Формирование устойчивости к гипотермии сортов Fragaria x ananassa Duch. разного эколого-географического происхождения в осенний период

Автор: Прудников П.С., Ожерельева Зоя Евгеньевна, Кривушина Диана Александровна, Зубкова Марина Ивановна
Журнал: Овощи России @vegetables
Рубрика: Агрохимия
Статья в выпуске: 6 (50), 2019 года.
Бесплатный доступ
Актуальность. Одним из значительных факторов, влияющих на рост и развитие земляники садовой, является отрицательная температура зимой и, особенно в ее бесснежный период. Представляло интерес изучить особенности окислительного стресса, накопления протекторных соединений, антиоксидантного и водного статуса в осенний период у сортов земляники садовой разного эколого-географического происхождения в условиях Орловской области. Материал и методы. Объектами исследований в осенний период служили листья сортов Fragaria ananassa различного эколого-географического происхождения: Царица, Alba, Sara, Korona, Кокинская ранняя, Соловушка, Урожайная ЦГЛ. Результаты. В осенний период при подготовке к зиме у сортов земляники Соловушка, Царица, Sara и Korona отмечали повышенную антиоксидантная активность (на примере ферментов СОД и каталазы), незначительное образование активных форм кислорода и, как следствие, низкое структурно-функциональное повреждение клеточных мембран. Также показано, что по мере снижения температуры происходило увеличение связанной и снижение свободной воды в листьях растений...
Земляника садовая, водный режим, пролин, перекисное окисление липидов, перекись водорода, каталаза, устойчивость, гипотермия
Короткий адрес: https://sciup.org/140245822
IDR: 140245822 | УДК: 634.75:631.524.85 | DOI: 10.18619/2072-9146-2019-6-80-83
Formation of resistance to hypothermia varieties Fragaria x ananassa Duch, different ecological-geographical origin in autumn period
Relevance. One of the most important factors which can influence on the growth and development of strawberries is the negative temperature in winter and especially in its snowless period. We were interested to study special aspects of the oxidative stress, accumulation of protector’s conjunctions, antioxidant and water status in autumn period of varieties strawberry garden different ecological-geographical origin in the conditions of the Orel region. Methods. The objects of the research were the leaves of Fragaria ananassa of different ecological and geographical origin in autumn period: Tsaritsa, Alba, Sara, Korona, Kokinskaya Rannya, Solovushka, Urozhainaya TsGL. Results. As the result was shown that in autumn period, when plants began preprocessing for winter, strawberry varieties Solovushka, Tsaritsa, Sara and Korona have been marked by increased antioxidant activity (for example, the enzymes SOD and catalase), little formation of reactive oxygen species and, in the end, low structural and functional damage to cell membranes...
Текст научной статьи Формирование устойчивости к гипотермии сортов Fragaria x ananassa Duch. разного эколого-географического происхождения в осенний период
Земляника садовая (Fragaria x ananassa Duch.) – одна из самых популярных и распространенных ягодных культур в мире. Свое широкое распространение земляника получила благодаря ее бес- спорным преимуществам в сравнении с другими ягодными культурами: скороплодность, высокие вкусовые качества, привлекательный вид и красивая окраска, богатый биохимический состав, лечебные свойства [1]. Расширение сортимента земляники возможно благодаря селекции и интродукции сортов из разных стран. Поскольку в России большинство насаждений садовых культур расположено в зоне рискованного земледелия, то важным условием успешного возделывания земляники является сочетание продуктивности сорта и его зимостойкости. Как правило, наиболее урожайные и крупноплодные сорта земляники имеют низкую зимостойкость, которая влияет не только на продуктивность, но и на жизнеспособность растений в целом. В связи с этим для Центрального региона РФ важным свойством, которым должен обладать сорт земляники является устойчивость к низким температурам и среднесуточным перепадам температур [2].
Устойчивое состояние растений земляники к гипотермии достигается при условии прекращения роста, прохождения фаз закаливания и вступления их в период покоя [3]. Известно, что в листьях и рожке земляники в осенний период при закалке происходит усиленный биосинтез антифризных и запасных соединений, а также изменяется водный статус, что в дальнейшем способствует растению переносить гипотермические условия зимы. Наряду с этим земляника относится к летне-зимне-зеленым растениям со слабо выраженным периодом покоя [4]. В связи с этим на фоне действия отрицательных температур или их перепадов в тканях растений может отмечаться развитие окислительного стресса.
