Формирование кластеров в процессе колонизации ареала двухвозрастной популяции

Автор: Кулаков М.П., Фрисман Е.Я.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Биология. Математическое моделирование
Статья в выпуске: 4 т.20, 2017 года.
Бесплатный доступ
Изучается модель пространственно-временной динамики популяций с возрастной структурой на одномерном ареале. Показано, что тип динамики определяется демографическими и миграционными параметрами, а также начальными численностями особей в разных частях ареала. Выявлено, что в ходе колонизации изначально пустого ареала основной тип динамики - это кластерная синхронизация, перемешанная с химерами, которая при определенных параметрах оказывается устойчивой и реализуется продолжительное время либо происходит быстрая десинхронизация и формируются некогерентные режимы.
Популяция, возрастная структура, миграция, пространственно-временная динамика, синхронизация
Короткий адрес: https://sciup.org/143161702
IDR: 143161702 | УДК: 574.34,
Formation of clusters at colonization of the area by the two-aged population
We study the model of spatial-temporal dynamics of populations with age structure on a one-dimensional area. We show that the type of dynamics is determined by demographic and migration parameters, as well as by the initial numbers of individuals in different parts of the area. It is shown that during colonization of the initially empty area, the main type dynamics is cluster synchronization including chimeras. Under certain parameters it demonstrates stability and can exist for a long time. Or, there is a rapid desynchronization, and incoherent modes arise.
Текст научной статьи Формирование кластеров в процессе колонизации ареала двухвозрастной популяции
Как известно, многие популяции характеризуются неравномерным или мозаичным распределением по ареалу, и существует множество гипотез такого распределения. Часто его связывают с географическими особенностями ареала. Например, естественные барьеры типа гор, рек и т.п. определяют то, куда особи популяции в принципе способны распространиться, т.е. предполагается, что особи достаточно подвижны и за непродолжительное время (за небольшое число поколений) способны перераспределиться по ареалу [5, 6]. Кроме того, пространственная приуроченность популяции к тому или иному местообитанию может быть связана с распределением по территории пищевого ресурса, необходимого для жизнедеятельности. В результате особи в большом числе локализуются на территориях с богатыми ресурсами, а на бедных территориях численность оказывается минимальной. С другой стороны, существует гипотеза о «внутренних» механизмах формирования неоднородного распределения, связанного с конкурентными взаимодействиями особей между собой или другими видами. Например, особи разных стадий развития могут отличаться своей подвижностью, конкурировать между собой или представителями другого вида. В результате, если существует несколько местообитаний, то особи мигрируют между ними вследствие этих взаимодействий или особенностей жизненного цикла, т.е. происходит вытеснение представителя той или иной стадии развития на другую территорию из-за конкурентных взаимоотношений. Взаимодействие же с другими видами может выражаться, например, в том, что происходит миграция хищника за жертвой (таксис) [3, 16].
Одним из методов изучения неравномерного распределения является математическое моделирование. В данной работе исследуется модель пространственной динамики структурированных популяций с дискретным временем и пространством, которая представляет собой систему или решетку связанных двумерных отображений (двух рекуррентных уравнений) [15]. Рассматривается популяция с двумя возрастными классами – младшим и старшим. Предполагается, что существует несколько местообитаний с проживающими там локальными популяциями с возрастной структурой, между которыми наблюдаются сезонные миграции особей старшего поколения [9, 10].
В рамках данной модели рассматриваются механизмы синхронизации (в том числе кластерной) и формирования неоднородного распределения в процессе расселения или колонизации одномерного линейного ареала (кольца) из единственной субпопуляции с ненулевой численностью. Подобная форма ареала возникает для популяций животных, у которых ареал вытянут вдоль какого-либо природного объекта, например, русла реки, долинного комплекса, берега острова или озера, склона горы, границы между лесом и лугом [2, 4, 7]. Заселение ареала из единственной локальной популяции наблюдается, например, в случае катастрофического уничтожения популяции почти на всем ареале и последующего его восстановления из единственной выжившей популяции. Кроме того, такая ситуация характерна для новых и пионерских для данной территории видов, заселяющих территорию [11].
