Флуоресцентно-гистохимическая характеристика тучных клеток матки крыс в процессе послеродовой инволюции

Автор: Диндяев Сергей Валерьевич, Касаткин Денис Витальевич
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 1 т.30, 2022 года.
Бесплатный доступ
Матка является в своем роде уникальным органом вследствие постоянного ремоделирования в течение репродуктивного периода тканей, ее составляющих. Нарушения инволюционных процессов, происходящих в этом органе после родов, могут стать причиной развития патологических состояний матки. Актуальность исследования определяется активным участием биогенных аминов в регуляции тканевого гомеостаза и гистогенеза матки. Целью работы явилось изучение закономерностей пространственной кооперации тучных клеток матки крыс и содержания в них катехоламинов, серотонина и гистамина в течение послеродового периода. С помощью флуоресцентно-гистохимических методов исследованы тучные клетки матки крыс 100 беспородных самок крыс репродуктивного возраста, разделенных на 11 групп по срокам послеродового периода. Работа выполнена на криостатных срезах препаратов тела матки. Оценивалась плотность пространственного распределения тучных клеток эндометрия и миометрия. Цитоспектрофлуориметрически в условных единицах регистрирующего прибора в клетках определялось содержание гистамина, серотонина и катехоламинов. В результате исследования установлено, что плотность пространственного распределения флуоресцирующих тучных клеток слизистой и мышечной оболочек тела матки минимальна в первые сутки после родов. Наибольшее количество тучных клеток наблюдается на 10-е сутки после родов. Содержание гистамина, серотонина и катехоламинов в тучных клетках миометрия тела матки максимально в первые сутки после родов и постепенно снижаясь, достигает наименьших значений на 10-15-е сутки послеродовой инволюции. Тучные клетки эндометрия характеризуются более высокой, по сравнению с миометрием, насыщенностью серотонином и катехоламинов. В тканевых базофилах миометрия отмечается более высокий уровень гистамина. Установлены корреляционные связи изменений содержания биоаминов. Плотность пространственного распределения тучных клеток эндометрия и миометрия и содержание в них нейромедиаторных биоаминов в течение послеродовой инволюции матки изменяется однонаправленно, несмотря на региональные различия. Возможно, что тучные клетки способствуют восстановлению симпатической иннервации матки в послеродовый период.
Матка крыс, послеродовый период, тучные клетки, гистамин, серотонин, катехоламины
Короткий адрес: https://sciup.org/143178519
IDR: 143178519
Fluorescence-histochemical characteristics of rat's uterine mast cells during its postpartum involution
The uterus is a unique organ of its kind due to the constant remodeling during the reproductive period of the tissues that make it up. Violations of the involution processes occurring in this organ after part can cause the development of pathological conditions of the uterus. The relevance of the study is determined by the active participation of biogenic amines in the regulation of tissue homeostasis and uterine histogenesis. The aim of the work was to study the patterns of spatial cooperation of mast cells in the uterus of rats and the content of catecholamine, serotonin and histamine in them during the postpartum period. Using fluorescent-histochemical methods, mast cells of the uterus of rats were studied in 100 outbred female rats of reproductive age, divided into 11 groups according to the postpartum period. The study was performed on cryostat sections of preparations of the uterine body. The density of the spatial distribution of endometrial and myometrium mast cells was assessed. The content of histamine, serotonin and catecholamine was determined by cytospectrofluorimetry in conventional units of the recording device in the cells. As a result of the study, it was found that the density of the spatial distribution of fluorescent mast cells in the mucosa and muscular layers of the uterine body is minimal on the first day after childbirth. The largest number of mast cells is observed on the 10th day after birth. The content of histamine, serotonin and catecholamine in the mast cells of the myometrium of the uterine body is maximum on the first day after birth and gradually decreasing, reaching the lowest values on the 10-15th day of postpartum involution. Mast cells of the endometrium are characterized by a higher saturation of serotonin and catecholamine compared to the myometrium. In tissue basophils of the myometrium, a higher level of histamine is noted. Correlations of changes in the content of bio amines have been established. The density of the spatial distribution of mast cells of the endometrium and myometrium and the content of neurotransmitter bioamines in them during the postpartum involution of the uterus changes unidirectional, despite regional differences. It is possible that mast cells contribute to the restoration of sympathetic innervation of the uterus in the postpartum period.
