Формирование напочвенного покрова в зависимости от глубины залегания кристаллических пород в скальных лесных сообществах Водлозерского национального парка (Архангельская обл.)
")
Автор: Тарасова Виктория Николаевна, Сонина Анжелла Валерьевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
Обосновывается выделение стадий формирования напочвенного покрова от эпилитных лишайников до сосудистых растений в зависимости от мощности почвенного покрова в условиях скальных растительных сообществ.
Архангельская область, скальные лесные сообщества, эпилитные, эпигейные лишайники, мхи, печеночники, сосудистые растения, формирование напочвенного покрова, глубина залегания кристаллических пород
Короткий адрес: https://sciup.org/148200830
IDR: 148200830 | УДК: 581.5
Formation of soil cover depending on occurrence depth of crystalline rocks in the rocky forest communities of Vodlozersky national park (Arkhangelsk region)
Justified the selection of stages in the formation of ground cover from the epilithic lichens to higher plants depending on the capacity of the soil cover in rocky plant communities.
Текст научной статьи Формирование напочвенного покрова в зависимости от глубины залегания кристаллических пород в скальных лесных сообществах Водлозерского национального парка (Архангельская обл.)
Cкальные лесные сообщества типичные для Фенноскандии и уникальные для таежной зоны в условиях севера проходят длительный этап становления. Прежде всего это связано с продолжительностью формирования почвенного покрова, которая зависит от природно-климатических условий: климата, рельефа, почвообразующих пород, растительности и др. [2]. Изменение поверхности скального субстрата и накопление органогенного слоя на начальных этапах почвообразования происходит в результате воздействия биотических факторов – действия микроскопических грибов, водорослей, акти-номицентов, эпилитных лишайников. На примитивной почве в зависимости от ее характерис-тик формируется растительный покров разного состава и структуры [4]. Изучение динамики его формирования в разных типах скальных фитоценозов в связи с развитием слоя почвы является основополагающим для понимания становления сообщества и разработки мер рационального использования этих природных комплексов. Актуальность такого рода исследований связана и с уязвимостью скальны х типов леса в результате различного антропогенного воздействия – пожаров, промышленной эксплуатации, рекреации [6]. Цель настоящей работы – выделение и обоснование стадий в формировании напочвенного покрова в скальных лесных сообществах в пределах геоботанического про-филя.
МАТЕРИАЛЫ И МЕТОДЫ
Работа выполнена на территории Архангельской области, в северо-западной части Национального парка «Водлозерский». Исследование проводилось на северо-восточном склоне возвышенности Му-ройгоры (максимальная высота около 200 м), представляющей собой выход на поверхность твердых кристаллических пород – диабазов, в виде обломочных глыб разного размера. Общая протяженность скального выхода составляет около 1,5 км, ширина – 150-200 м. В его пределах формируется
комплекс растительных сообществ, включающий в себя ельники, сосняки, а также смешанные еловососновые леса, нередко с примесью лиственных пород. По склону горы был заложен геоботанический профиль шириной 20 м и длиной 120 м, на котором на 7 пробных площадях размером 20х10 м выполнены полные геоботанические описания, включающие определение общих характеристик сообществ, характеристик деревьев и напочвенного покрова [5].