Вместе с тем вопросы расселения и формирования неравномерного распределения в данной модели тесно связаны с вопросами пространственной синхронизации и десинхронизации, т.е. тем, как особенности динамики популяции на удаленных территориях связаны между собой, и тем, как это влияет на формируемые пространственные структуры. Данный вопрос, в частности, предполагает, что динамика популяции носит колебательный характер, т.е. численность не постоянна и периодически или апериодически изменяется во времени, или сложным образом флуктуирует. В этой связи интересны разнообразные нелинейные феномены, характерные для связанных колебательных элементов. Это, например, такие явления, как синхронизация, в том числе хаотическая и кластерная, пространственно-временной хаос, пространственно-временная перемежаемость, химеры и др. [1, 12–14]. Данная работа посвящена исследованию таких явлений в структурированных популяциях методами математического моделирования и теории синхронизации.
Модель пространственной динамики популяции с возрастной структурой В работе исследуется следующая математическая модель с дискретным временем и пространством, предложенная в [10]:
( i ) ( i ) ( i ) ( i )
x n + 1 = a y n exp — X^n — y n )
i + P
(i) (i) (i) m v (j) (i) , yn +1 s xn + v n + ^\yn yn )
-
2 P j = i - P
( i = 1,2, _ , N ). (1) где xn ( i ) и yn ( i ) – численности младшего и старшего возрастных классов в i -м локальном участке в n -й сезон, a – репродуктивный потенциал или максимальный коэффициент рождаемости, s и v – коэффициенты выживаемости соответствующих возрастных групп, параметр N определяет степень участия младшего возрастного класса в плотностно-зависимой регуляции рождаемости [9, 10]. Рассматривался ареал в форме одномерного кольца, а это значит, что имеется N связанных местообитаний с проживающими там субпопуляциями.
Связь в системе (1) представляет собой нелокальную миграцию старших особей, а это означает, что доля взрослых, равная величине m, один раз в сезон покидает свое родное местообитание и мигрирует не только на соседние участки, но и на P удаленных от него территорий, соответственно слева и справа от его родного участка. Если P =1, то связь локальная и связанными оказываются только соседние субпопуляции. Например, первая связана со 2-й и с N- й. Если P= 2, то первая популяция связана со 2-й и 3-й, а также N- й и N- 1-й, и т.д. В этом смысле расстояние между субпопуляциями определяется их расположением на кольцевом ареале, т.е. номером i , а также радиусом связи, равным величине P . Такое модельное описание миграции предполагает, что особи явно отличаются своей мобильностью (при P> 1), т.е. одна их часть способна мигрировать лишь локально (в соседние местообитания с номерами i ± 1 ), а другая часть перемещается на более отдаленные территории (с номерами i ± 2 , i ± 3 и т.д., но не далее чем i ± P , где P < N /2).
Надо заметить, что в отсутствии связи (т.е. m = 0) каждая локальная популяция, которая описывается этой моделью, демонстрирует весьма сложное поведение [9]. Например, в зависимости от соотношений демографических параметров системы (1) возможна потеря устойчивости по двум сценариям. Во-первых, через образование предельной инвариантной кривой, во-вторых, через удвоение периода, и, соответственно, при определенных популяционных параметрах формируются квазипериодические, а при других хаотические режимы динамики численности. Кроме того, в этом случае наблюдается мультистабильность, т.е. в зависимости от начальных численностей взрослых y 0 и детей x 0 формируются принципиально разные динамические режимы, например, колебания численностей с разными периодами. В этой связи особый интерес вызывает то, к чему приводит миграционная связь между популяциями со столь сложным динамическим поведением.
Синхронизация, кластеризация, химеры в ходе колонизации
Основные феномены, которые возникают в модели (1), связаны с явлением синхронизации динамики соседних и удаленных субпопуляций [8–10]. Если все субпопуляции оказываются синхронными, т.е. их динамика максимально близка друг к другу (совпадают периоды, близки фазы и амплитуды колебаний), то говорят о полной синхронизации. Если же можно выделить две или бо- лее группы синхронных элементов, то говорят о кластерной синхронизации или, просто, кластеризации. Кроме того, возможна ситуация, когда среди синхронных элементов могут начать появляться одиночные несинхронные элементы, которые выбиваются из общего ритма колебаний. В этом случае говорят о химерных состояниях или, для простоты, о химерах [13]. И, наконец, все элементы могут быть несинхронными друг другу, и тогда говорят о несинхронном или о некогерентном поведении. Кроме того, несинхронные элементы сами могут образовать кластер, о котором можно говорить, если одновременно с ним существует кластер с синхронной динамикой.
