Форма кроны Araucaria araucana (Molina) K.Koch в разных возрастных состояниях и экологических условиях естественных местообитаний
 K.Koch в разных возрастных состояниях и экологических условиях естественных местообитаний")
Автор: Казакова Наталья Леонидовна, Антонова Ирина Сергеевна
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 3, 2015 года.
Бесплатный доступ
В естественных местообитаниях штата Неукен, Аргентина, проведены геоботанические описания горных лесов по трансектам. Они проложены на склонах от пустошей дна межгорных долин (800 м над уровнем моря) до чистых араукарников (на высоте 1600 м). Выделены четыре типа лесных сообществ разного флористического состава с участием Araucaria araucana. Составлены их вертикальные профили. Форма крон деревьев A. araucana разных возрастных состояний подробно описана по ряду признаков. Установлена зависимость формы кроны от экологических условий сообщества. Показано, что положение араукарии чилийской в фитоценозе и конкурентные отношения в нём в значительной степени трансформируют форму кроны в конкретном возрастном состоянии.
Араукария, аргентина, неукен, растительное сообщество, профиль растительного сообщества, возрастное состояние, онтогенетическая стадия, форма кроны, псевдомутовка, прирост, побег
Короткий адрес: https://sciup.org/146116589
IDR: 146116589 | УДК: 581.5(828.1);
The crown shape of Araucaria araucana (Molina) K.Koch under the different age stages and ecological conditions in natural habitats
The geobotanical studies on transects of the natural mountain forests in the State of Neuquen, Argentina have been carried out. Transects were established on the slopes, starting from the clearings at the bottom of valleys (800 m above sea level) to the true Araucarian forests (1,600 m). Four types of forest communities of different floristic composition with the participation of Araucaria araucana have been defined. Vertical profiles of the mentioned forests have been drawn. Crown architecture of A. araucana was described in details at the different age states of trees. The dependence of the crown's shape from the environmental conditions of the phytocenosis was revealed. The position of A. araucana in the phytocenosis and its relations inside it greatly influence its crone shape in each particular age.
Текст научной статьи Форма кроны Araucaria araucana (Molina) K.Koch в разных возрастных состояниях и экологических условиях естественных местообитаний
Введение. Успешность выживания растения в конкретных условиях обитания зависит от того, как протекает реализация его генетической программы развития, проявляющаяся в последовательности морфогенетических состояний. Здесь имеют значения две особенности развития: первая - жизнеспособность растений в данный возрастной период, вторая - длительность протекания той или иной стадии. Изучение изменений формы кроны в разных возрастных состояниях может показать, как и на какой именно стадии растение реагирует на абиотические, биотические или антропогенные факторы или их комплекс. Форма кроны как комплексный ответ растения на воздействие среды в рамках реализации генетической программы изучается в России со второй половины XX в. В настоящее время это направление исследований актуально для крон деревьев умеренной зоны (Антонова и др., 2012; Антонова, Фатьянова, - 135-
2013, 2014). Выявление особенностей строения кроны, позволяющее выделять и узнавать объект в разнообразии фенотипических реализаций, предлагаемых природой, становится важной общебиологической задачей.
С позиций выявления особенностей формы кроны в разных возрастных состояниях дерева интересно исследовать одно из древнейших древесных растений Araucaria агаисапа, в естественных местообитаниях провинции Неукен, Аргентина. В связи с этим нашей целью было составление характеристики формы кроны А. агаисапа в естественных местообитаниях в разные возрастные периоды.
История вопроса. Агаисапа агаисапа - один из древнейших видов древесных растений, с просто организованной архитектурой кроны, которая, тем не менее, позволяет ей успешно существовать и адаптироваться к разным условиям среды (Grosfeld et al, 1999; Tomlinson, 2002). У молодых деревьев в благоприятных условиях ветви размещены вдоль всего ствола, от самого его основания. Обычно они отходят от ствола сближено, почти на одном уровне, «мутовками» по 37 ветвей. Гросфельд с сотрудниками дали таким образованиям название «псевдомутовки» (Grosfeld et al, 1999). Направление роста ветви горизонтально, со слегка поднятыми вверх концами. По мере старения ветви обвисают (Donoso, 1997). С возрастом араукарии, как правило, сбрасывают ветви в нижней части ствола, постепенно этот процесс охватывает дерево почти до самого верха, стволы оказываются увенчанными уплощенными зонтиковидными кронами (Hoffmann, 1991; Grosfeld, 1994; Marticorena, Rodrigues, 1995; Grosfeld et al., 1999; Tomlinson, 2002).
Архитектуру кроны А. агаисапа многие авторы определяли моделями Массарт и Раух (Halle et al., 1978; Veillon, 1978; Veillon, 1980; Tomlinson, 2002).
Морфологические признаки характерные для деревьев А. агаисапа разного возраста и мест произрастания были описаны Гросфельдом с сотрудниками в 1994 (Grosfeld et al, 1994). Эти исследователи выявили 3 порядка ветвления, включая ствол и развитие генеративных органов на осях третьего порядка. Образование ветвей второго и третьего порядка у А. агаисапа происходит не ежегодно. Количество ветвей второго порядка, образующихся в одной псевдомутовке, увеличивается с возрастом (до 8 шт.). Ветви араукарии чилийской второго и/или третьего порядка могут продолжать свой рост, несмотря на спящую апикальную меристему главной оси. Выявлены три морфологические стратегии роста молодой араукарии в различных местообитаниях: 1) в подлеске - типичная зонтиковидная форма с тонкими ветвями, 2) в условиях полного освещения, богатых почв и достаточного увлажнения - пирамидальная форма и 3) на бедных почвах, при малом количестве осадков и обильном освещении -зонтиковидная форма с крепкими ветвями.