Текст научной статьи Флуоресцентно-гистохимическая характеристика тучных клеток матки крыс в процессе послеродовой инволюции
Введение. В области фундаментальной медицины среди современных исследований одно из ведущих мест занимают работы, посвященные изучению за- кономерностей процессов гистогенеза, регенерации, структурных основ жизнедеятельности органов и тканей [1-2]. Матка является в своем роде уникальным органом вследствие постоянного ремоделирования в течение репродуктивного периода тканей, ее составляющих. В настоящее время актуальной задачей является профилактика послеродовых осложнений. Нарушения инволюционных процессов, происходящих в этом органе после родов, могут стать причиной развития патологических состояний матки [3–6]. Нейромедиаторные биогенные амины принимают активное участие в регуляции тканевого гомеостаза и гистогенеза [7–9], в том числе в процессе инволюции матки [10–12]. Большое значение в обмене биоаминов отводится тучным клеткам (далее – ТК) [78, 13]. В настоящее время изучена роль тканевых базофилов в обеспечении матки крыс биоаминами в течение полового цикла [7]. Практически отсутствуют данные об их локализации, пространственной кооперации и динамики насыщенности биоаминами в течение послеродового периода.
Цель исследования: выявить закономерности пространственной кооперации тучных клеток тела матки крыс и установить содержание в них катехоламинов, серотонина и гистамина в течение послеродового периода.
Материалы и методы исследования. Работа выполнена в осенне-зимний период на 55 беспородных самках крыс репродуктивного возраста стандартной массы, распределенных по срокам послеродового периода – 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15 сутки. На выполнение исследования получено заключение локального этического комитета Ивановской государственной медицинской академии Минздрава России от 09.12.2015 г. (протокол № 6). Материалом исследования явились нефиксированные криостатные срезы тела матки. Животных выводили из опыта путем дислокации шейных позвонков согласно «Правил проведения работ с использованием экспериментальных животных» (приказ Минздрава СССР от 13.11.1984 г. № 724).
Для выявления катехоламинов и серотонина в ТК в нефиксированных крио-статных срезах тела матки использован па-раформальдегидный метод Фалька-Хилларпа в модификации Е.М. Крохиной [14]. Образующийся серотониново-параформальдегидный флуорофор имеет максимумы возбуждения 370–410 нм и эмиссии 520–530 нм. Катехоламины флуоресцируют с максимумами возбуждения и эмиссии, соответственно, 410 и 480–490 нм. Для дифференцировки гистамина в ТК применен флуоресцентногистохимический метод Кросса-Эвана-Роста [15] с использованием ортофталевого альдегида (фирма «MERCK-Schuchardt»); в данном варианте метода гистамин флуоресцирует в желтой области спектра с максимумами возбуждения и эмиссии, соответственно, 365–410 и 515–520 нм.
С целью исключения аутофлуоресценции часть препаратов исследовалась без обработки параформом ортофталевым альдегидом. Препараты, приготовленные по вышеуказанным методам, изучались с помощью люминесцентного микроскопа ЛЮМАМ-И3, стандартного набора светофильтров, адекватных режиму флуоресценции биоаминов (ЖС-18, ФС 1-2, СЗС 24-4, БС 8-3). Цитоспектрофотометриче-ская установка ФМЭЛ-1А (выходное напряжение 2100В, ФЭУ-73), соединенная с регистрирующим устройством, с помощью отрезных интерференционных фильтров позволяет дифференцировать свечение серотонина, катехоламинов и гистамина и определять количественное содержание биоаминов (в условных единицах). На стандартной площади препарата подсчитывалось количество флуоресцирующих ТК. За одну варианту статистических массивов принималось количество клеток в 10 полях зрения.
Для выявления гликозаминогликанов в ТК осуществлена окраска криостат-ных срезов альциановым синим-сафранином в прописи Дезага [16]. В окрашенных мазках проводилась оценка плотности пространственного распределения ТК методом, аналогичным при подсчете флуоресцирующих клеток. Статистический анализ осуществлялся с помощью приложений платформы MS Exel.
Определялись коэффициенты параметрической и непараметрической корреляций. Достоверность различий математических ожиданий оценивали по критериям Стьюдента и Вилкоксона-Мании Уитни. Критический уровень значимости при проверке статистических гипотез принимался равным 0,05.