Описание напочвенного покрова выполнено на площадках размером 1х1 м, на которых отмечали видовое разнообразие и проективное покрытие видов мохово-лишайникового и травяно-кустарничкового ярусов. На каждой площадке измеряли относительную освещенность (люксметром), сквозис-тость (сквозистомером Ипатова), глубину залегания кристаллической породы (в 5 точках площадки при помощи металлического щупа). Отдельно проводили описание эпилитного лишайникового покрова с помощью рамки 10х20 см, в поле описания которой отмечалось число видов лишайников, общее покрытие, покрытие отдельных видов и характеристики местообитания: тип субстрата, свойства субстрата, угол наклона поверхности, экспозицию к сторонам света. Всего в пределах профиля выполнено 140 описаний напочвенного покрова, 214 описаний эпилитного лишайникового покрова. Собрано около 200 образцов лишайников, мхов и сосудистых растений. Определение лишайников выполнено в лабораторных условиях на кафедре ботаники и физиологии растений ПетрГУ, в гербарии Хельсинского университета, в лаборатории лихенологии и бриологии БИН РАН, мхов – в Институте биологии КНЦ РАН, печеночников – в ПАБСИ. Коллекции хранятся в гербарии ПетрГУ (PZV). Обработка данных выполнена на основе регрессионного анализа [4].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Вершина Муройгоры покрыта четвертичными отложениями, на которых развивается ельник скальный черничный зеленомошный с участием ели, сосны и березы. Ниже по склону горы мощность
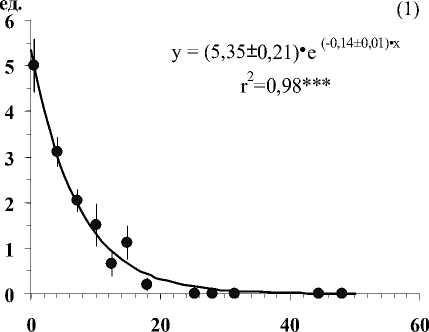
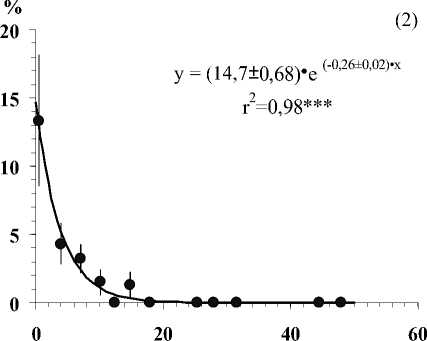
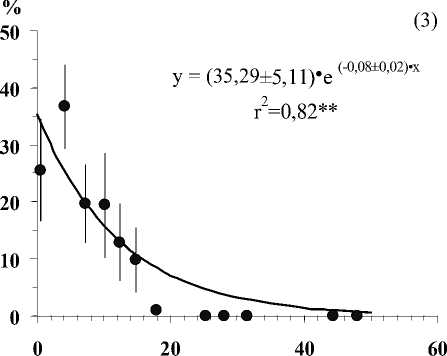
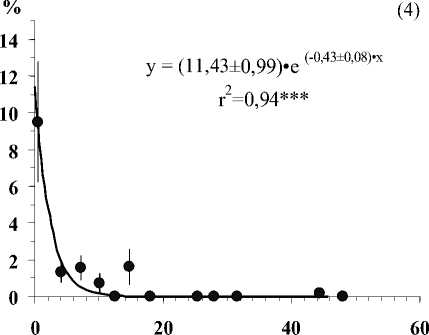
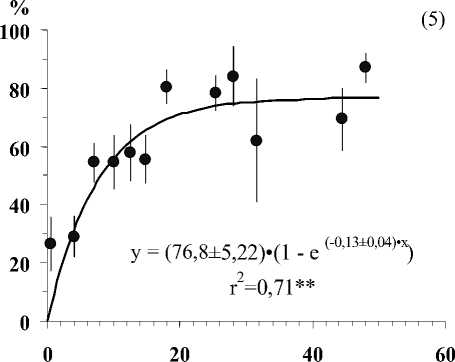
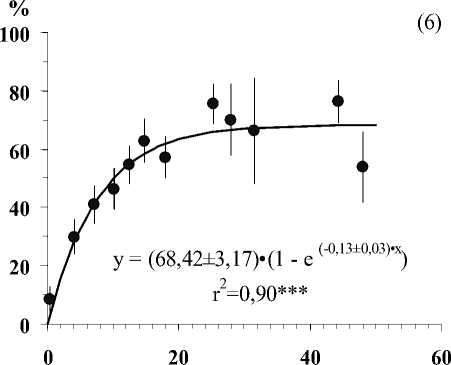
Глубина залегания кристаллических пород, см
Рис . Средние значения числа видов лишайников (1), проективного покрытия Cladonia amaurocraea (2), Clado-nia rangiferina (3), общего покрытия печеночных мхов (4), общего покрытия мхов (5) и общего покрытия травяно-кустарничкового яруса (6) при разной глубине залегания кристаллических пород в скальных сообществах. Примечание. Вертикальными линиями показана ошибка среднего. Линии на графиках проведены в соответствии с уравнениями регрессии. Коэффициенты регрессионных уравнений и корреляции отличны от нуля при уровнях значимости, соответственно 0,01(**) и 0,001 (***).