По мнению Бюрнса и Гросфельда, зонтиковидная форма араукарии в подлеске может быть формой ожидания благоприятных для развития условий (Burns, 1991; Grosfeld, 1999). Отмечено, что угнетенные стволики могут отмирать, а из спящих почек в их комлевой части формируются новый ствол с кроной зонтиковидной формы. Вывал деревьев верхнего яруса может спровоцировать рост особей подлеска. Дальнейшее их развитие пойдет в естественном ритме. Максимальное количество «псевдомутовок», образующееся на дереве в условиях хорошего роста, 18-20 шт. По мере дальнейшего развития в генеративном возрастном состоянии крона приобретает конусовидную форму; в условиях хорошего освещения, достатка осадков и богатства почв - пирамидальную крону, а в условиях бедных почв и недостатка осадков при хорошем освещении формируется типичная зонтиковидная форма (Grosfeld et al., 1999). Максимальное количество мегастробилов образующихся на ветви - 5 шт., после чего ветвь продолжает свое существование только в вегетативном состоянии. У особей мужского пола нарастание ветви после образования генеративного органа моноподиальное, у особей женского пола - симподиальное. Для синильных особей араукарии отмечается зонтиковидная форма высоко поднятой кроны, с переходом от билатерально к радиальному положению ветвей 3 порядка. Для Л. агаисапа отмечается реитерация (общая или частичная) как ответ на нарушения или без видимых на то причин (ответ на среду). У особей, растущих в тяжелых условиях, обнаруживается вегетативная мультипликация, которая с помощью корней компенсирует недостаток семенного возобновления (Grosfeld et al., 1999; Tomlinson, 2002). Многочисленные исследования, проведенные Бурроусом, показали наличие большого количества резервной меристемы в теле дерева (Burrows 1987, 1990, 1999).
Зависимость плодоношения Л. агаисапа от факторов окружающей среды, синхронизация плодоношения по годам удаленных друг от друга популяций араукарии чилийской, а также влияния локальной среды на эти оценки были выявлены Сангвинетти и Китсбергером (Sanguinetty, Kitzberger, 2008).
Прирост молодых растений (до 30-150-летнего возраста) может достигать 45 см ежегодно, а затем уменьшается до 10-15 см (Муравьева, 1974). В естественных насаждениях Л. агаисапа, смешанных с Nothofagus pumilio (Poep.et Endl), отмечают прирост от 5 до 8,2 см/год, в диаметре от 2,34 до 2,7 мм в год. Темпы роста данного вида, как правило, от 1 до 2,28 м3/га в год (Schmidt et al, 1980; Cavieres, 1987). Исследования, проводимые Драком с сотрудниками, направленные на разработку методов рационального пользования араукариевыми лесами, говорят о необходимости поддерживать смешанные араукариево-нотофагусовые леса (Drake et al, 2005). По их оценкам, именно там араукарии находят лучшие условия для своего возобновления и роста, поддерживая при этом разновозрастность сообщества. Проблему семенного возобновления авторы видят в малой площади рассеивания семян под материнским деревом (Munos, 1984; Vita, 1996; Drake, 2005).
Люск и Ле-Кесн рассматривали вопрос о периодичности образования псевдомутовок у молодых особей Л. агаисапа, основываясь на приросте годичных колец и количестве псевдомутовок у молодых особей араукарии в условиях полного освещения и затенения (Lusk, Le-Quesne, 2006). На всех наблюдаемых особях образование пседомутовок не совпадало с приростом годовых колец и не было ежегодным. В условиях хорошего освешения псевдомутовки образовывались в среднем раз в 2 года на открытых местах и раз в 3 года в условиях леса. Констатирован лучший прирост ствола между последовательными ветвлениями в хорошо освещённых местообитаниях. Количество образующихся псевдомутовок, по мнению авторов, является лучшим неинвазивным методом для определения возраста молодых особей (Lusk, Le-Quesne, 2008). Гросфельд с сотрудниками также отмечают неежегодное образование псевдомутовок в затененных условиях; апикальная меристема затененных молодых араукарий может находиться в спящем состоянии несколько лет (до 9), а затем продолжить развитие оси (Grosfeld et al., 1999; Tomlinson, 2002).
На западных склонах Анд в среднем и высоком поясе араукария образует насаждения в смеси с N. pumilio и N. antarctica; с увеличением высотности присутствие этих пород уменьшается, и формируются чистые насаждения (Donoso, 1993; Veblen 1982). В низком горном поясе араукарии отмечаются в смеси с N. dombeyi (Mirb.) Oerst и N. oblique (Mirb.) Oerst (Donoso, 1981, Galardo, 1980; Veblen et al, 1995). Ha восточных склонах Анд араукарии образуют чистые леса и отдельные островные участки с Austrocedrus chilensis (D.Don) Pic.Senn, et Bizzarri в «степях» Патагонии (Burns, 1991).
