Флуктуации и сукцессии в лесных сообществах на фоне изменения климата

Автор: Маслов Александр Анатольевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
В заповедных лесных участках Московской обл. на протяжении 25 лет изучены разные формы природной динамики лесных сообществ (сукцессии, флуктуации) и их переходы. Показано, что основными формами динамики на заповедных территориях являются флуктуации, сукцессии в результате изменения климата и поздние фазы вторичных сукцессий после пожаров и рубок. Флуктуации и циклические сукцессии могут переходить во вторичные и вековые сукцессии в результате изменения климата.
Сукцессии, флуктуации, изменение климата, заповедные лесные участки, московская обл
Короткий адрес: https://sciup.org/148200814
IDR: 148200814 | УДК: 630
Fluctuations and successions in forest communities under climate change
Different types of natural forest dynamics on community level (successions, fluctuations and transitional types) were studied in strict forest reserves of the Moscow region during 25 years. It was shown that principal types of dynamics in protected areas are fluctuations, successions resulting from climate change and late secondary successions after forest fires and clear-cuttings. Fluctuations and cyclic successions could turn to secondary successions and secular successions as a result of climate change.
Текст научной статьи Флуктуации и сукцессии в лесных сообществах на фоне изменения климата
К концу XX века в геоботанической литературе сложились достаточно полные представления о различных формах динамики растительных сообществ [11, 9, 1, 15]. Если исключить специфический случай первичных сукцессий на вновь возникших минеральных субстратах и не рассматривать сезонную динамику, то большинство авторов выделяет четыре формы динамики. 1) Флуктуации – непродолжительные и ненаправленные малые сдвиги состава или структуры популяций в ответ на небольшие нарушения. Продолжительность «возврата» сообщества в исходное состояние не должна превышать 10 лет [11]. 2) Циклические сукцессии (демутации) – восстановление компонентов растительности при их нарушении на площади, сопоставимой с площадью сообщества. По времени такие сукцессии превышают 10 лет. 3) Вторичные сукцессии – восстановление зрелого сообщества после масштабного нарушения (пожары, ураганы, рубки, вспышки дендрофагов). 4) Вековые сукцессии (гологенез) протекают в пределах ландшафта как результат длительных изменений п риродной среды или климата. Типичным примером такой сукцессии является рост верхового болота.
Гораздо менее полными являются представления о переходе одних форм динамики растительных сообществ в другие, а также о том, как разные формы динамики друг с другом взаимодействуют при изменении внешних условий (климата). Особую ценность для анализа разных форм природной динамики представляют данные прямых и длительных наблюдений на постоянных пробных площадях (ППП) в условиях заповедного режима. Рассмотрим конкретные формы динамики и их переходы друг в друга на примере природных процессов в заповедных лесных участках Московской обл. за 25 лет наблюдений.
МАТЕРИАЛЫ И МЕТОДЫ
На протяжении трех десятилетий в различных типах заповедных условно-коренных лесов Московской области ведется работа по мониторингу
природной динамики [6]. Разработанная база данных по динамике древостоев, подроста, подлеска и нижних ярусов обновляется с частотой 1–5 лет. Достоверность выявления флуктуаций и сукцессионных трендов обеспечивается большими размерами выборки на площадках 0.2×0.2 м.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Типичный ход автогенных вторичных сукцессий описан нами в хвойных лесах бореального типа из класса Vaccinio-Piceetea . В сосняках этого класса – зеленомошнике, черничнике, долгомошнике сосна повсеместно представлена популяциями регрессивного типа, подрост отсутствует, тогда как ель имеет инвазионный тип популяций, переходящий в нормальный тип [7]. В послепожарном столетнем сосняке-зеленомошнике наряду с внедрением ели наблюдается замещение раннесукцессионных видов ( Cladina spp., Festuca ovina ) позднесукцессионными видами – такими как Vaccinium myr-tillus , Hylocomium splendens и Sphagnum girgensoh-nii [5]. На рис. 1 показана динамика популяции Vaccinium myrtillus , за 25 лет ставшей содоминан-том травяно-кустарничкового яруса наряду с Vac-cinium vitis-idaea .
Флуктуации и циклические сукцессии (демутации) в литературе описаны как обратимые сдвиги в составе (или структуре) сообществ в ответ на слабые (флуктуации) или сильные нарушения (циклические сукцессии). Априори предполагается, что возврат в исходное или близкое состояние происходит на фоне стабильного климата. Возникает вопрос – настолько стабильным является климат на протяжении нескольких циклов флуктуаций (масштаб времени – десятилетия) или на протяжении одного цикла демутации (масштаб времени для лесных сообществ порядка 50–100 лет)?