Как правило, растения на стрессовые воздействия реагируют однотипно, но при этом отличаются по устойчивости и скорости физиологических и структурных перестроек [5]. Разный уровень устойчивости обусловлен физиолого-биохимическими особенностями сортов.
В связи с этим представляло интерес изучить особенности антиоксидантного и водного статуса, окислительного стресса и накопления протекторных соединений в осенний период у сортов земляники садовой разного эколого-географического происхождения в условиях Орловской области Российской Федерации.
Условия, материалы и методы
Объектами исследований за период сентябрь-ноябрь служили листья сортов земляники садовой различного эколого-географического происхождения: Кокинская ранняя, Соловушка, Урожайная ЦГЛ, Царица (Россия); Alba (Италия); Sara (Швеция); Korona (Голландия).
Для изучения состояния водного режима земляники садовой определяли фракционный состав воды в листьях методом Окунцова-Маринчик согласно методическим рекомендациям [6]. Метод основан на изменении концентрации раствора сахарозы при погружении в него ткани листа. Во взвешенные на весах бюксы наливают 2 мл 30% раствора сахарозы. Листья земляники садовой измельчают в лабораторной мельнице. Подготовленные пробы листьев массой 0,4 г погружают в 30% раствор сахарозы. Исходя из исходного объема раствора, начальной и конечной его концентрации, определяют количество воды, отнятой раствором из тканей листа. По разнице содержания общей воды и воды, перешедшей в раствор, рассчитывают содержание связанной воды. Концентрацию сахарозы в растворе определяют на рефрактометре.
Для оценки формирования устойчивости земляники к стрессу использовали методические рекомендации [7]. Так об интенсивности окислительного стресса судили по развитию перекисного окисления липидов (ПОЛ) на основе определения малонового диальдегида (МДА) по качественной реакции с тиобарбитуровой кислотой и гидроперекисей липидов, используя роданид аммония. О количестве активных форм кислорода судили по содержанию пероксида водорода с помощью тетрахлористого титана. Для изучения антиоксидантного статуса определяли содержание свободного пролина с помощью нингидринового реактива, а также активность ферментов: супероксид-дисмутазы (СОД), используя нитросиний тетразолий, и каталазы – методом учета выделяющегося кислорода. Используя резорциновый реактив, устанавливали уровень содержания сахарозы в тканях листа [8]. Достоверность результатов оценивали в программе MS Exel по Доспехову [9].
Результаты и обсуждения
При формировании устойчивости существенное значение имеет функционирование антиоксидантной системы защиты, которая предотвращает развитие окислительного стресса и в частности перекисного окисления мембранных липидов (ПОЛ) на фоне действия неблагоприятных факторов среды.
Об интенсивности повреждения клеточных мембран судили по накоплению конечного продукта ПОЛ – малонового диальдегида (МДА), промежуточного продукта ПОЛ – гидроперекисей, содержанию перекиси водорода (как одного из представителей активных форм кислорода) и активности антиоксидантных ферментов: супероксид-дисмутазы (СОД), каталазы и пероксидазы. Результаты исследования показали, что по мере понижения температуры окружающей среды интенсивность накопления МДА у всех сортов увеличивалась. Однако у сортов Соловушка, Sara, Korona и Царица интенсивность повреждения мембранных липидов была существенно ниже, чем у остальных генотипов. Как видно из таблицы 1, у сортов Соловушка, Sara, Korona и Царица содержание малонового диальдегида в ноябре увеличилось на 17,5-25,7% по сравнению с октябрем, тогда как у остальных сортов повысилось на 31,7-56,3%. При этом абсолютные показания МДА у сортов Соловушка, Sara, Korona и Царица в ноябре были ниже по сравнению с другими, что свидетельствует о более значительном повреждении структурно-функциональной целостности клеточных мембран у остальных сортов.
Разный уровень накопления МДА в исследуемых сортах, по-види-мому, связан как с разной степенью образования в клетках активных форм кислорода, так и более высокой активностью антиоксидантной системы защиты, нейтрализующей активные формы кислорода. В связи с этим, возникла необходимость определения перекиси водорода как одного из представителей активных форм кислорода.