мощность четвертичных отложений уменьшается, доля участия ели в древостое постепенно снижается и ельник сменяется сосняком скальным брусничным лишайниково-зеленомошным. Середину склона покрывают сосняки скальные лишайниковые, отли- чающиеся достаточно разреженным древостоем из сосны. При переходе от середины к подножью склона мощность четвертичных отложений постепенно нарастает и сосняк лишайниковый последовательно сменяется смешанным елово-сосновым сообщест- вом, затем ельником черничным зеленомошным. Давность последнего пожара в изученных сообществах, определенная на основе анализа кернов, составляет 173 года. Возраст древостоя варьирует от 342 до 69 лет; преобладают деревья возрастом 120150 лет. Таким образом, растительный комплекс Муройгоры представляет собой сочетание различных сообществ на основе мощности почвенного слоя.
Примитивные почвы в северо-западном регионе можно разделить на четыре типа: корковые – формирующиеся на первых стадиях почвообразования, когда с появлением лишайников происходит накопление органического вещества и ускоряются процессы выветривания кристаллических пород; органогенные – характерной особенностью которых является образование лесной подстилки, которая залегает на скальном основании; щебнистые – почвы, в профиле которых появляется минеральный щебнистый горизонт с низким содержанием мелкозема; слабодифференцированные – являющиеся переходной стадией к неполноразвитым подзолам [6]. В пределах изученного профиля все эти стадии представлены в разном количественном соотношении в зависимости от типа растительного сообщества. Проанализировано формирование напочвенного покрова от эпилитных лишайников до развитых мохово-лишайникового и травяно-кустарничкового ярусов. Анализ, таким образом, включает эпилитные, эпигейные лишайники, печеночники, мхи и сосудистые растения.
В пределах обследованной площади (0,24 га) выявлено 35 видов эпилитных лишайников, среди которых преобладают накипные и листоватые биоморфы. Это указывает на длительность существования эпилитного покрова и наличие сукцессионных смен в направлении от накипных к листоватым и кустистым жизненным формам в связи с изменениями состояния скальной поверхности. По значениям покрытия и частоте встречаемости в описаниях выделяется два типа лихеносинузий. Первая – синузия с доминированием накипного вида Rhizo-carpon hochstetteri (Kцrb.) Vain., в которую входят в качестве содоминантов накипные виды родов Por-pidia и Rhizocarpon , вторая – синузия листоватого лишайника Arctoparmelia centrifuga (L.) Hale (табл.) с существенной долей участия накипных видов и включением листоватых, таких как Melanelia stygia (L.) Essl., M. hepatizon (Ach.) A. Thell.
В мохово-лишайниковом и травяно-кустарничковом ярусах встречаются 9 видов сосудистых растений, доминантами из которых являются Vac-cinium vitis-idaea L., V. myrtillus L., Ledum palustre L. Empetrum nigrum L. Моховый покров представлен 31 видом, среди которых 17 – мхи, 14 – печеночники. Среди мхов доминируют – Pleurozium schreberi (Brid.) Mitt. , Hylocomium splendens (Hedw.) Bruch et al., Polytrichum uniperinum Hedw., а также виды родов Schagnum ( S. russowii Warnst., S. girgensohnii Russov.) и Dicranum ( D. flexicaule Brid. , D. fuscescens
Turner); среди печеночных – Ptilidium ciliare (L.) Hampe, Neoorthocaulis floerkei (F.Weber et D.Mohr) L. Sцderstr., De Roo et Hedd., Barbilophozia hatcheri (A. Evans) Loeske, Tetralophozia setiformis (Ehrh.) Schljakov. Лишайниковый покров образуют 22 вида с преобладанием Cladonia amaurocraea Flцrke. Schaer ., Cladonia arbuscula (Wallr.) Flot., Cladonia rangiferina (L.) Weber ex F. H. Wigg., Cladonia stellaris (Opiz) Pouzar & Vezda.
В результате исследования установлено, что показатели видового разнообразия и обилия видов травяно-кустарничкового и мохово-лишайникового ярусов тесно связаны с глубиной залегания кристаллических пород (Рис.). По доминантным видом и ценотическим особенностям отдельных видов в напочвенном покрове выделяются 5 стадий с выраженными характерными чертами (см. табл.).