Результаты исследования и обсуждение. ТК в криостатных срезах, обработанных по методу Фалька-Хилларпа в модификации Е.М. Крохиной, определяются по характерной желтовато-зеленой флуоресценции гранул, располагаются они преимущественно одиночно. В эндо- метрии ТК располагаются преимущественно в соединительной ткани, некоторые из них локализуются под эпителием слизистой оболочки или непосредственно в нем (рис. 1). Плотность пространственного распределения флуоресцирующих ТК слизистой и мышечной оболочек тела матки минимальна в первые сутки после родов (рис. 2). В это время клетки располагаются в основном периваскулярно, цитоплазма большинства из них плотно заполнена гранулами. В некоторых тканевых базофилах наблюдаются признаки дегрануляции.
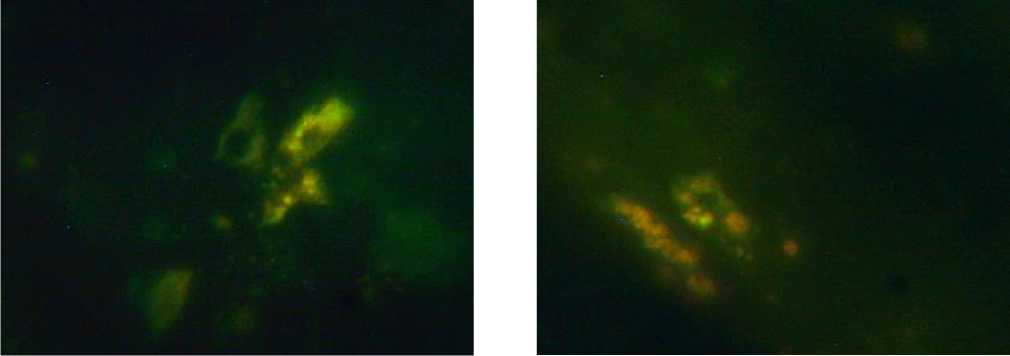
а б
Рис. 1. Тучные клетки эндометрия тела матки крыс. Обозначения: а – 2-е сутки после родов, б – 10-е сутки после родов. Окр.: метод Фалька-Хилларпа. Ув.: х900, масляная иммерсия.
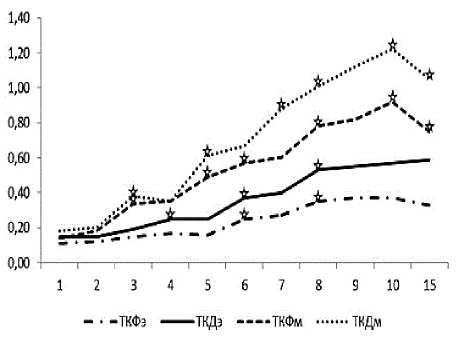
К 6-м суткам после родов плотность флуоресцирующих ТК в эндометрии увеличивается примерно в два раза, а в миометрии – в 4 раза (p<0,05). Визуально отмечаются клетки с выраженной дегрануляцией. Максимального уровня плотность ТК достигает максимума к 10-м суткам, за исключением эндометрия, где на 15-е сут-
Рис. 2. Динамика плотности пространственного распределения тучных клеток, флуоресцирующих (ТКФ) и окрашенных по методу Дезага (ТКД) эндометрия (э) и миометрия (м) тела матки крыс в течение послеродового периода. По оси ординат – средняя плотность пространственного распределения тучных клеток, по оси абсцисс – сутки после родов. Достоверность отличий оценочного параметра от такового в предыдущий период исследования p<0,05.
ки отмечается недостоверное увеличение этого параметра по сравнению с 10-ми сутками.
Плотность пространственного распределения ТК, выявляемых при окрашивании срезов альциановым синим-сафранином по Дезага, начиная с 4–5-х суток, достоверно выше, чем плотность флуоресцирующих тканевых базофилов. Динамика плотности клеток, выявляемых данным методом, в течение послеродовой инволюции матки аналогична изменению количества флуоресцирующих ТК (рис. 2).