С начала метеорологических наблюдений в конце XIX века и до второй половины XX века характерной особенностью климата центра Русской равнины, как отмечалось в литературе, являлись периодически повторяющиеся (примерно раз в 10 лет) экстремальные зимы и засухи [14]. В результате экстремальных зим (с морозами за –40°) происходило масштабное повреждение и даже гибель
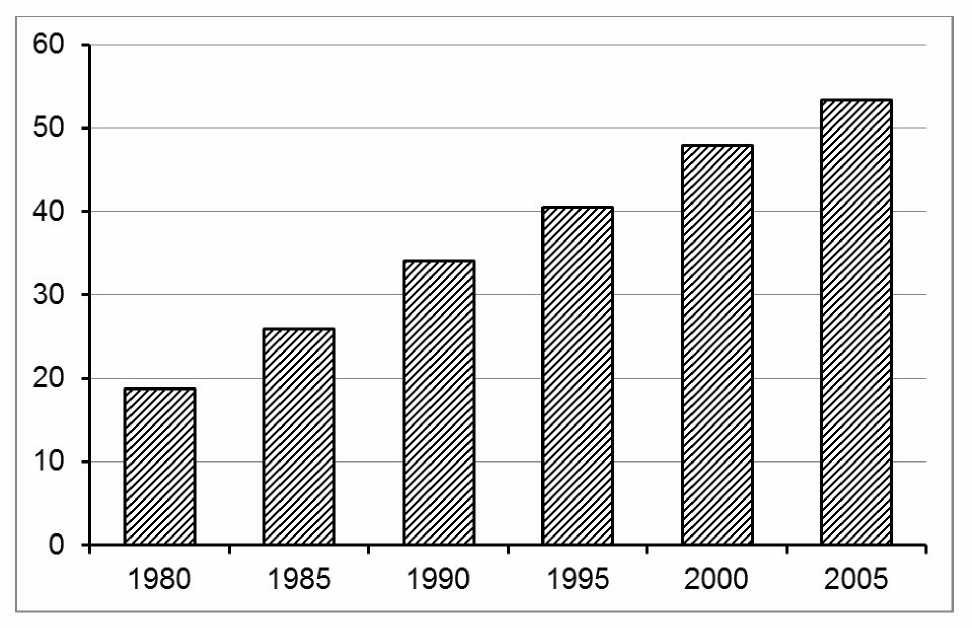
Рис. 1 . Динамика корневой встречаемости (%) черники ( Vaccinium myrtillus ) в сосняке-зеленомошнике (количество учетных площадок N=2000).
неморальных древесных пород: ясеня, вяза, клена, серьезное повреждение дуба, а также гибель надземных побегов лещины.
Повторяющиеся примерно раз в 10 лет экстремальные зимы создавали в смешанных неморальных лесах из класса Querco-Fagetea классический цикл флуктуаций численности лещины – доминирующей породы подлеска. Семилетний ход такой флуктуации – с момента гибели стволов лещины в зиму 1978/79 г. и до смыкания вновь возникшего полога был описан в работе [13]. Полный цикл флуктуации, по мнению авторов, составляет 10 лет. По иронии судьбы, данный цикл оказался последним в ряду флуктуаций.
Во второй половине XX века экстремальные зимы в центре Русской равнины случались все реже и реже. Зима 1978/79 г. «с морозом за –40°» была последней в ряду наблюдений. За последующие 30 лет ни одна из холодных (неэкстремальных) зим не приводила к гибели или серьезному повреждению дуба, ясеня, вяза, лещины. В европейской части России наблюдается потепление климата, особенно сильное с конца 1980-х годов [3]. Одновременно с существенным увеличением безморозного периода (на 39 дней с 1948 по 1998 г.) наблюдается увеличение общего количества осадков.
В связи с прекращением периодически наблюдавшихся экстремальных зим, потеплением, увеличением безморозного периода в хвойно-широколиственных неморальных лесах с конца XX века наблюдается сильная перестройка. Широколиственные породы и неморальные виды нижних яру- сов получили новое конкурентное преимущество, их роль и значение за 30 лет увеличились. Восстановление яруса из лещины из флуктуации перешло в сукцессию. Однако тип, продолжительность такой сукцессии в настоящее время оценить затруднительно. Новая экстремальная зима в любой момент может прервать процесс.