Район и методы исследования. Сбор материала производился на западной границе провинции Неукен, у оз. Мокуэуэ, на высотах 8001600 м над уровнем моря. Количество осадков в районе исследований варьирует от 800 до 2000 мм (Grosfeld et al, 1999; Donoso, 2006). Исследование проводилось в ноябре 2013 г. В результате рекогносцировочных работ были получены следующие сведения: 1) общая ландшафтная характеристика района, 2) состав основных лесообразующих видов, разнообразие и распределение различных типов лесных сообществ1, 3) факторы внешнего нарушения и примерная степень нерушенности лесных сообществ. Кроме того, были проанализированы литературные и картографические материалы по данной территории. Проложены три трансекты по склонам гор, окружающим оз. Мокуэуэ; первая и вторая трансекты располагались на склоне восточной экспозиции, от долины до чистого араукариевого леса, третья, аналогичная, была на западном склоне. Выбор места для заложения пробных геоботанических площадей производился в соответствии с изменениями растительного покрова верхних лесных ярусов в типичных местообитаниях. В каждой трансекте были заложено по три геоботанических площади, размером 20x20 м. Кроме того, была исследована геоботаническая пробная площадь такого же размера на равнинной части (750 м над уровнем моря). На заложенных участках сделаны общие описания леса с учетом следующих показателей: координаты площадей по GPS, уклон, экспозиция, высоты над уровнем моря, формула древостоя, ярусность, средние и максимальные высоты деревьев разных видов, разных ярусов, средний и максимальный диаметр деревьев разных ярусов и видов, форма и высота поднятия кроны, сомкнутость, сквозистость, видовой состав подлеска, его проективное покрытие и высота. Описаны восстановления подроста в пределах каждой геоботанической площадки (взяты по 3 участка 3x3 м), произведен подсчет подроста разных групп (ювенильные растения - до 0,15 м; ранние имматурные растения четырёх категорий по высотам -до 0,5 м; от 0,5 до 1,5 м; от 1,5 до 3 м; более 3 м). Количество проанализированных растений подроста на различных стадиях составляет 307 шт. Выполнены подробные биоморфологические описания подроста и формы крон А. агаисапа в разных возрастных состояниях. Проведены описания и измерения всех побегов трех модельных деревьев. Впервые на основе методики, предложенной для деревьев умеренной зоны на территории России (Смирнова и др., 1984; Восточноевропейские..., 1994), выделены возрастные состояния для Л. агаисапа.
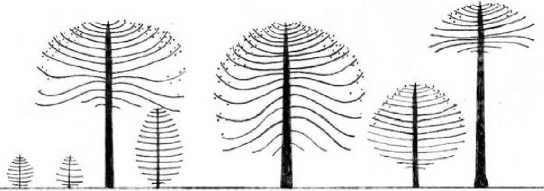
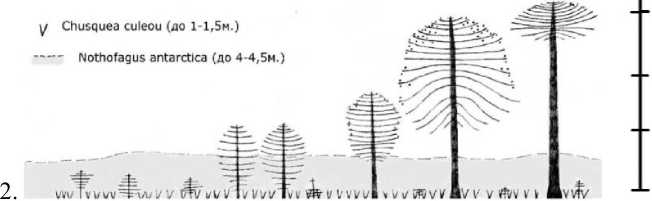


Рис. 1 . Типичный профиль растительного сообщества. 1. Пустошь с Araucaria агаисапа; 2. Араукарно-южнобучник бамбуковый; 3. Южнобуково-араукарник бамбуковый; 4. Араукарник кустарниковый
Результаты и обсуждение. Анализ собранного материала позволил выделить 4 типа местообитаний А. агаисапа на разной высоте в характерных для провинции лесорастительных условиях: долинная пустошь с араукарией и 3 типа сомкнутых сообществ - араукарно-южнобучник бамбуковый (Агаисапа агаисапа - Nothofagus antarctica -Chusquea culeou), южнобуково-араукарник бамбуковый (Araucaria агаисапа + Nothofagus pumilio, Nothofagus dombeyi - Chusquea culeouf араукарник кустарниковый (Araucaria агаисапа - Chusquea culeou, Pernettya mucronata (L.F.) Hook et Arn, Maytenus magelanica (Lam.) Hook.f., Myginda disticha (Hook.f.) J.B.Urban, Tepualia stipularis (Hook, et Arn.) Griseb.) (рис. 1).
Tаблица 1
Признаки кроны А.агаисапа в исследованных местообитаниях
|
Местообитания |
Пустошь с араукарией |
Араукарно-южнобучник бамбуковый |
Южнобуково-араукарник бамбуковый |
Араукарник кустарниковый |
|
Синильное возрастное состояние S1-S2" |
||||
|
высота дерева, м |
15-17 |
18-20 |
23-25 |
25-30 |
|
диаметр ствола, см |
65-75 |
62-64 |
80-85 |
92-98 |
|
протяженность кроны, м |
4-4,5 |
3,8-4,2 |
3-4 |
3-3,5 |
|
проекция кроны |
округлая |
округлая |
неправильно многоугольная |
неправильно многоугольная |
|
количество псевдомутовок, шт |
13-14 |
12-13 |
12-13 |
11-12 |
|
расположение генер. органов в кроне (псевдомутовки обозначены как «псм») |
4 верхние псм, единичные |
4 верхние псм, единичные |
3 верхние псм, единичные |
4 верхние псм, единичные |
|
радиус кроны, м |
3,5 - 4,2 |
3,8 - 4,5 |
3,5 - 4,5 |
3,2 - 4,2 |
|
форма кроны |
зонтиковидная 1, 2 |
зонтиковидная 1, 2 |
зонтиковидная 1, 2 |
зонтиковидная 1, 2 |
|
G2-G3 Генеративные возрастные состояния |
||||
|
высота дерева, м |
12-14 |
15-17 |
15-20 |
19-20 |
|
диаметр, см |
54-56 |
53-56 |
60-63 |
63-69 |
|
протяженность кроны, м |
9-10 |
7-8 |
5-6 |
4-5,5 |
2 Возрастные состояния выделены на основе методики, приведенной в книге «Восточноевропейские широколиственные леса», 1994.