Рассмотрим особенности формирования неоднородного пространственного распределения по ареалу при следующих начальных условиях:
-
(1) (100) (1) (4 9) (51)
x 0 ”• x 0 y 0 ”• y 0 y 0
которые приводят к задаче заселения или колонизации изначально пустого ареала. Здесь существенно, что ареал имеет кольцевую форму и поэтому не столь важно, какую именно ненулевую субпопуляцию рассматривать.
В этом случае пространственно-временную динамику системы (1) при начальных условиях (2) удобно представить тремя способами, показанными на рис. 1, где у 050) = 1.6 . Выбранные популяционные параметры a , s , v и ρ в данном случае соответствуют устойчивым колебаниям периода 2 (2-цикл) локальной популяции.
На рис. 1, во-первых, приведены профили (снизу) асимптотических пространственно-временных режимов, на которых по оси абсцисс отложен номер субпопуляции i , а по оси ординат значения переменной y за достаточно большой период времени. В результате численность i -й субпопуляции в n -й сезон, равная у Пi ) ( i = 1,2, ^ , N ),
= y 0100) = 0, y 05 0) > 0, (2)
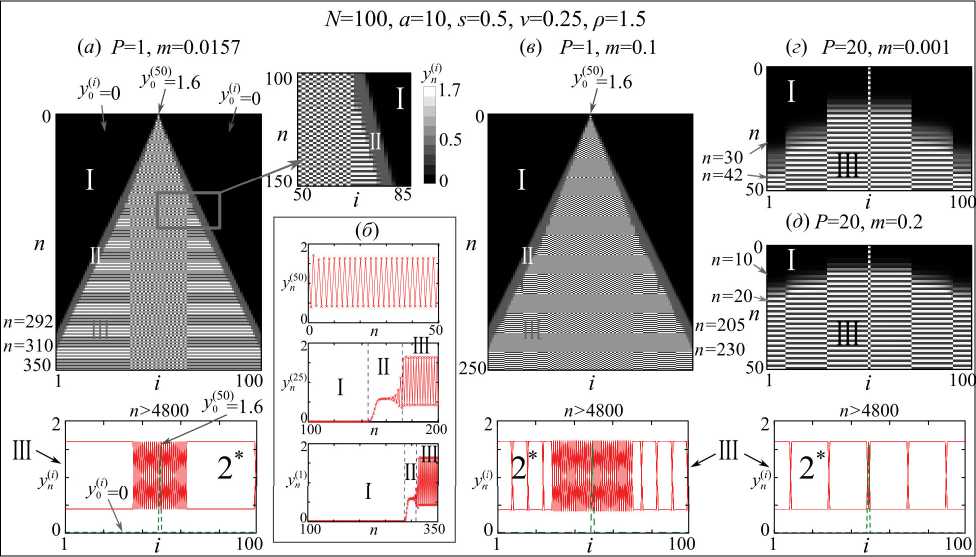
Рис. 1. Кластеры и химеры, возникающие в процессе колонизации кольцевого ареала, состоящего из 100 местообитаний, из единственной ненулевой субпопуляции с численностью y 0 (5 0) при различных параметрах связи, а также профили (снизу). I–III – этапы колонизации, число – регистрируемый период колебаний, 2* – химера
Fig. 1. Clusters and chimeras arising at colonization of the ring-shaped area, consisted of 100 habitats, of a single non-zero subpopulation with the number equal to y 0 (5 0) , at various couple parameters; and profiles, as well (below). I–III – the stages of colonization, figure – a detected period of fluctuation, 2* – chimera
соответствует одной точке на профиле, соединив которые, для всех N субпопуляций получаем одну из линий профилей для n -го сезона. Вычислив достаточное число итераций ( n = 0,1,2,..,5000) уравнений (1) и отбросив большую их часть от начала (оставив n = 4800,4801,..,5000), получаем несколько линий, изображенных на профилях, которые полностью характеризуют асимптотическую пространственную динамику. Например, часть этих линий сливается, что, с одной стороны, говорит о том, что имеет место точная повторяемость значений численностей через определенное число итераций (проверено численно с точностью 10-9), а, с другой, по их числу легко судить о периоде колебаний (с точностью 10-9), который отмечен для каждого профиля. Если такой повторяемости нет, то имеет место нерегулярная или хаотическая динамика во времени. Кроме того, видно, что некоторые линии пересекаются, а это значит, что когда численность субпопуляций в одной группе (кластере) достигает максимальных значений, другие субпопуляции демонстрируют минимальные значения, и наоборот, т.е. их динамика оказывается несинхронной (не совпадают фазы). Таким образом, по виду профилей легко судить о наличии кластеров синхронных субпопуляций, у которых полностью совпадает фаза и период колебаний.