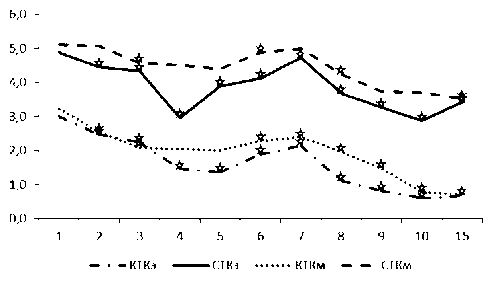
Рис. 3. Динамика содержания серотонина (СТК) и катехоламинов (КТК) в тучных клетках эндометрия (э) и миометрия (м) тела матки крыс в течение послеродового периода. По оси ординат – среднее содержание моноаминов в у.е., по оси абсцисс – сутки после родов. ☆ – достоверность отличий данного оценочного параметра от такового в предыдущий период исследования p<0,05.
Микроспектрофлуориметрия аль-дегид-индуцированного свечения ТК в препаратах, приготовленных по методу Фалька-Хилларпа в модификации Е.М. Крохиной, выявляет в них на протяжении всего послеродового периода наличие катехоламинов и серотонина. В тканевых базофилах эндометрия матки наибольшее содержание указанных биоаминов отмечается в первые сутки (рис. 3).
Сразу после родов уровень моноаминов постепенно достоверно снижается. В теле матки уменьшение содержания серотонина продолжается до 4-х суток, а катехоламинов – до 5-х, после чего отмечается увеличение этого показателя до 7-х суток. Минимальные значения уровня моноаминов наблюдаются на 10-е сутки.
В мышечной оболочке ТК чаще располагаются группами, иногда рядом с формирующимися нервными волокнами. Содержание серотонина и катехоламинов в ТК миометрия тела матки максимально в пер- вые сутки после родов (см. рис. 3). Постепенно снижаясь, оно достигает наименьших значений на 15-е сутки послеродовой инволюции.
При обработке криостатных срезов по методу Кросса-Эвана-Роста с целью вы-
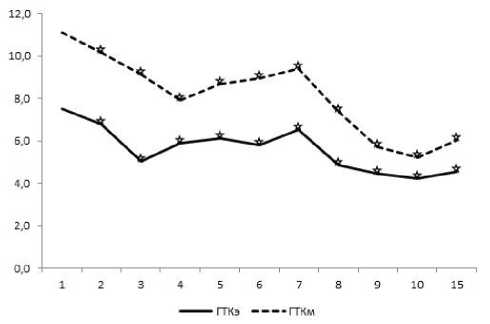
Рис. 4. Динамика содержания гистамина в тучных клетках эндометрия (э) и миометрия (м) тела матки крыс в течение послеродового периода. По оси ординат – среднее содержание моноамина в у.е., по оси абсцисс – сутки после родов. ☆ – достоверность отличий данного оценочного параметра от такового в предыдущий период исследования p<0,05.
явлении тканевого гистамина ТК характеризуются интенсивной зеленовато-желтой флуоресценцией гранул. Самый высокий уровень гистамина в ТК эндометрия приходится на первые сутки после родов (рис. 4). Начиная со вторых суток, содержание моноамина волнообразно достоверно снижается, достигая наименьших значений к 10-м и 15-м суткам. Необходимо отметить достоверное увеличение уровня гистамина на 7-е сутки. Аналогичным образом изменяется уровень гистамина в тканевых базофилах миометрия (см. рис. 4). Сравнивая насыщенность гистамином ТК различных оболочек матки в процессе ее послеродовой инволюции, можно отметить, что значимо большее его содержание выявляется в миометрии. Анализ величины коэффициента параметрической линейной корреляции между количеством серотонина и катехоламинов в ТК эндометрия и миометрия матки демонстрирует увеличение силы связи исследуемых параметров от средней (r=0,60–0,62) в первые сутки после родов до сильной (r=0,74–0,82) к 7-м суткам.
Данные рангового корреляционного анализа демонстрируют высокую силу положительной взаимосвязи изменений в течение послеродового периода практически всех сравниваемых показателей биоаминового статуса ТК. Так, для изменений количества серотонина и катехоламинов в ТК тела матки коэффициент корреляции составляет в эндометрии 0,873, в миометрии – 1,0. Из результатов рангового корреляционного анализа обращает внимание также положительная взаимосвязь изменений содержания гистамина и серотонина в ТК эндометрия тела (ρ=0,800), гистамина и катехоламинов (ρ=0,864). В ТК миометрия этот показатель выше – 0,955. Отрицательная высокая взаимосвязь характеризует изменения содержания моноаминов в ТК и плотности пространственного распределения тканевых базофилов (ρ=–0,600–0,982).