- 141 -
|
Местообитания |
Пустошь с араукарией |
Араукарно-южнобучник бамбуковый |
Южнобуково-араукарник бамбуковый |
Араукарник кустарниковый |
|
проекция кроны |
округлая |
округлая |
неправильно многоугольная |
неправильно многоугольная |
|
количество псевдомутовок, шт |
15-22 |
17-22 |
12-20 |
12-18 |
|
радиус кроны, м |
4,5 - 7 |
4,2 - 5 |
4,2 - 5,5 |
4,0 - 5,0 |
|
форма кроны |
шарообразная |
овальная |
зонтиковидная 2, 3 |
зонтиковидная 1, 2,3 |
|
G1 Генеративное возрастное состояние |
||||
|
высота дерева, м |
9-12 |
8-11 |
12-15 |
15-19 |
|
диаметр, см |
35-40 |
35-40 |
40-46 |
44-48 |
|
протяженность кроны, м |
6-7,5 |
5-6 |
3,5-4 |
3,5-4,5 |
|
проекция кроны |
округлая |
округлая |
неправильно многоугольная |
неправильно многоугольная |
|
количество псевдомутовок, шт |
16-18 |
14-18 |
10-14 |
10-13 |
|
расположение генер. органов в кроне |
1/2 кроны (середина, верх) |
1/3 кроны (верх) |
единично в верхней части |
единично или отсутствуют |
|
радиус кроны, м |
3,4 до 3,8 |
2,9 до 3,5 |
2,0 до 2,8 |
2,2 до 2,8 |
|
форма кроны |
широко яйцевидная |
яйцевидна, эллиптическая |
зонтиковидная 2,3 |
зонтиковидная 1,2,3 |
|
Virg. Виргинильное возрастное состояние |
||||
|
высота дерева, м |
6-8 |
6-8 |
7-12 |
10-15 |
|
диаметр, см |
32-33 |
23-28 |
35-40 |
40-45 |
|
протяженность кроны, м |
5,5-7,5 |
5,5-7,0 |
3-4,5 |
3-4,5 |
|
проекция кроны |
округлая |
неравномерная с тенденцией к округлой |
неправильно многоугольная |
неправильно многоугольная |
|
количество псевдомутовок, шт |
12-17 |
10-16 |
3-10 |
6-10 |
|
расположение генер. органов в кроне |
нет |
нет |
нет |
нет |
|
радиус кроны, м. |
1,5 - 2,0 |
1,0- 1,5 |
1,2 -2,0 |
1,2 - 2,2 |
|
форма кроны |
эллиптическая |
обратно яйцевидная |
зонтиковидная 1, 2, 3; обратно пирамидальная; |
зонтиковидная 1, 2, 3; обратно пирамидальная; |
|
1mm. Имматурное возрастное состояние |
||||
|
высота дерева, м |
3-6 |
3-6 |
3-7 |
3-10 |
|
диаметр, см |
16-18 |
16-18 |
15-23 |
15-26 |
|
протяженность кроны, м |
3,5-4,5 |
2-3,5 |
0,8-3,2 |
0,8-3,5 |
|
Местообитания |
Пустошь с араукарией |
Араукарно-южнобучник бамбуковый |
Южнобуково-араукарник бамбуковый |
Араукарник кустарниковый |
|
проекция кроны |
округлая |
неправильно многоугольная |
неправильно многоугольная |
неправильно многоугольная |
|
количество псевдомутовок, шт |
8-10 |
6-10 |
4-8 |
3-9 |
|
расположение генер. органов в кроне |
нет |
нет |
нет |
нет |
|
радиус кроны, м |
1,2 до 1,5 |
1,0 до 1,3 |
1,0 о 1,3 |
1,1 до 1,5 |
|
форма кроны |
эллиптическая с уменьшением длин к земле |
эллиптическая; конусовидная; зонтиковидная 3 |
зонтиковидная 1, 2,3 |
зонтиковидная 1, 2,3 |
|
1mm. Подрост менее Зх метров |
||||
|
форма кроны |
эллиптическая, конусовидная |
эллиптическая, конусовидная, зонтиковидная 3, прямоугольная, ромбовидная, паукообразная |
зонтиковидная 1,2,3, паукообразная, обратно пирамидальная |
зонтиковидная 1,2,3, паукообразная, обратно пирамидальная, многоствольная |
Таблица 2
Формы кроны Л. агаисапа, выделенные в типичных местообитаниях провинции Неукен
|
Зонтиковидная 1 (до 10%)* |
Зонтиковидная 2 (10-20%)** |
Зонтиковидная 3 (20-30%)*** |
||||||
|
Шаровидная |
Овальная |
Обратно яйцевидная |
||||||
|
Яйцевидная |
Эллиптическая |
-- |
Пирамидальная |
|||||
|
Обратно пирамидальная |
Паукообразная |
Ромбовидная |
||||||
|
И. |
Многоствольная |
Примечание. *, **, *** - пояснения в тексте.
Названия лесных сообществ даны на основе применения эколого-фитоценотического принципа классификации лесных сообществ (Методы изучения лесных сообществ, 2002). При выделении основных классификационных единиц на первый план выдвигаются признаки растительного сообщества, хотя значение придается и комплексу факторов местообитания.
Сопоставление форм кроны у последовательно сменяющихся стадий от зрелой генеративной до имматурной показывают, что пирамидальная форма кроны Л. агаисапа встречается редко и только в молодом возрасте. Зонтиковидные формы, выделенные нами при описании крон А. агаисапа, различаются по соотношению высоты кроны к общей высоте дерева: 1) 10% и меньше от общей высоты дерева, 2) 11 - 20% и 3) 21 - 30% (табл. 2).
Во всех типах лесорастительных условий синильные особи (т. е. деревья прекращающие или прекратившие плодоношение) имеют зонтиковидные кроны (рис. 1: 1, 2). В зависимости от местообитания они значительно отличаются по высоте дерева и диаметру. При этом радиус (около 4 м), высота кроны (3 - 4 м) и количество сближенных псевдомутовок (11 - 13 шт) остаются примерно одинаковыми. Формы крон синильных особей наиболее похожи во всех типах местообитаний (табл. 1).