Во-вторых, на рис. 1 (сверху) показана динамика субпопуляций во времени, где по оси абсцисс отложен номер субпопуляции i , по оси ординат дискретное время n сверху вниз, а значению численностей соответствует яркость точек на пересечении номера i и времени n . Черной точке соответствует нулевая численность субпопуляций, белой – максимальная, а серой – некоторые промежуточные значения (легенда показана на выноске в рис. 1 а ). Такое представление, в отличие от профилей асимптотических пространственно-временных режимов, позволяет проследить динамику с первых итераций модельных уравнений и в деталях изучить процесс колонизации.
Наконец, динамика отдельных субпопуляций на рис. 1б представлена в виде графиков численностей, т.е. показана зависимость y(i) от дис-n кретного времени n для фиксированных номеров местообитаний i.
Численные эксперименты позволяют заключить, что модельный процесс расселения состоит из трех этапов, обозначенных на рис. 1 латинскими цифрами I, II и III.
-
(I) На первом этапе происходит начальное заселение ареала из единственной непустой
субпопуляции с выбранным в соотношениях (2) номером i = 50 ( y 0 (50) > 0 ). На рис. 1 а это начальное состояние расположено на вершине полосатого треугольника, а справа на рис.1 б показана динамика численности именно этой субпопуляции, которая представлена колебаниями с периодом 2 практически с самых первых итераций модельных уравнений (рис. 1 б ). Благодаря миграции пустые местообитания постепенно заполняются мигрантами, и чем дальше местообитание расположено от начального, тем позже оно заполняется. Следует ожидать, что это произойдет ровно через N /2 сезонов. Однако этого не происходит, что связано с несколькими обстоятельствами.
Во-первых, помимо механического движения по типу диффузии (миграционный член в системе (1)), в популяции присутствует смертность особей старшего возрастного класса (член vy ), в том числе вновь прибывших, а также выживаемость неполовозрелых особей (член sx ). В результате прирост численности на колонизируемых территориях оказывается значительно ниже того, что имел бы место при простой диффузии и отсутствии смертности ( s= 1, v= 1). Таким образом, за N /2 сезонов «края» ареала никто не достигает (модельная численность меньше10–32). Во-вторых, модельные темпы воспроизводства на колонизируемых территориях оказываются значительно ниже темпов смертности и миграции на сопредельные территории, что, вероятно, связано с небольшими численностями мигрантов и/или с низким репродуктивным потенциалом. В результате, например, 24-ю субпопуляцию мигранты достигают примерно через 150 сезонов, а 1-ю лишь через 292 сезона (рис. 1 б ). В данном случае мы считаем, что местообитание колонизировано, если плотность населения там значительно больше нуля (больше 10–6). Графически этот этап на рис. 1 а изображен в виде двух черных треугольников, где численность субпопуляций примерно равна нулю (меньше 10–6).
-
(II) На втором этапе, после того как численность начинает превышать некоторое критическое значение (более 10–6) и можно считать, что в местообитании проживает достаточное число особей, происходит качественная смена типа динамики – от монотонного роста очень малых величин к ограниченному (логистическому) росту со скоростями порядка репродуктивного потенциала a . Этот рост указывает на то, что рождаемость на этом этапе полностью компенсирует смертность и эмиграцию и происходит активное заселение территории. Вместе с тем ограниченный рост
продолжается до тех пор, пока эмиграционный отток полностью компенсирует рост внутренней конкуренции, однако при достижении некоторых критических плотностей включаются плотностно-зависимые механизмы регуляции, описываемые экспонентой в системе (1), и динамика переходит к устойчивым колебаниям с периодом 2. Соответственно этот этап на рис. 1 а показан серой полосой, где динамика субпопуляций оказывается квазистационарной.