При определении пероксида водорода была показана корреляционная связь между МДА и H 2 O 2 (в октябре – 0,67, а в ноябре – 0,79). В сортах Соловушка, Korona, Царица и Sara не только уровень накопления перекиси водорода был значительно ниже, но и снижены абсолютные значения H 2 O 2 как в октябре, так и в ноябре по сравнению с остальными генотипами (табл. 1).
Вместе с тем, известно, что в присутствии ионов двухвалентного железа перекись водорода может разлагаться с образованием гидроксильного радикала (реакция Фентона), который чрезвычайно активен и разрушает почти любую встретившуюся ему молекулу [10]. Инициирование цепной реакции (ПОЛ) начинается с того, что в липидный слой мембран или липопротеинов внедряется свободный радикал. Чаще всего этим радикалом и является радикал гидроксила. Будучи небольшой по размеру незаряженной частицей, он способен проникать в толщу гидрофобного липидного слоя и вступать в химическое взаимодействие с полиненасыщенными жирными кислотами, входящими в состав биологических мембран [11].
Гидроксильный радикал, отнимающий водород от СН 2 - групп ненасыщенных жирных кислот липидов, преобразуется в липидный радикал. Липидный радикал вступает в реакцию с растворенным в среде молекулярным кислородом, при этом образуется новый свободный радикал – радикал липоперекиси. Этот радикал атакует одну из соседних молекул фосфолипида с образованием гидроперекиси липида (ГП) и нового липидного радикала. Чередование двух последних реакций и представляет собой цепную реакцию пероксидации (перекисного окисления) липидов. Образующиеся радикалы инициируют новые цепи окисления липидов [12]. Весте с тем, перекиси липидов – нестойкие вещества и легко распадаются [13]. В местах присоединения перекисных радикалов жирные кислоты разрываются на фрагменты, на краях которых расположены альдегидные группы, обладающие высокой реакционной способностью. Если разрыв произошел с двух сторон, происходит образование более устойчивых вторичных продуктов, к которым относят кислородсодержащие соединения: альдегиды и диальдегиды, в частности, малоновый диальдегид [14]. Таким образом, количественное определение гидроперекисей позволяет непосредственно судить об интенсивности протекании свободно-радикальных процессов.
Проведенный анализ по содержанию гидроперекисей липидов – промежуточных продуктов ПОЛ показал, что пик образования последних в исследуемых сортах пришелся на октябрь. В ноябре на фоне увеличения МДА уровень ГП характеризовался снижением своих значений относительно октября. Причем коэффициент корреляции между содержанием гидроперекисей и малоновым диальдегидом в октябре составлял 0,77, а в ноябре – 0,96. Вместе с тем, остаются неясными причины снижения гидроперекисей в ноябре, тогда как уровень МДА и перекиси водорода возрастали. Возможно, это связано как с увеличением активности каталазы, так и с увеличением ее биосинтеза, а, следовательно, и с уменьшением доли свободных двухвалентных катионов железа в цитозоле (входят в активный центр фермента) (табл. 1), что в дальнейшем способствовало нейтрализации пероксида водорода и препятствовало его преобразованию в гидроперекиси. Кроме того, уменьшение доли свободных двухвалентных ионов железа могло произойти и в результате уменьшения доли свободной воды в цитозоле, их включению в активные центры других ферментов, о чем косвенно свидетельствует увеличение содержания пептидных соединений. Все эти вопросы требуют дальнейших детальных исследований. Вместе с тем, сорта Соловушка, Sara, Korona и Царица в течение двух осенних месяцев, характеризовались низким уровнем гидроперекисей, что может дополнительно свидетельствовать о низком темпе процессов липопероксидации.