Сходные формы крон в среднем возрастном и старом генеративном возрастных состояниях (G2 - G3) наблюдаются в местообитаниях долинной пустоши с А. агаисапа (1) и южнобукового леса с бамбуком (2) (рис. 1). На открытых местообитаниях долинная пустошь с А. агаисапа крона шарообразная и более широкая (до 7 м). В сообществах с N. antarctica крона деревьев араукарий возвышается над пологом нотофагуса, она уже и более вытянута (до 5 м). Количество генеративных органов, образуемых в этих условиях, наибольшее в сравнении со всеми приведенными местообитаниями. В среднем возрастном и старом генеративном возрастных состояниях (G2-G3) в сообществах южнобуково-араукарного леса с бамбуком (3) и араукарника кустарникового (рис. 1: 4) геометрия крон мало различается и представлена зонтиковидной формой в ее 2, 3 вариантах. В сообществе южнобуково-араукарного леса с бамбуком (рис. 1: 3) в данных возрастных состояниях наблюдаются разнообразные проявления вторичного роста, ветвей кроны (табл. 1). Особи синильного и генеративного возрастных состояний отличаются наличием или отсутствием плодоношения, стадия G2 характеризуется активным плодоношением, G3 - уменьшением количества генеративных органов, нерегулярным плодоношением, в синильном состоянии S1
Вестник ТвГУ. Серия "Биология и экология". 2015. № 3 плодоношение прекращается, образуются единичные генеративные органы, в S2 плодоношения нет, крона активно изреживается, начинаются процессы отмирания дерева (Чистякова и др. 1979). По нашим наблюдениям, синильное возрастное состояние у A. araucaria может растягиваться на сотни лет.
Форма крон деревьев в молодом генеративном возрастном состоянии G1 в местообитаниях долинной пустоши с А. агаисапа (рис. 1: 1) (широко яйцевидная) и араукарно-южнобукового леса с бамбуком (рис. 1: 2) (яйцевидная, эллиптическая) отличается радиусом и высотой зеленой части кроны, а также количеством генеративных органов и количеством сохранившихся фотосинтезирующих нижних ветвей; в сообществах с N. antarctica их меньше. Форма крон G1 для сообществ нотофагусово-араукарного леса с бамбуком (3) и араукарника кустарникового (рис. 1:4) — зонтиковидная в вариантах 2, 3, на участке 4 (рис. 1) встречаются особи со сплющено зонтиковидной 1 формой кроны (табл. 1).
Виргинильные особи в сообществах долинной пустоши с А. агаисапа (рис. 1: 1) и араукарно-южнобукового леса с бамбуком (рис. 1: 2) имеют эллиптические и обратно яйцевидные формы соответственно, различающиеся радиусом кроны. В сообществах южнобукового-араукарного леса с бамбуком (рис. 1: 3) и араукарнике кустарниковом (рис. 1: 4) габитусы виргинильных деревьев совпадают - зонтиковидные в вариантах 1, 2, 3 и обратно пирамидальные (табл. 1, 2).
Имматурное возрастное состояние в сообществе долинной пустоши с А. агаисапа (рис. 1: 1) представлено эллиптической или яйцевидной формой кроны. В данном местообитании с самых первых этапов наблюдается ровный, последовательный рост, без признаков угнетения вследствие затенения другими растениями. В сообществе араукарно-южнобукового леса с бамбуком (рис. 1: 2) выявлено наибольшее разнообразие форм крон. Здесь существует два полога основных конкурентов: заросли бамбука С. culeou и ярус кроны N. antarctica. Полог, образованный С. culeou, характеризуется средней высотой 1,2 м (max -1,7 м), его проективное покрытие достигает 70-80 %, местами - до 40-50%. Встречаются небольшие по площади вытоптанные скотом участки, в целом бамбук образует сомкнутый покров. Полог, образованный N. antarctica, в среднем имеет высоту 4 м, кроны смыкаются в единый покров, в нижней части полога много отсохших частей растения. Нотофагус антарктический -многоствольное дерево с рыхлой древесиной, искривлённым стволом, нередко имеет кустовидную форму кроны.
Кроны молодых деревьев А. агаисапа проявляют вариативность в форме, изменяются по мере увеличения возраста (паукообразные, ромбовидные, эллиптические, прямоугольные, зонтиковидные в варианте 3 (рис. 1). В сообществах южнобуково-араукарного леса с бамбуком (рис. 1: 3) и араукарнике кустарниковом (рис. 1: 4) формы крон имматурных растений зонтиковидные 1, 2, 3, обратно пирамидальные, паукообразные, в сообществе 4 (рис. 1) появляются многоствольные особи (табл. 2). Именно здесь растения испытывают сильный недостаток света и питания. Эффекты многоствольности молодых деревьев А. агаисапа отмечались и ранее в работах Гросфельда с сотрудниками и Томлинсона (Grosfeld et al., 1999; Tomlinson, 2002). Условия семенного возобновления на 4 участке самые сложные. Несмотря на большое количество опадающих семян, в чистых кустарниковых араукарниках прорастание и выживаемость проростков и подроста невысокая, подрост отмирает на ранних стадиях своего существования и характеризуется малой жизненностью (Казакова, Антонова, 2014). В условиях смешанного с нотофагусами араукариевого леса количество опадающих семян уменьшается с долей участия араукарии в составе древостоя, но, тем не менее, семена здесь имеют лучшие показатели прорастания, и присутствует разновозрастной подрост.