-
(III) На третьем этапе динамика всех субпопуляций становится колебательной и в ряде случаев наблюдается синхронизация. Вместе с тем популяции, близкие к начальной 50-й популяции, часто оказываются не синхронными друг другу, а удаленные могут образовывать кластеры синхронных элементов. Данный факт подтверждается профилями снизу, которые показывают асимптотическую динамику системы (1) через достаточно большое число сезонов ( n >4800).
Вместе с тем время начала второго этапа колонизации существенно определяется радиусом связи и коэффициентом миграции. Причем эта зависимость довольно нетривиальная. Например, увеличение коэффициента миграции m примерно в 6 раз ( m= 0,0157 на рис. 1 а и m= 0,1 на рис. 1 в ) приводит к тому, что полное заполнение ареала произойдет на 80 сезонов позже (230 сезонов на рис. 1 в против 310 на рис. 1 а ), а с ростом радиуса связи P в 20 раз ( P= 1 на рис. 1 в и P= 20 на рис. 1 г ), это произойдет примерно от 6 до 10 раз быстрее, в зависимости от коэффициента миграции (рис. 1 в и рис. 1 г - д ). Кроме того, с ростом радиуса связи второй этап расселения становится короче и практически не наблюдается для больших значений P .
По всей видимости, второй этап, в принципе, возможен лишь в случае сценария удвоения периода, а при сценарии Неймарка-Сакера он не наблюдается и после первого этапа расселения субпопуляции демонстрируют квазипериодиче-ские колебаний либо формируется один из резонансных или мультистабильных режимов. В этом смысле это обобщает ранее полученные результаты в исследованиях расселения в системе связанных популяций с непересекающимися поколениями, описываемых системой локально связанных одномерных отображений Рикера [8].
Серия других вычислительных экспериментов показала, что асимптотический пространственно-временной режим существенно зависит от параметров связи. В частности, при очень слабой связи, как в первых двух случаях на рис. 1, получить хоть какой-нибудь режим синхронизации очень проблематично. В этом случае преимущественно возникает множество небольших кластеров, сосуществующих с кластером несинхронных элементов, т.е. наблюдаются прежде всего фазовые химеры, а полная синхронизация, в том числе кластерная, возможна при довольно большой связи, как в последних двух примерах (рис. 2г-д).
Кроме того, начальная популяция с ненулевой численностью, из которой идет заселение ареала, часто становится неким центром, вокруг которого располагаются несинхронные субпопуляции, и профили на рис. 2 демонстрируют это утверждение. Здесь 50-я субпопуляция, в ряде случаев, оказывается несинхронной соседним, которые, в свою очередь, образуют кластеры довольно причудливых форм.
Помимо прочего, на рис. 2 показано, как усложняются пространственно-временные режимы при разных значениях параметров. Если в первом случае наблюдаются преимущественно синхронные колебания с периодом 4 (за исключением несинхронной начальной популяции и некоторых других), то по мере того как происходит каскад удвоения периода и формируется развитой хаотический режим, все субпопуляции оказываются несинхронными. Причем они окончательно теряют свою синхронность и формируется пространственно-временной хаос лишь после того, как ареал полностью заполнится. Например, в первом и втором случаях (рис. 2 а - б ) сразу после заполнения ареала формируется семь кластеров и один несинхронный элемент, представленный начальной субпопуляцией. Но только в первом случае кластеры оказываются устойчивыми и существуют продолжительное время (на время счета), в то время как во втором они быстро десинхронизируются и уже через 20 итераций имеют абсолютно несинхронное поведение.
Аналогичная ситуация наблюдается и в других примерах, где сразу после колонизации ареала наблюдается один несинхронный элемент и три кластера, расположенные примерно на тех же местах во всех шести случаях (рис. 2 в - з ). Однако в зависимости от параметров связи и начальной численности асимптотические пространственные режимы оказываются принципиально разными. Это, например, несинхронный режим на рис. 2 в , режим сосуществования кластеров с синхронной и несинхронной динамикой на рис. 2 г - е и, наконец, когерентная динамика с разным числом кластеров на рис. 2 ж - з .
