDIOXONIL
A.S. Orina1 8 , O.P. Gavrilova1, T.Yu. Gagkaeva1, E.P. Arabina1, A.A. Burkin2, G.P. Kononenko2
-
1Al l-Russian Institute of Plant Protection , 3, sh. Podbel’skogo, St. Petersburg, 196608 Russia, e-mail orina-alex@yandex.ru ( И corresponding author), olgavrilova1@yandex.ru , t.gagkaeva@mail.ru , arabina2001@gmail.com ;
-
2Al l-Russian Research Institute of Veterinary Sanitation, Hygiene, and Ecology — Branch of FSC ARRIEV RAS, 5, Zvenigorodskoe sh., Moscow, 123022 Russia, e-mail aaburkin@mail.ru , kononenkogp@mail.ru
The authors declare no conflict of interests
Final revision received July 13, 2024
Accepted August 19, 2024