Анализ активности антиоксидантных ферментов показал их существенную интенсивность работы у сортов с низким уровнем МДА, гидроперекисей и перекиси водорода, за некоторым исключением для супероксиддисмутазы. Так у сортов Соловушка, Царица, Корона
Таблица 1. Содержание МДА, перекиси водорода и гидроперекисей липидов в листьях земляники Table 1. Content of MDA, hydrogen peroxide and lipid hydroperoxides in the leaves of strawberry plants
|
Сорт |
МДА, мкМоль/г |
Перекись водорода, мкМоль/г |
Гидроперекиси липидов, усл. ед. |
|||
|
октябрь |
ноябрь |
октябрь |
ноябрь |
октябрь |
ноябрь |
|
|
Соловушка |
3,8±0,28 |
4,5±0,25 |
1,5±0,05 |
3,5±0,12" |
2,22±0,04 |
1,51±0,04" |
|
Sara |
4,0±0,12 |
4,7±0,20" |
1,9±0,10 |
3,9±0,12" |
2,32±0,05 |
1,33±0,03" |
|
Урожайная ЦГЛ |
3,9±0,12 |
5,8±0,17" |
2,2±0,17 |
7,4±0,30" |
2,85±0,07 |
1,67±0,04" |
|
Alba |
4,8±0,17 |
7,5±0,15" |
2,9±0,15 |
16,9±0,21" |
3,03±0,08 |
2,88±0,08 |
|
Кокинская ранняя |
4,1±0,14 |
5,4±0,23" |
3,1±0,15 |
15,7±0,32" |
2,97±0,08 |
2,12±0,05" |
|
Царица |
3,7±0,13 |
4,5±0,18" |
1,6±0,06 |
2,7±0,09" |
2,32±0,07 |
1,31±0,03" |
|
Korona |
3,5±0,12 |
4,4±0,20" |
1,7±0,06 |
2,9±0,11" |
2,11±0,06 |
1,32±0,04" |
|
НСР 0,05 |
0,15 |
0,17 |
0,14 |
0,31 |
0,11 |
0,15 |
" достоверность отличий против октября при Р<0,05
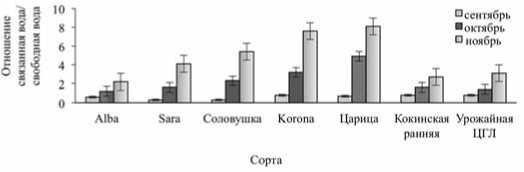
Рис. Отношение связанная вода/свободная вода в листьях сортов земляники садовой в осенний период
Fig. The ratio of bound water/free water in the leaves of strawberry varieties in the autumn period и Sara активность СОД, фермента, утилизирующего супероксидра-дикал с образованием перекиси водорода, достоверно не изменился в ноябре по сравнению с октябрем, тогда как у остальных генотипов она повысилась на 14,6-22,2% (табл. 2). Это в какой-то степени с одной стороны объясняет разный уровень перекиси водорода и гидроперекисей в исследуемых сортах.
Кроме того, изучение активности другого антиоксидантного фермента – каталазы, показало ее значительную интенсификацию у сортов Соловушка, Царица, Корона и Sara. Активность расщепления перекиси водорода каталазой у данных сортов увеличилась в ноябре на 37,7-50,5% против октября, тогда как у остальных – на 15,7-25,5% (табл. 2).
Состояние водного режима в осенний период является одним из важных факторов, определяющих успешную перезимовку растений [15].
Известно, что у зимостойких сортов плодовых культур отношение связанная/свободная вода выше, чем у не зимостойких [16]. При определении фракционного состава воды в сентябре в листьях земляники отметили низкое отношение связанной воды к свободной. В
Таблица 2. Активность СОД и каталазы в листьях растений земляники Table 2. SOD and Catalase activity in the leaves of strawberry plants
|
Сорт |
СОД, усл.ед. |
Каталаза, мл O2/мин |
||
|
октябрь |
ноябрь |
октябрь |
ноябрь |
|
|
Соловушка |
55,5±1,20 |
54,2±1,25 |
10,5±0,50 |
15,2±0,36" |
|
Sara |
66,9±2,39 |
63,1±2,06 |
12,2±0,35 |
16,8±0,50" |
|
Урожайная ЦГЛ |
69,2±1,91 |
79,3±1,29" |
10,8±0,46 |
12,5±0,40" |
|
Alba |
53,6±1,64 |
65,5±3,04" |
11,7±0,35 |
14,1±0,47" |
|
Кокинская ранняя |
71,0±0,86 |
83,5±3,21" |
9,8±0,36 |
12,3±0,42" |
|
Царица |
53,4±1,87 |
52,5±1,78 |
10,7±0,40 |
16,1±0,44" |
|
Korona |
52,7±1,84 |
51,5±1,55 |
11,4±0,44 |
15,8±0,50" |
|
НСР 0,05 |
0,43 |
1,10 |
1,10 |
1,50 |