Интересно, что разрушение кластеров, которые всегда образуются после полного заполне-
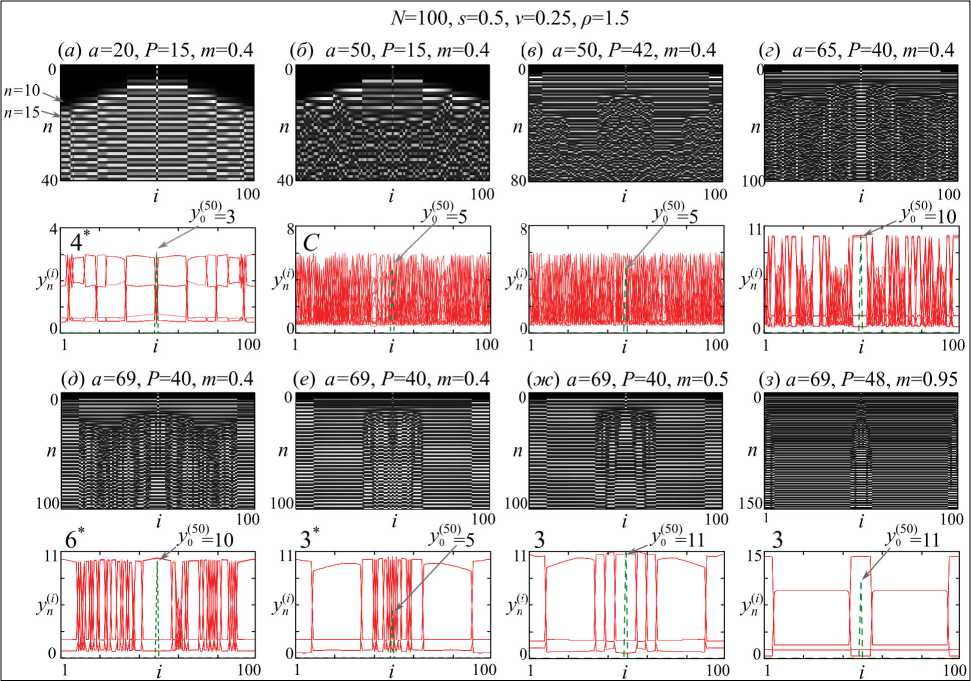
Рис. 2. Ход колонизации (сверху) и профили асимптотических пространственно-временных режимов (снизу) при разных популяционных и миграционных параметрах, а также начальных численностях. Число – регистрируемый период колебаний, C – хаотическая динамика, верхний индекс «*» – химера
Fig. 2. Process of colonization (at the top), and profiles of asymptotic spatial-temporal regimes (below), at different population and migration parameters and the initial population numbers Figure-a detected period of fluctuation, «*» – chimera ния ареала, происходит по-разному. Например, в последних трех примерах (рис. 2е-з) в окрестности начальной популяции, из которой происходит заселение, несинхронными оказываются довольно большое число субпопуляций. Однако в первом (рис. 2е) они остаются несинхронными навсегда, но в последних двух примерах они все же синхронизируются через довольно продолжительный период времени (более 100 и 150 сезонов соответственно) и образуются один (рис. 2ж) или несколько (рис. 2з) кластеров синхронных элементов.
Выводы
Таким образом, в работе обнаружено, что тип пространственно-временной динамики популяций с возрастной структурой существенно определяется управляющими параметрами, а так- же начальными численностями особей в разных частях ареала. Основные типы динамики связаны с когерентным или синхронным поведением связанных популяций, частным проявлением которого является кластерная синхронизация, а также химеры как переходное состояние между синхронным и несинхронным поведением. В этом смысле динамика структурированных популяций демонстрирует мультистабильность в самом широком смысле, которая проявляется в следующем: различные начальные численности приводят не только к кластерам, элементы которых отличаются лишь фазами колебаний («классическая» когерентная динамика), но также разными амплитудами и периодами в смежных кластерах («смешанная» кластерная синхронизация). Например, кластер с хаотической динамикой сосуществует с кластером, испытывающим регулярные колебания, либо кластер синхронных элементов реализуется одновременно с кластером некогерентных элементов. Кроме того, в рамках данной модели описан режим, известный как уединенные состояния [14], интересный тем, что на фоне слабых синхронных или несинхронных колебаний возникают одиночные некогерентные элементы с много большей амплитудой.
С использованием предложенной модели в работе изучен процесс расселения или колонизации изначально пустого ареала из единственной субпопуляции с ненулевой численностью. Показано, что это проходит в три этапа.