На рис. 2 представлен ход динамики популяции лещины в смешанном ельнике с осиной немораль-но-кисличном (ППП 65–1). К моменту последней экстремальной зимы 1978/79 г. лещина была представлена редко стоящими кустами высотой до 3–5 м. В 1980 г. (момент начала регулярных учетов на ППП) все старые стволы лещины стояли мертвыми, но появились молодые прикорневые побеги высотой до 1 м. За 10 лет лещина восстановила первоначальную численность и высоту. Флуктуация завершилась, но процесс перешел в сукцессию. Через 20 лет число стволов лещины удвоилось, высота отдельных кустов превысила 6 м (рис. 2). При этом лещина сформировала сомкнутый ярус по всей территории ельника, существенно изменив освещенность на уровне нижних ярусов.
Наиболее сильные изменения видового состава и структуры фитоценозов обнаружены нами в группе субнеморальных кисличных ельников на стыке классов Vaccinio-Piceetea и Querco-Fagetea [8], где изначально в составе сообществ были представлены группы бореальных, субнеморальных и неморальных видов, наблюдалось максимальное видовое разнообразие и наименьшая концентрация доминирования [4].
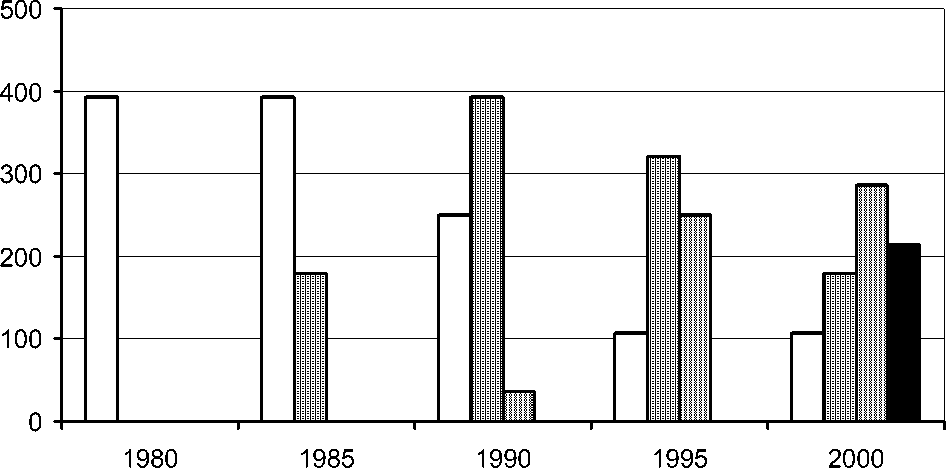
□0.50–1.50 ■ 1.51–3.00 п 3.01–4.50 ■ >4.50
Рис. 2 . Динамика распределения кустов лещины ( Corylus avellana ) по высоте в ельнике неморально-кисличном (ППП 65–1) после последней экстремальной зимы 1978/79 г., экз./га.
Одновременно с формированием яруса из лещины в кисличных типах леса появляется подрост клена. В нижних ярусах кисличных типов леса уменьшается роль видов олиготрофной группы черники-брусники, но возрастает роль видов эвтрофной группы копытня и папоротников. Так, Galeobdolon luteum за 25 лет наблюдений из единично представленного вида превратился в доминирующий вид ельника-кисличника, достигнув встречаемости 50% [7]. Увеличивают свое участие в кисличных ельниках и такие неморальные виды как цирцея ( Circaea alpina ) и медуница ( Pulmonaria obscura ). В результате сукцессии возрастает богатство почвы доступным азотом, что было показано с помощью индикации по шкалам Элленберга.
В то же время более краткосрочные и типичные флуктуации – такие как зарастание пороев кабанов и краткосрочное разрастание нитрофильных видов в местах падения крон живых деревьев за последние десятилетия не претерпели существенных изменений и завершаются восстановлением исходной растительности за 5–10 лет.
В связи с отсутствием экстремальных зим, общим потеплением климата и продолжающимися экстремальными засухами (особенно сильными – 2002 и 2010 гг.) в кисличных и неморальнокисличных ельниках Центра Русской равнины с начала XXI века наблюдаются и другие сукцессии – на стыке циклических и вековых климатогенных. Это – масштабные усыхания ельников в результате длительных вспышек короеда типографа [10]. Как показали наши исследования, отмирание материнского полога в заповедных кисличных и немораль-но-кисличных ельниках приводит к формированию яруса древостоя / подлеска из рябины и/или лещины при незначительном участии ели в возобновлении [2]. Описанный классиками «еловый лес как трехстадийная система» [12] на обширных площадях уступает место неморальным сообществам совершенно другого типа, меняется также традиционный ход вторичных сукцессий.