" достоверность отличий против октября при Р<0,05
Таблица 3. Содержание сахарозы и свободного пролина в листьях растений земляники Table 3. The content of sucrose and free proline in the leaves of strawberry plants
|
Сорт |
Сахароза, мг/г |
Пролин, мг/кг |
||||
|
сентябрь |
октябрь |
ноябрь |
сентябрь |
октябрь |
ноябрь |
|
|
Соловушка |
1,30±0,06 |
2,73±0,15* |
2,65±0,13* |
1,19±0,06 |
13,34±0,67* |
19,38±0,90*" |
|
Sara |
0,98±0,05 |
3,50±0,14* |
2,68±0,12*" |
6,07±0,30 |
19,87±0,98* |
27,55±1,38*" |
|
Урожайная ЦГЛ |
1,54±0,08 |
3,65±0,15* |
2,23±0,10*" |
1,87±0,11 |
12,48±0,68* |
14,93±0,79* |
|
Alba |
1,78±0,09 |
3,41±0,17* |
2,04±0,11*" |
2,03±0,11 |
11,80±0,59* |
14,90±0,74*" |
|
Кокинская ранняя |
1,32±0,07 |
2,57±0,11* |
2,31±0,11* |
2,34±0,12 |
14,95±0,79* |
24,28±1,33*" |
|
Царица |
1,43±0,05 |
7,05±0,28* |
8,34±0,29*" |
3,95±0,15 |
14,34±0,49* |
27,65±1,11*" |
|
Korona |
1,15±0,05 |
4,16±0,15* |
6,46±0,26*" |
2,65±0,11 |
11,32±0,40* |
28,73±1,15*" |
|
НСР 0,05 |
0,18 |
0,20 |
0,19 |
0,31 |
0,72 |
1,67 |
*достоверность отличий против сентября при Р<0,05 " достоверность отличий против октября при Р<0,05
октябре это отношение увеличилось в 1,8-7,7 раз. В ноябре оно было выше (в 1,6-18,0 раз), чем в предыдущие осенние месяцы (рис.). Наибольший показатель фракций воды к началу зимовки отметили у сортов – Соловушка, Царица, Sara, Korona.
Таким образом, видно, что в осенний период для растений земляники садовой характерно увеличение связанной и снижение свободной воды в листьях. При этом у сортов Соловушка, Царица, Sara, Korona к концу осени определили наибольший показатель отношения связанная вода/свободная.
Согласно ряду исследователей, пролин оказывает осмопротек-торные свойства в стрессовых условиях [17,18]. Важную роль в качестве осмопротекторов наряду с пролином играют и низкомолекулярные углеводы [19]. Сахара увеличивают водоудерживающую способность коллоидов протоплазмы, защищая от образования льда и чрезмерного обезвоживания клеток. В наших исследованиях, на фоне понижения температуры воздуха, в октябре, отмечалось значительное в 1,92-3,57 раза накопление в листьях земляники содержания сахаров (табл. 3). По мере снижения температуры воздуха в период сентябрь-ноябрь отмечали увеличение содержания в листьях и аминокислоты пролин. При холодовой адаптации увеличение содержания пролина зарегистрировано у растений земляники другими авторами [20]. В наших исследованиях наиболее интенсивный пик накопления пролина у всех сортов отмечался в октябре (в 3,27-11,21 раза по сравнению с сентябрем), тогда как в ноябре (в 1,19-1,60 раза против октября) (табл. 3). Максимальным накоплением сахаров отличались сорта земляники Sara, Korona, Царица и Урожайная ЦГЛ (количество углеводов в октябре увеличилось в 2,37-4,93 раза по сравнению с сентябрем) (табл.3).
У сортов Соловушка и Кокинская ранняя количество сахаров по сравнению с сентябрем увеличилось в меньшей степени – в 1,95 и 2,10 раза, соответственно. При этом наибольшим уровнем накопления аминокислоты характеризовались сорта Соловушка и
Кокинская ранняя. Количество пролина в данных сортах в ноябре по сравнению с сентябрем возросло в 16,20 и 10,38 раза, соответственно. Вместе с тем, по абсолютным значениям наибольший уровень пролина в ноябре отмечался у сортов Sara, Korona, Соловушка, Кокинская ранняя и Царица.