На первых двух происходит первичное заполнение ареала, которое сопровождается ограниченным ростом численностей в разных местообитаниях. Выявлено, что колонизация кольцевого ареала с N потенциальными местообитаниями и локальной связью (миграцией на смежные территории) невозможна за короткое время (всегда больше N /2). Кроме того, колонизация идет быстрее при больших коэффициентах миграции и радиусах связи.
На последнем этапе формируются устойчивые колебания, которые в ряде случаев оказываются синхронными, и образуются кластеры. Показано, что существует сложная связь между параметрами связи, начальной численностью и формируемым режимом. В частности, кластеры возможны при сильной связи (радиусе и силе), а при слабой связи возникают преимущественно несинхронные или «смешанные» режимы. Кроме того, оказалось, что после первичного заполнения ареала, до выхода на устойчивый режим, кластеры возникают практически всегда. Однако в зависимости от параметров связи они либо существуют продолжительное время, т.е. оказываются устойчивыми, либо десинхронизируются и формируются некогерентные режимы.
Список литературы Формирование кластеров в процессе колонизации ареала двухвозрастной популяции
- Богомолов С.А., Стрелкова Г.И., Schöll E., Анищенко В.С. Амплитудные и фазовые химеры в ансамбле хаотических осцилляторов//Письма в ЖТФ. 2016. Т. 42, вып. 14. С. 103-110
- Гилев А.В. Закономерности пространственного распределения и научные основы охраны рыжих лесных муравьев//Зоологический журнал. 2010. Т. 89, № 12. С. 1413-1420
- Говорухин В.Н., Моргулис А.Б., Тютюнов Ю.В. Медленный таксис в модели хищник-жертва//Доклады академии наук. 2000. Т. 372, № 6. 730-732
- Громов В.С. Пространственно-этологическая структура популяций грызунов. М.: Т-во науч. изданий КМК, 2008. 581 с
- Домбровский Ю.А. Пространственная структурированность и жизнеспособность популяций//Журнал общей биологии. 1985. Т. 46, № 2. С. 278-283
- Домбровский Ю.А., Тютюнов Ю.В. Структура ареала, подвижность особей и живучесть популяции//Журнал общей биологии. 1987. Т. 48, № 4. С. 493-498
- Коробченко М.А. Расширение ареала крота европейского (talpa europaea) в долине реки Северный Донец//Зоологический журнал. 2009. Т. 88, № 4. С. 465-472
- Кулаков М.П. Закономерности кластеризации динамики численности популяций, находящихся в процессе расселения особей по линейному ареалу//Региональные проблемы. 2015. Т. 18, № 4. С. 33-39
- Кулаков М.П., Неверова Г.П., Фрисман Е.Я. Мультистабильность в моделях динамики миграционно-связанных популяций с возрастной структурой//Нелинейная динамика. 2014. Т. 10, № 4. С. 407-425
- Кулаков М.П., Фрисман Е.Я. Кластеризация и химеры в пространственной динамике популяций с возрастной структурой на кольцевом ареале//Региональные проблемы. 2016. Т. 19, № 4. С. 5-11
- Лобков В.А. Причины и особенности динамики ареалов некоторых млекопитающих//Ученые записки Таврического национального университета. Серия «Биология, Химия». 2004. Т. 17, № 56. С. 3-11
- Павлов Е.А., Осипов Г.В. Синхронизация и хаос в сетях связанных отображений в приложении к моделированию сердечной динамики//Компьютерные исследования и моделирование. 2011. Т. 3, № 4. С. 439-453
- Семенова Н.И., Анищенко В.С. Переход «когерентность -некогерентность» с образованием химерных состояний в одномерном ансамбле//Нелинейная динамика. 2016. Т. 12, № 3. С. 295-309
- Шепелев И.А., Вадивасова T.Е. Уединенные состояния в 2D-решетке бистабильных элементов при глобальном и близком к глобальному характере взаимодействия//Нелинейная динамика. 2017. Т. 13, № 3. С. 317-329
- Castro, M.L., Silva, J.A.L, Justo D.A.R., Stability in an age-structured metapopulation model//J. Math. Biol. 2006. Vol. 52. P. 183-208
- Tyutyunov Yu.V., Titova L.I., Senina I.N. Prey-taxis destabilizes homogeneous stationary state in spatial Gause-Kolmogorov-type model for predator-prey system//Ecological Complexity. 2017. Vol. 31. P. 170-180
