Фотосинтетическая активность листьев кукурузы в центральном агроклиматическом районе Республики Коми

Автор: Т.К. Головко, Р.В. Малышев, Г.Н. Табаленкова
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Статья в выпуске: 7 (83), 2025 года.
Бесплатный доступ
Кукуруза – одна из основных культур современного мирового растениеводства, относится к группе теплолюбивых растений. Наблюдаемое в последние десятилетия потепление климата в Северном полушарии, появление раннеспелых и холодостойких сортов и гибридов создают предпосылки для выращивания кукурузы на зеленую массу на Севере. Нами установлено, что в условиях центрального агроклиматического района Республики Коми современные гибриды кукурузы способны накапливать 350-500 ц/га зеленой массы. При температуре +20º С и высокой освещенности листья холодостойких раннеспелых гибридов Дорка и Уральский 150 ассимилировали СО2 со скоростью 20–25 мкмоль/м2с. Зона температурного оптимума фотосинтеза составляла +18... +28º С. Сделано заключение о способности раннеспелых и холодостойких гибридов кукурузы активно фотосинтезировать и формировать урожай зеленой массы при умеренной температуре.
Кукуруза, фотосинтез, дыхание, продуктивность, температура, холодный климат
Короткий адрес: https://sciup.org/149149452
IDR: 149149452 | УДК: 581.1:581.132:633.15 | DOI: 10.19110/1994-5655-2025-7-109-115
Photosynthetic activity of corn leaves in the central agro-climatic region of the Komi Republic
Corn is among the main crops in modern world crop production. It belongs to the group of heat-loving plants. The climate warming observed in the Northern Hemisphere in recent decades, as well as the availability of early-maturing and cold-resistant varieties and hybrids, create opportunities for growing of corn for green mass in the North. We have found that the modern corn hybrids can accumulate 350–500 centners per hectare of green mass in the central agro-climatic region of the Komi Republic. At a temperature of +20º C and the high light intensity, leaves of the cold-resistant, early-ripening hybrids Dorka and Uralskiy 150 assimilate CO2 at a rate of 20–25 μmol/m2s. The temperature optimum for photosynthesis is +18.. +28º C. The authors have concluded about the ability of early-ripening and cold-resistant corn hybrids to actively photosynthesise and yield at moderate temperatures.
Текст научной статьи Фотосинтетическая активность листьев кукурузы в центральном агроклиматическом районе Республики Коми
Продуктивность является интегральным результатом всех функций растительного организма, зависит от генотипических особенностей и физиолого-биохимических свойств видов и сортов, их взаимодействия со средой. В физиологически приемлемых условиях фотосинтетический аппарат растений обеспечивает реализацию генетически детерминированной программы роста и развития, используя энергию солнечной радиации и другие ресурсы (СО2, вода, минеральные элементы). Значительная часть территории РФ, в том числе Республика Коми, расположена на Севере в зоне рискованного земледелия [1-3]. Сельскохозяйственное производство растениеводческой продукции ограничивается здесь коротким вегетационным периодом и недостатком тепла, что определяет подбор возделываемых культур с учетом критериев соответствия их метаболизма климату. Приоритетное значение в этом отношении имеют изучение физиолого-биохимических и экологических закономерностей формирования продуктивности, выявление особенностей продукционного процесса в условиях холодного климата.
Кукуруза – одна из основных культур современного мирового растениеводства. По валовым сборам зерна она занимает третье место в мире после пшеницы и риса. Кукуруза – растение тропического происхождения с С4– типом фотосинтеза. С4–тип углеродного метаболизма характеризуется наличием двух фотосинтетических циклов, локализованных в различных клетках листа [4]. Это обеспечивает эффективное использование растениями воды и реализацию потенциала продуктивности данной культуры в регионах с теплым засушливым климатом. В Европе кукурузу на зерно возделывают до 52о с. ш., на силос – до 60о с. ш. В полевых условиях ведущим фактором, опреде- ляющим уровень и суточную динамику скорости ростовых процессов кукурузы, является температура. При благоприятном световом режиме и увлажнении максимальная скорость роста в условиях средней полосы России была зарегистрирована при температуре +28...+30о С [5]. Нижняя граница оптимальной зоны температур зависела от комплексного влияния факторов внешней среды и в большинстве случаев отмечена в диапазоне +20...+22° С.