Таким образом, в процессе послеродовой инволюции происходит восстановление высокого уровня корреляционных связей содержания серотонина и катехоламинов в биоаминпозитивных элементах матки, отражая общую закономерность о равновесии процессов анаболизма-катаболизма в рамках гомеостаза клеток, тканей и органов [7-8]. Указанные моноамины являются антагонистами по многим аспектам своего биологического действия, поэтому их количество в нормально функционирующей матке должно быть сбалансировано. Оптимальное соотношение содержания биогенных аминов в микроокружении рабочих клеток создает в нем баланс обменных процессов, регенерационной и функциональной активности, влияет на адаптационные процессы [7].
Высокая насыщенность ТК эндометрия и миометрия биоаминами в первые сутки инволюции матки может быть связана с их повышенной активностью во время родов, когда они активно секретируют серотонин, гистамин, простагландины и другие биологически активные вещества [8]. Окситоцин замедляет поглощение серотонина ТК [13]. Возможно, он оказывает аналогичный эффект и на гистамин. Тканевые базофилы участвуют в обеспечении катехоламинами и серотонином структур миометрия [7]. Для гладких миоцитов этой оболочки матки они, видимо, являются основным поставщиком гистамина.
«После родов программируемый не некротический и не воспалительный процесс ремоделирования соединительной ткани матки приводит к быстрому расщеплению коллагена» (цит. по [8], c. 329), продукция которого в период беременности возрастает многократно. Этот возврат к нормальной структуре органа инициируется интерстициальной коллагеназой. Показана способность гладких миоцитов матки продуцировать этот фермент при стимуляции этого процесса серотонином [11]. Возможно, что с этим связано установленное нами выраженное уменьшение уровня серотонина в тканевых базофилах миометрия в первые сутки после родов, а также значительное увеличение плотности ТК в течение послеродового периода. Отмечается участие ТК в изменении цитоархитектоники эндометрия, ремоделировании кровеносных сосудов [13, 17-18], способность ТК выделять коллагеназу [8]. Это в совокупности с вышеуказанным определяет увеличение после родов плотности данных клеток в матке.
Значительное увеличение плотности тканевых базофилов в ранний послеродовый период может быть связано с отсутствием в первые сутки после родов флуоресцирующих нервных волокон. Установлено, что при денервации участков внутренних органов в них происходит увеличение числа ТК, компенсирующих, очевидно, утрату нервного влияния [19].
Возможно, что тканевые базофилы способствуют восстановлению симпатической иннервации матки в послеродовый период. Установлено, что многие цитокины и медиаторы, продуцируемые ТК, влияют на рост и дифференцировку нервных клеток [20]. Особое место во взаимодействии между ТК и нервными клетками занимают фактор роста нервных клеток (NGF) и гистамин, которые проявляют выраженную биологическую активность в отношении обоих типов клеток.
К 7-му дню структура популяции гладких миоцитов рогов близка к параметрам дородовой матки, что свидетельствует о завершении в ней инволюционных преобразований [4]. Интерес вызывает установленный нами факт, что к 7-м суткам после родов усиливается в миометрии дегрануляция ТК, повышается уровень в них гистамина и с этого дня значительно увеличивается плотность нервных волокон, как свободных, так и в составе периваскулярных сплетений. Значительное уменьшение уровня серотонина в ТК эндометрия и миометрия сразу после родов может быть связано с его участием в гемостазе путем вазоконстрикции маточных сосудов, в процессах послеродовой реконструкции тканей в ходе послеродовой инволюции матки [21].
Заключение. Таким образом, ТК в матке являются основным источником ги- стамина и дополнительным продуцентом серотонина и катехоламинов. Они также поглощают, накапливают и инактивируют излишки моноаминов, которые не были утилизированы рабочими клетками. Это объясняет характерные топографические кооперации тканевых базофилов и симпатических нервных волокон, а также кровеносных капилляров. Нельзя исключать и возможности выполнения ТК в ряде случаев функции переносчиков биоаминов между различными оболочками матки [7]. Можно предположить, ведущую роль ТК в паракринной регуляции гистофизиологи-ческих процессов, происходящих в матке в ходе ее послеродовой инволюции, особенно на ранних этапах. Возможно, что тканевые базофилы способствуют восстановлению симпатической иннервации матки в послеродовый период.