В ноябре по мере снижения температуры у некоторых сортов отмечали уменьшение количества сахаров относительно уровня октября, что может быть связано с их активной тратой на связывание свободной воды в качестве энергетического субстрата на процессы дыхания, синтез белка, аминокислот и т.д. При этом в большей степени содержание сахаров уменьшилось у сортов Sara, Урожайная ЦГЛ и Alba (в 1,31-1,57 раза относительно октября). Вместе с тем у сортов Царица и Korona содержание сахаров продолжало увеличиваться. У сортов Соловушка и Кокинская ранняя изменение уровня сахаров было не достоверно. Как и в случае с пролином, наибольшим содержанием сахаров по абсолютным значениям в ноябре выделились сорта Соловушка, Sara, Царица и Korona, наименьшим – Alba.
Выводы
Таким образом, установлено, что в осенний период при подготовке к зиме у растений земляники садовой отмечено увеличение связанной и снижение свободной воды в листьях. При этом к концу осени у сортов Соловушка, Царица, Sara и Korona на фоне высокого содержания свободного пролина и сахаров наблюдали наибольший показатель отношения связанная/свободная вода, что говорит о высокой адаптационной способности данных сортов. Наряду с этим, сорта Соловушка, Царица, Sara и Korona, характеризовались и повышенной антиоксидантной активностью на примере ферментов СОД и каталазы, незначительным уровнем образования активных форм кислорода и, как следствие, низким структурно-функциональным повреждением клеточных мембран.
Об авторах:
Pavel S. Prudnikov – Candidate of Biological Sciences
Zoya E. Ozherelieva – Candidate of Agricultural Sciences ,
ResearcherID: C-7380-2017
Diana A. Krivushina – Graduate student
Marina I. Zubkova – researcher
-
• Литература
-
1. Говорова Г.Ф., Говоров Д.Н. Земляника: прошлое, настоящее, будущее. М., 2004. 347 с.
-
2. Абызов В.В. Биологические особенности и хозяйственная ценность сортов земляники в условиях средней полосы России: автореф. дис. …канд. с.-х. наук. Мичуринск-наукоград РФ, 2008. 23 с.
-
3. Стольникова Н.П. Культура земляники в Западной Сибири. ФГБНУ НИИСС. Барнаул: ИП Колмогоров И.А., 2014. 182 с.
-
4. Серебрякова Т.Ю., Воронин Н.С., Еленевский А.Г., Батыгина Т.Б., Н.И. Шорина Н.И. Ботаника с основами фитоценологии. Анатомия и морфология растений. М.: ИКЦ «Академкнига», 2006. 543 с.
-
5. Удовенко Г.В. Механизмы адаптации растений к стрессам. Физиология и биохимия культурных растений. 1979;11(2):99-107.
-
6. Ожерельева З.Е., Прудников П.С., Зубкова М.И., Кривушина Д.А., Князев С.Д. Определение морозостойкости земляники садовой в контролируемых условиях (методические рекомендации). Орел: ВНИИСПК, 2019. 25 с.
-
7. Прудников П.С., Ожерельева З.Е. Физиолого-биохимические методы диагностики устойчивости плодовых культур к засухе и гипертермии (методические рекомендации). Орел: ВНИИСПК, 2019. 46 с.
-
8. Туркина М.В., Соколова C.B. Изучение мембранного транспорта сахарозы в растительной ткани. Физиология растений. 1972;19(5):912-919.
-
9. Доспехов Б.А. Методика полевого опыта. М.: Агропромиздат, 1985. 351 с.
-
10. Владимиров Ю.А., Азизова О.А., Деев А.И. Свободные радикалы в живых системах. Итоги науки и техники. Биофизика. 1999;(29):249.
-
11. Пискарев И.М., Иванова А.Г., Самоделкин М.Н., Иващенко М.Н. Инициирование и исследование свободно-радикальных процессов в биологических экспериментах. Н. Новгород: ФГБОУ ВО Нижегородская ГСХА, 2016. 140 с.
-
12. Skorzynska-Polit E. Lipid peroxidation on plant cells, its physiological role and changes under heavy metal stress. Acta Societatis Botanicorum Poloniae. 2007;74:49-54.
-
13. Лелевич В.В. Биологическая химия. ГрГМУ. Гродно. 2009. 275 с.
-
14. Halliwell, B., Gutteridge, J.M.C.: The importance of free radicals and catalitic metal ions in human diseases. Molec. Aspects Med. 1985;(8):89-193.
-
15. Ожерельева З.Е. Изменения водного режима сортов земляники садовой в осенний период. Плодоводство, семеноводство, интродукция древесных растений. 2018;(21):166-169.
-
16. Кушниренко М.Д. Физиология водообмена и засухоустойчивости плодовых культур. Кишинёв: Штиинца, 1975. 215 с.