Обсуждение. Гросфельдом с сотрудниками и Томлинсоном (Grosfeld et al., 1999; Tomlinson, 2002) было дано подробное описание морфологии А. агаисапа; в качестве архитектурных единиц роста кроны были выделены порядки ветвления, ярусная организация роста, «псевдомутовки». Ими было сделано описание организации ветвей и расположение репродуктивных органов на мужских и женских деревьях. Гросфельд и его сотрудники (Grosfeld et al, 1999) отметили различия в положении репродуктивных органов для мужских и женских экземпляров А. агаисапа, а также особенности дальнейшего роста ветви после акта размножения. Женские шишки расположены терминально на осях третьего порядка, последующий рост ветви продолжается симподиально из точки, расположенной под макростробилом. Одна ветвь третьего порядка может образовывать до 5 макростробилов, после чего переходит к вегетативному существованию с последующим отмиранием. Микростробилы образуются также на осях третьего порядка, и располагаются латерально, образуя группу (псевдомутовку) из 2 - б шишек. После пыления нарастание ветви моноподиальное. Количество псевдомутовок микростробил на одной ветви - до б, в дальнейшем ветвь также ведет вегетативное существование с последующим отмиранием. Наши наблюдения и описания
Вестник ТвГУ. Серия "Биология и экология". 2015. № 3 репродуктивных различий полностью соотносятся с данными литературы (Grosfeld et al., 1999). Остается открытым вопрос временного интервала периодов между последовательными репродуктивными актами на ветви. Репродуктивные органы в кроне дерева всегда расположены на хорошо освещенных ветвях и, в зависимости от места произрастания особи (в пологе леса или на открытом пространстве), могут располагаться либо на самых верхних ветвях, либо быть рассеяны по всей контурной площади кроны.
Гросфельд и сотрудники обратили внимание на особенности развития дерева в разных условиях окружающей среды (Grosfeld et al., 1999): на открытых местообитаниях, в пологе леса и на скальных участках, при разном количестве света, воды и богатства почв. Это позволило им подтвердить мнение многих авторов о том (Halle, Oldeman, 1970; Halle et al., 1978; Barthelemy, 1988; Masotti et al., 1995; Barthelemy et al., 1997), что окружающая среда не влияет на элементарные последовательности развития вида, но моделирует их выраженность. Авторами была выделена высокая вариативность развития деревьев в переходных возрастных состояниях.
В целом наши наблюдения схожи с описаниями, представленными ранее (Grosfeld et al, 1999; Tomlinson, 2002), но вносят некоторые уточнения в описание геометрии форм крон А. агаисапа. Так называемая авторами пирамидальная форма кроны в условиях открытых местообитаний была нами определенна как эллиптическая и даже, в условиях полного освещения и достатка воды, как шаровидная. Зонтиковидность синильных особей, независимо от условий среды, нами подтверждается. В зависимости от стадии синильности дерева были выделены три последовательные стадии зонтиковидности (табл. 2) по процентному отношению высоты кроны к общей высоте дерева.
Растения, произрастающие в пологе леса, имеют тенденции к зонтиковидности кроны (табл. 2); преобладание той или иной формы кроны дерева зависит от породного состава древостоя. Дрейк и его коллеги (Drake et al., 2005) уже отмечали, что А. агаисапа находит лучшие условия для своего прорастания и роста в смешанных с нотофагусами насаждениях. Однако в таких смешанных лесах (до высот 1300 - 1350 м) А. агаисапа сталкивается с острой конкуренцией за пространство, воду и свет с С. culeou (Donoso, 2006). Плотные заросли бамбука в нижнем ярусе леса создают условия, в которых молодые ювенильные и имматурные особи А. агаисапа проявляют большую вариативность в геометрии своих крон. Они активно реагируют на положение в пространстве изменением формы. Нами отмечены 9 типов форм крон для имматурных растений в условиях смешанных лесов с разными видами рода нотофагус. Различия форм крон А. агаисапа в условиях леса с N. antarctica и с N. dombeyi, N. obliqua, определяются на виргинильной стадии. В лесу с N. antarctica, молодые деревья, переросшие полог бамбука и N. antarctica (около 4 м), получают достаточно света и образуют обратно яйцевидную или эллиптическую форму кроны (табл. 2). Под пологом в смешанном лесу с N. dombeyi и N. pumilio возникновение такой формы - исключение, связанное с возможным вывалом более высокого дерева. В таких условиях крона образует зонтиковидные или обратно-пирамидальную формы. Кроме того, при большей общей высоте дерева в условиях полога высота кроны и количество псевдомутовок меньше.
При переходе в другие возрастные состояния заданные в виргинильной стадии различия формы кроны сохраняются. Возникают отличия проекций крон в условиях леса с N. antarctica и с TV. pumilio, N. dombeyi. В первом случае, от виргинильного возрастного состояния до синильного проекция кроны округлая. В условиях леса с N. dombeyi и N.pumilio проекция кроны становится неправильно многоугольной, начиная с имматурной стадии. Таким неравномерным вытяжением ветвей дерево реагирует на поток света и межкроновое пространство.
В связи с тем, что А. агаисапа имеет только три порядка ветвления, реакция её кроны на воздействие внешней среды проявляется в количестве псевдомутовок и расстоянии между ними, общей протяженности кроны по стволу, высоте дерева, соотношении общей высоты дерева и протяженности кроны.
На уровне ветви ответом на внешние факторы являются изменения ее длины, наличие регулярного ветвления третьего порядка, вариабельность расположения осей в пространстве, а также различия в продолжительности жизни осей третьего порядка и их сохранность на осях второго порядка.
На ранней стадии своего развития рост кроны молодого дерева зависит от периодов работы апикальной меристемы, как и отмечали ранее другие авторы (Grosfield et al., 1999; Tomlinson, 2002); периоды активного роста и состояния покоя (до 9 лет) апикальной меристемы напрямую зависят от условий роста дерева. При недостатке света, питательных веществ, воды или пространства деревья чаще всего образуют форму зонтика с 1 - 2 ярусами живых ветвей в кроне. Томлинсон (Tomlinson, 2002) такую форму называет паукообразной (тонкие ветви, практически полное отсутствие осей третьего порядка, тонкие и острые листья). Кроме того, Гросфельд с сотрудниками (Grosfeld et al., 1999) отмечали переход от билатеральной симметрии к радиальной по мере старения дерева.