ВЫВОДЫ
Таким образом, в условиях Центра Русской равнины в природных лесах на заповедных территориях продолжаются флуктуации, автогенные и климатогенные сукцессии под влиянием изменения климата. Автогенные сукцессии характерны для бореальных лесов из класса Vaccinio-Piceetea . В кисличных и неморально-кисличных ельниках (на стыке классов Vaccinio-Piceetea и Querco-Fagetea ) наблюдается рост участия неморальных пород кустарников и деревьев (лещина, клен) и неморальных видов нижних ярусов. Прекращение экстремальных зим, увеличение безморозного периода со второй половины XX века существенно изменило ход природных сукцессий и флуктуаций. Часть флуктуаций переходит в сукцессии. Потенциально это – процесс обратимый.
Работа частично поддержана грантом Президента РФ для государственной поддержки ведущих научных школ РФ НШ-2807.2012.4.
Список литературы Флуктуации и сукцессии в лесных сообществах на фоне изменения климата
- Василевич В.И. Некоторые новые направления в изучении динамики растительности//Ботан. ж., 1993. Т. 78, № 10. C. 1-15.
- Ермаков, А.Л. Начальный этап возобновления деревьев в очаге усыхания ели после вспышки короеда типографа/А.Л. Ермаков, А.А. Маслов//Труды Звенигородской биологической станции. 2011. Т. 5. С. 149-150.
- Груза Г.В. Изменение климатических условий европейской части России во второй половине XX века/Г.В. Груза, Э.Я. Ранькова//Влияние изменения климата на экосистемы. М., 2001. С. I.9-I.16.
- Маслов А.А. 1998. Динамика фитоцено-экологических групп видов и типов леса в ходе природных сукцессий заповедных лесов центра Русской равнины//Бюл. МОИП. Отд. биол. Т. 103. № 2. С. 34-43.
- Маслов А.А. Сукцессионная динамика древостоя и нижних ярусов в послепожарном 100-летнем сосняке лишайниково-зеленомошном//Лесоведение. 2002. № 2. С. 23-29.
- Маслов А.А. Мониторинг эталонных лесных экосистем в заповедных лесных участках//Мониторинг природного наследия. М.: Товарищество научных изданий КМК. 2009а. C. 21-37.
- Маслов, А.А. Мониторинг биоразнообразия и процессов природной динамики в заповедных лесных участках: программа и итоги работ за 25 лет//Структура и функции лесов Европейской России. М.: Товарищество научных изданий КМК. 2009б. C. 172-190.
- Маслов А.А. Проблемы классификации заповедных ельников Московского региона с применением флористических критериев//Растительность Восточной Европы: классификация, экология и охрана. Брянск: Ладомир, 2009в. C. 143-146.
- Миркин Б.М. Антропогенная динамика растительности//Итоги науки и техники. Ботаника. М.: ВИНИТИ, 1984. Т. 5. C. 139-235.
- Мозолевская, Е.Г. Особенности развития вспышки массового размножения короеда типографа в ближнем Подмосковье/Е.Г. Мозолевская, В.А. Липаткин//Лесное хозяйство. 2003. № 1. С. 31-33.
- Работнов Т.А. Фитоценология. 2-е изд. М.: Изд-во МГУ, 1983. 296 с.
- Работнов Т.А. Еловый лес как трехстадийная сукцессионная система//Бюл. МОИП. Отд. биол., 1994. Т. 99, № 2. C. 53-60.
- Речан С.П. Флуктуационная динамика лещиново-снытевой дубравы Подмосковья/С.П. Речан, Т.В. Малышева//Бюл. МОИП. Отд. биол., 1992. Т. 97, № 2. C. 90-96.
- Сазонов Б.И. Суровые зимы и засухи. Л.: Гидрометеоиздат, 1991. 240 с.
- Glenn-Lewin D.C., van der Maarel E. Pattern and process of vegetation dynamics/D.C. Glenn-Lewin, E. van der Maarel//Plant succession: theory and prediction. L., etc: Chapman & Hall, 1992. P. 11-59. (Population and community biology series; V. 11).