-
17. Джавадиан Н., Каримзаде Г., Мафузи С., Ганати Ф. Вызванные холодом изменения активности ферментов и содержания пролина, углеводов и хлорофиллов у пшеницы. Физиология растений. 2010;57(4):580-588.
-
18. Прудников П.С., Кривушина Д.А., Гуляева А.А. Компоненты антиоксидантной системы и интенсивность ПОЛ Prunus cerasus L. при действии гипертермии и засухи. Селекция и сорторазведение садовых культур. 2016;(3):116-119.
-
19. Чудинова Л.А., Орлова Н.В. Физиология устойчивости растений: учеб. пособие к спецкурсу. Пермь. Перм. ун-т, 2006. 124 с.
-
20. Luo Y., Tang H. Zhang Y. Production of reactive oxygen species and antioxidant metabolism about strawberry leaves to low temperatures. J. Agr. Sci. 2011;(3):89-96.
-
-
• References
-
1. Govorova G.F., Govorov D.N. Strawberries: past, present, future. M., 2004. 347 p. (In Russ.)
-
2. Abyzov VV Biological features and economic value of strawberry varieties in the middle zone of Russia: abstract. dis....cand. sciences. Michurinsk, 2008. 23 p. (In Russ.) 3. Stolnikova N.P. Strawberry culture in Western Siberia. Barnaul: IP Kolmogorov I.A., 2014. 182 p. (In Russ.)
-
4. Serebryakova T.Yu., Voronin N.S., Elenevsky A.G., Batygina T.B., Shorina N.I., Savinykh N.P. Botany with the basics of phytocenology. Anatomy and morphology of plants. M.: IKC "Akademkniga", 2006. 543 p. (In Russ.)
-
5. Udovenko G.V. Mechanisms of plant adaptation to stresses. Physiology and biochemistry of cultivated plants. 1979;11(2):99-107. (In Russ.)
-
6. Ozherelieva Z.E., Prudnikov P.S., Zubkova M.I., Krivushina D.A., Knyazev S.D. Determination of frost resistance of strawberry garden under controlled conditions (guidelines). Orel: VNIISPK, 2019. 25 p. (In Russ.)
-
7. Prudnikov P.S., Ozherelieva Z.E. Physiological and biochemical methods for the diagnosis of resistance of fruit crops to drought and hyperthermia (guidelines). Orel: VNIISPK, 2019. 46 p. (In Russ.)
-
9. Dospekhov B.A. Methodology of field experience. M.: Agropromizdat, 1985. 351 p. (In Russ.)
-
10. Vladimirov Yu.A., Azizova O.A., Deev A.I. Free radicals in living systems // Itogi Nauki i Tekhniki. Biophysics. 1999;29:249. (In Russ.)
-
11. Piskarev I.М., Ivanova A.G., Samodelkin M.N., Ivashchenko M.N. Initiation and research of free-radical processes in biological experiments. N. Novgorod, 2016. 140 p. (In Russ.)
-
12. Skorzynska-Polit E. Lipid peroxidation on plant cells, its physiological role and changes under heavy metal stress. Acta Societatis Botanicorum Poloniae. 2007;(74):49-54.
-
13. Lelevich V.V. Biological chemistry. State Medical University. Grodno. 2009. 275 p. (In Russ.)
-
14. Halliwell, B., Gutteridge, J.M.C. The importance of free radicals and catalitic metal ions in human diseases. Molec. Aspects Med. 1985;(8):89-193.
-
15. Ozherelieva Z.E. Changes in the water regime of varieties of garden strawberries in the autumn period. Fruit growing, seed growing, introduction of woody plants. 2018;(21):166-169. (In Russ.)
-
16. Kushnirenko M.D. Physiology of water exchange and drought tolerance of fruit crops. Chisinau: Shtiintsa, 1975. 215 p. (In Russ.)
-
17. Javadian N., Karimzade G., Mafuzi S., Ganati F. Cold-induced changes in enzyme activity and the content of proline, carbohydrates and chlorophylls in wheat. Plant Physiology. 2010;57(4):580-588. (In Russ.)
-
18. Prudnikov P.S., Krivushina D.A., Gulyaev A.A. The components of the antioxidant system and the LPO intensity of Prunus cerasus L. under the action of hyperthermia and drought. Selection and cultivation of garden crops. 2016;(3):116-119. (In Russ.)