В условиях чистых араукарников преобладающей формой кроны является зонтиковидная. Отмечено 6 форм крон в ранней имматурной стадии (до 3 м). При переходе к поздней имматурной стадии (более 3 м) вариативность уменьшается (табл. 2). Здесь же выявлены многоствольные особи, что подтверждает наблюдения других авторов (Grosfield et al., 1999; Tomlinson, 2002). Авторами также отмечается порослевой рост и множественность реитерации, возникающая из ствола или корней и замещающая отмерший стволик. Так же, как и Гросфельд с сотрудниками и Томлинсон (Grosfield et al., 1999; Tomlinson, 2002), считаем зонтиковидную, паукообразную и многоствольную формы стадиями ожидания растением благоприятных для роста условий.
Вопрос об определении фактического возраста дерева араукарии чилийской неинвазивными методами вызывает значительные затруднения. Единственным точным способом его определения является пересчет годовых колец. Люск и Ле-Кесн (Lusk, Le-Quesne, 2008) проводили исследования периодичности образования псевдомутовок на ювенильных деревьях (1,5 - 6,7 м). Ими было установлено, что деревья этого возраста в условиях хорошего освещения образуют ярус ветвей в среднем 1 раз в 2,1 года. У деревьев в условиях подлеска ярус образуется раз в 2,6 года. Полученные результаты применимы только для условий исследованного региона и только для ювенильных особей. Периодичности образования псевдомутовок для особей в возрастных состояниях imm., virg., Gl, G2, S не установлены. Определение стадии онтогенеза является единственным неинвазивным способом примерного определения возраста дерева араукарии чилийской с учетом мест и условий его произрастания. Проблема неинвазивного определения возраста дерева А. агаисапа остается открытой и требует дальнейшего исследования.
Заключение. Впервые описаны формы крон Araucaria агаисапа разных возрастных состояний по методике, предложенной для деревьев умеренной зоны (Восточноевропейские..., 1994; Смирнова и др., 1984), в типичных местообитаниях вида. Показана поливариантность форм кроны в зависимости от условий среды. В сообществе с N. antarctica выделено наибольшее количество форм крон имматурного возрастного состояния растений, что связано с необходимостью преодоления двух пологов конкурентов - С. culeou и N. antarctica. Только преодолев оба полога, крона А. агаисапа приобретает симметричную наполненную ветвями эллиптическую форму. Формирование эллиптической или шаровидной формы кроны араукарии происходит на открытых местообитаниях значительно раньше, чем в сообществе. В сомкнутых сообществах южнобуково-араукарного леса с бамбуком такую форму можно наблюдать только на вывалах и на опушках. В сообществах кустарниковых араукарников благоприятных условий для развития эллиптической формы кроны не образуется. В связи с этим абрис дерева, сформировавшийся в имматурном возрастном состоянии, в условиях чистого или смешанного леса (с N. pumilio, N. dombeyi) практически не меняется (форма зонтиковидная 1, 2, 3). Габитус Л. агаисапа в синильном возрасте одинаков для всех местообитаний (зонтиковидная 1, 2). Форма крон отражает высокую адаптивную способность к условиям среды и большое светолюбие вида. Наибольший приспособительный потенциал кроны для араукарии чилийской отмечается на ранних имматурной и виргинильной стадиях развития. Наибольшая геометрическая пластичность формы кроны свойственна А. агаисапа в условиях смешанного араукарно-южнобукового леса с бамбуком (Araucaria агаисапа - Nothofagus antarctica - Chusquea culeouY По мере старения растения независимо от условий произрастания форма сменяется на зонтиковидную. Араукария чилийская имеет только три порядка ветвления, включая ствол, при этом в сообществах выявлено 13 форм кроны.
Дальнейшие исследования вариативности форм кроны и реакции растения на среду позволят понять механизмы адаптивности дерева и помогут в разработке неинвазивных методов определения возраста араукарии чилийской.
Список литературы Форма кроны Araucaria araucana (Molina) K.Koch в разных возрастных состояниях и экологических условиях естественных местообитаний
- Антонова И.С., Фатьянова Е.В. 2013. К вопросу о строении ветвей деревьев умеренной зоны в контексте онтогенетических состояний//Вестн. ТвГУ. Сер.: Биология и Экология. Вып. 32. № 31. С. 7-24.
- Антонова И.С., Фатьянова Е.В., Зайцева Ю.В. 2012. Мультимасштабность побеговых систем некоторых деревьев умеренной зоны (разнообразие, классификация, терминология)//Актуальные проблемы современной биоморфологии/под ред. Н.П. Савиных. Киров. С. 390-402.
- Восточноевропейские широколиственные леса. 1994/Р.В. Попадюк, А.А. Чистякова, С.И. Чумаченко, О.И. Евстигнеев, Л.Б. Заугольнова, И.И. Истомина, Л.Л. Киселева, С.А, Костяев, О.В. Смирнова, Н.А. Торопова, А.В. Французов, Л.Г. Ханина, Н.И. Чернов, Т.О Вяницкая./ред. О.В. Смирнова. М.: Наука. 364 с. Ипатов В.С. 1990.
- Отражение динамики растительного покрова в синтаксономических единицах.//Бот. журн. Т. 75. № 10. С. 1380-1388.
- Казакова Н.Л., Антонова И.С. 2014. К характеристике развития проростков и имматурных растений Araucaria araucana (Molina) K. Koch (Araucariaceae) в естественных местообитаниях (Аргентина, штат Неукин)//Modern Phytomorphology. № 5. С. 193-207.