-
• Первая стадия – эпилитные лишайники на голом скальном субстрате.
-
• Вторая стадия выделяется на маломощном, до 1 см органогенном слое, где доминируют лишайники рода Cladonia с рано исчезающими горизонтальными талломами ( C. rangiferina, C. amauro-craeа) , а также виды рода Cladonia с развитыми горизонтальными талломами. Последние не образуют большого покрытия, но вносят существенный вклад в биоразнообразие яруса (см. рис.). Характерной чертой является включение в сообщество печеночников, с наибольшим для них разнообразием и обилием. В силу низкой конкурентоспособности они осваивают менее пригодные для мхов участки, а их способность образовывать различные биоморфы позволяет им расти между «веточками» в куртинах лишайников или формировать рыхлые ковры из плотно прикрепленных к субстрату побегов [1].
-
• Третья стадия – мощность почвенного покрова не превышает 7 см. Здесь хорошо развит мохово-лишайниковый ярус с преобладанием кустистых сильно разветвленных лишайников с рано исчезающими горизонтальными талломами ( Cladonia rangiferina, С. stellaris ). Возрастает роль мхов, на долю которых приходится около половины общего покрытия яруса, которые вытесняют печеночники и лишайники с горизонтально развитыми талломами. В сообщество включаются сосудистые растения.
-
• Четвертая стадия – мощность почвенного покрова составляет от 7 до 20 см. В моховолишайниковом ярусе преобладают мхи, с небольшим участием встречается Cladonia rangiferina . Здесь хорошо развит травяно-кустарничковый ярус, доминирует Vaccinium myrtillis ; отмечается максимальное значение покрытий Vaccinium vitis-idaea, Ledum palustre, Empetrum nigrum.
Пятая стадия – мощность почвы возрастает до 20–50 см. Напочвенный покров сложен в основном мхом Hylocomium splendens и кустарничком Vaccin-ium myrtillis с его абсолютным доминиро-ванием (см. табл.). Более развитый слой почвы позволяет чернике формировать довольно мощные подземные
Таблица . Стадии формирования напочвенного покрова в зависимости от глубины залегания кристаллических пород
Таким образом, выделенные стадии формирования напочвенного покрова согласуются с глубиной залегания кристаллических пород, или мощностью развития почвенного слоя. Установлено, что в процессе образования почвенного горизонта в отсутствие внешних нарушений происходит последовательная смена доминантных видов напочвенного покрова до момента наступления стационарного состояния. В наших условиях это состояние наступает при мощности почвенного слоя более 20 см и характеризуется стабильным участием в напочвенном покрове двух доминантных видов Hylocomium splendens и Vaccinium myrtillis.
Выражаем глубокую благодарность к.б.н. Е.А. Боровичеву (ПАБСИ) и к.б.н. М.А. Бойчук (Инсти- тут биологии КНЦ РАН) за помощь в определении видов печеночников и мхов.
Список литературы Формирование напочвенного покрова в зависимости от глубины залегания кристаллических пород в скальных лесных сообществах Водлозерского национального парка (Архангельская обл.)
- Боровичев Е.А. Формы роста печеночников (Marchantiophyta) Лапландсокго заповедника (Мурманская область)//Вестник МГТУ. 2011. Т. 14. № 3. С. 583-586.
- Дюкова Г.Р. Экология почвообразования, динамика и основные свойства почв Пензенской области//Известия ПГПУ. Естественные науки. № 1 (5). 2006. С. 50-57.
- Ивантер Э. В., Коросов А. В. Ведение в количественную биологию. Петрозаводск: Изд. ПетрГУ, 2003. 304 с.
- Кравченко А.В., Тимофеева В.В. О связи состава и структуры нижних ярусов скальных сосновых лесов и подстилающих коренных пород//Разнообразие почв и биоразнообразие в лесных экосистемах средней тайги. М.: Наука, 2006. C. 52-67.
- Методы изучения лесных сообществ. СПб.: НИИХимии СПбГУ, 2002. 240 с.
- Основы ландшафтной экологии европейских таежных лесов России. Петрозаводск: КарНЦ РАН, 2008. 227 с.