-
19. Chudinova L.A., Orlova N.V. Physiology of plant resistance: textbook. allowance for the special course. Permian. Perm. Univ., 2006. 124 p. (In Russ.)
-
20. Luo Y., Tang H. Zhang Y. Production of reactive oxygen species and antioxidant metabolism about strawberry leaves to low temperatures. J. Agr. Sci. 2011;(3):89-96.
-
Список литературы Формирование устойчивости к гипотермии сортов Fragaria x ananassa Duch. разного эколого-географического происхождения в осенний период
- Говорова Г.Ф., Говоров Д.Н. Земляника: прошлое, настоящее, будущее. М., 2004. 347 с.
- Абызов В.В. Биологические особенности и хозяйственная ценность сортов земляники в условиях средней полосы России: автореф. дис..канд. с.-х. наук. Мичуринск-наукоград РФ, 2008. 23 с.
- Стольникова Н.П. Культура земляники в Западной Сибири. ФГБНУ НИИСС. Барнаул: ИП Колмогоров И.А., 2014. 182 с.
- Серебрякова Т.Ю., Воронин Н.С., Еленевский А.Г., Батыгина Т.Б., Н.И. Шорина Н.И. Ботаника с основами фитоценологии. Анатомия и морфология растений. М.: ИКЦ "Академкнига", 2006. 543 с.
- Удовенко Г.В. Механизмы адаптации растений к стрессам. Физиология и биохимия культурных растений. 1979;11(2):99-107.
- Ожерельева З.Е., Прудников П.С., Зубкова М.И., Кривушина Д.А., Князев С.Д. Определение морозостойкости земляники садовой в контролируемых условиях (методические рекомендации). Орел: ВНИИСПК 2019. 25 с.
- Прудников П.С., Ожерельева З.Е. Физиолого-биохимические методы диагностики устойчивости плодовых культур к засухе и гипертермии (методические рекомендации). Орел: ВНИИСПК, 2019. 46с.
- Туркина М.В., Соколова C.B. Изучение мембранного транспорта сахарозы в растительной ткани. Физиология растений. 1972;19(5):912-919.
- Доспехов Б.А. Методика полевого опыта. М.: Агропромиздат, 1985. 351 с.
- Владимиров Ю.А., Азизова О.А., Деев А.И. Свободные радикалы в живых системах. Итоги науки и техники. Биофизика. 1999;(29):249.
- Пискарев ИМ., Иванова А.Г., Самоделкин М.Н., Иващенко М.Н. Инициирование и исследование свободно-радикальных процессов в биологических экспериментах. Н. Новгород: ФГБОУ ВО Нижегородская ГСХА, 2016. 140 с.
- Skorzynska-Polit E. Lipid peroxidation on plant cells, its physiological role and changes under heavy metal stress. Acta Societatis Botanicorum Poloniae. 2007;74:49-54.
- Лелевич В.В. Биологическая химия. ГрГМУ. Гродно. 2009. 275 с.
- Halliwell, B., Gutteridge, J.M.C.: The importance of free radicals and catalitic metal ions in human diseases. Molec. Aspects Med. 1985;(8):89-193.
- Ожерельева З.Е. Изменения водного режима сортов земляники садовой в осенний период. Плодоводство, семеноводство, интродукция древесных растений. 2018;(21):166-169.
- Кушниренко М.Д. Физиология водообмена и засухоустойчивости плодовых культур. Кишинёв: Штиинца, 1975. 215 с.
- Джавадиан Н., Каримзаде Г., Мафузи С., Ганати Ф. Вызванные холодом изменения активности ферментов и содержания пролина, углеводов и хлорофиллов у пшеницы. Физиология растений. 2010.57(4):580-588.
- Прудников П.С., Кривушина Д.А., Гуляева А.А. Компоненты антиоксидантной системы и интенсивность ПОЛ Prunus cerasus L. при действии гипертермии и засухи. Селекция и сорторазведение садовых культур. 2016;(3):116-119.
- Чудинова Л.А., Орлова Н.В. Физиология устойчивости растений: учеб. пособие к спецкурсу. Пермь. Перм.ун-т, 2006. 124 с.
- Luo Y., Tang H. Zhang Y. Production of reactive oxygen species and antioxidant metabolism about strawberry leaves to low temperatures. J. Agr. Sci. 2011;(3):89-96.