- Методы изучения лесных сообществ. 2002/Е.Н. Андреева, И.Ю. Баккал, В.В. Горшков, И.В. Лянгузова, Е.А. Мазная, В.Ю. Нешатаев, В.Ю. Нешатаева, Н.И. Ставрова, В.Т. Ярмишко, М.А. Ярмишко. СПб.: НИИХимии СПбГУ. 240 с.
- Муравьева О.А., Борхвардт В.С., Жилин С.Г. 1974. Семейство араукариевые (Araucariaceae)//Жизнь растений: в 6-ти томах/ред. А.Л. Тахтаджян. М.: Просвещение. Т. 4. С. 335-343.
- Смирнова О.В., Чистякова А.А., Истомина И.И. 1984. Вазисинильность как одно из проявлений фитоценотической толерантности растений//Журн. общ. биол. Т.45. №2. С. 216-225.
- Burns B. 1991. The regeneration dynamics of Araucaria araucana. PhD Thesis. Univ. of Colorado. 211 p.
- Burrows G.E. 1987. Leaf axil anatomy in the Araucariaceae//Australian Journal of Botany. № 35. P. 631-640.
- Burrows G.E. 1990. The role of axillary meristems in coppice end epicormic bud initiation in Araucaria cunninghamii//Botanical Gazette. № 151. P. 293-301.
- Burrows G.E. 1999. Wollemi pine (Wollemia nobilis, Araucariaceae) possesses the same unusual leaf axil anatomy as other investigated members of the family//Australian Journal of Botany 47. P.61-68.
- Cavieres A. 1987. Estudio de crecimiento de Araucaria araucana (Mol.) Koch, en un bosque virgen de Araucaria-Lenga//Tesis para optar al. título de Ing. Forestal. Univ. de Chile. 98 p.
- Donoso C. 1981. Ecología Forestal. El: Bosque y su medio ambiente. Editorial Univ. Santiago. 369 p.
- Donoso C.1997. Árboles nativos de Chile. Guía de reconocimiento. Octava edición. Marisa Cúneo ediciones. Valdivia. 116 p.
- Donoso, C. 1993. Bosques templados de Chile y Argentina. Variación, estructura y dinámica. Ecología Forestal. Editorial Univ. Santiago. 484 p.
- Donoso C. 2006. Las species arboreas de los Bosque Templados de Chile y Argentina. Autoecologia. Valdivia Chile: Marisa Cuneo Ediciones. 678 p.
- Drake F.A., Herrera M.A., Acuña E.C. 2005. Propuesta de manejo sustentable de Araucaria araucana (Mol.) Koch//Bosque. No. 26(1). P. 23-32.
- Galardo R. 1980. Vegetación del bosque de Araucaria araucana (Mol.) Koch en la Cordillera de los Andes (Lonquimay, provincia de Malleco). En Boletín Técnico. № 57. Univ. de Chile. 25 p.
- Grosfeld J. 1994. Arquitectura y dinámica de crecimiento de Araucaria araucana (Mol.) K.Koch (Araucariaceae)//Tesís de Licenciatura Universidad Nacional del Comahue. Centro Regional Univ. Bariloche.
- Grosfeld J., Barthélémy D., Brion C. 1999. Architectural variation of Araucaria araucana (Molina) K. Koch (Araucariaceae) in its natural habitat//The evolution of plant architecture/eds. M.H. Kurman, A.R. Hemsley. London: Royal Botanic Gardens Kew. P. 109-122.
- Hallé F., Oldeman R.A.A., Tomlinson P.B. 1978. Tropical trees and forests an architectural analisis. Berlin: Springer Verlag. 441 p.
- Hoffmann A.E. 1991. Flora silvestre de Chile. Zona araucana. 2 ed. Santiago: Ediciones Fundación Claudio Gay. 258 p.
- Tomlinson P.B. 2002. Crown structure in Araucariaceae. Kalaheo: Harvard Forest, Petersham MA and National Tropical Botanical Garden. P. 55-65.
- Marticorena C.Y., Rodrígues R. 1995. Flora de Chile. (Pterydophyta-Gymnospermae). Vol. I. Concepción: Editorial Aníbal Pinto. Univ. Concepción. 352 p.
- Lusk C.H., Le-Quesne C. 2000. Branch whorls of juvenile Araucaria araucana (Molina) Koch: are they formed annually?//Revista Chilena de Historia Natural. № 73. Р. 497-501.
- Schmidt H., Toral, M., Burgos P. 1980. Aspectos de estructura y de regeneración natural para el manejo silvícola de los bosques de Araucaria-Lenga//Forestry problems of genus Araucaria. Brazil: IUFRO meeting held in Curitiba. P. 159-166.
- Sanguinetty J., Kitzberger T. 2008.Petterns and mechanisms of masting in the large-seeded southern hemisphere conifer Araucaria araucana//Austral Ecology. № 33. P. 78-87.
- Veblen T. 1982. Regeneration patterns in Araucaria araucana forests in Chile//Journal of Biogeography. № 9. P. 11-28.
- Veblen T., Burns T., Kitzberger A., Lara R. 1995. The ecology of conifers of Southern South America//Ecology of the southern conifers/Eds N.J. Enright, R.S. Hill. Victoria Carlton: Melborne Univ. Press. P. 120-155.
- Veillon J.M. 1978. Architecture of the New Caledonian species of Araucaria//Tropical trees as living systems/Eds Tomlinson, P.B. Zimmermann, H.M. Cambridge: Cambridge University Press. P. 233-245.
- Veillon J.M. 1980. Architecture des espéces néocalédoniennes du genre Araucaria//Candollea. № 35. Р. 609-640.
- Vita A. 1996. Los tratamientos silviculturales. 2 ed. Santiago: Univ. de Chile. 156 p.