Сложилось мнение, что условия центрального и южного агроклиматических районов Республики Коми (59–62 с. ш.) не пригодны для выращивания кукурузы. Опыты по ее возделыванию, проведенные в середине прошлого столетия, позволили установить некоторые особенности роста растений и сделать вывод о возможности получения достаточно высоких урожаев зеленой массы (до 500 ц/га и более), а в благоприятные годы и початков молочно-восковой спелости [6]. Однако культура не получила распространения из-за риска потери урожая в годы с холодным летом. Наблюдаемое в последние десятилетия заметное потепление климата в Северном полушарии, появление раннеспелых и холодостойких сортов и гибридов, использование современных технологий создают условия для выращивания кукурузы на зеленую массу в Сибири [7, 8] и на европейском северо-востоке России [9, 10]. За последние полвека сумма температур >10о С в центральном агроклиматическом районе РК возросла на 200о С, а продолжительность вегетации – почти на неделю. По прогнозам, к 2080 г. лето на территории республики может потеплеть на 6-7о С. Это создает условия для экспансии зоны возделывания кукурузы для получения зеленой массы на Севере.
Предварительное экологическое сортоиспытание в 2016-2017 гг. семи сортообразцов и гибридов кукурузы современной селекции подтвердило возможность получения в центральном агроклиматическом районе РК в среднем 350 ц/га зеленой массы хорошего качества [9, 11]. При этом величина урожая колебалась в зависимости от генотипа и погодных условий в пределах 240770 ц/га. За три года (2018–2020) средняя продуктивность надземной массы гибрида кукурузы Дорка составила 355 ц/га [10], что сопоставимо со средними урожаями кукурузы, возделываемой на силос, по РФ [12].
Фотосинтез является основой продукционного процесса, но систематических исследований фотосинтетической деятельности кукурузы на территории республики практически не проводилось. Актуальность таких исследований обусловлена необходимостью изучения физиологических основ урожайности кукурузы в условиях холодного климата, характеристики устойчивости сортов и гибридов, их пригодности для использования в северном растениеводстве.
Цель работы – изучение фотосинтеза листьев двух холодостойких и раннеспелых гибридов кукурузы Дорка и Уральский 150.
Материалы и методы
Растения гибридов кукурузы Дорка и Уральский 150 выращивали на опытном участке Института биологии Коми НЦ УрО РАН. Почва опытного участка подзолистая, средней степени окультуренности. Для характеристики сезонных изменений фотосинтетической активности измеряли скорость СО2-обмена в средней части завершивших рост листьев верхнего яруса гибрида Дорка в открытой системе с помощью ИК-газоанализатора (LI-7000, США) при температуре +20° С и плотности потока фотосинтетически активной радиации (ФАР) от 0 до 1600 мкмоль квантов/м2с. В качестве источника света использовали систему красных (максимум при 634 нм) и синих (максимум при 447 нм) светодиодов в соотношении 11 : 1. Листья отделяли от растения и сразу же помещали в термостатируемую листовую камеру для измерения поглощения СО2 на свету и выделения СО2 в темноте. Биологическая повторность – 6–7, для каждой повторности использовали свежеотобранный лист. Измерения осуществляли в фазы пяти (первая декада июля), десяти (третья декада июля) листьев и в фазу выметывания метелки (третья декада августа). Для характеристики влияния температуры на фотосинтез кукурузы использовали простой гибрид Уральский 150. Определения проводили в наиболее теплом месяце – июле (среднесуточная температура – +17–18о С) в фазе пяти-семи листьев при температуре +10, 20, 30 и 40оС и плотности потока фотосинтетически активной радиации (ФАР) от 0 до 1600 мкмоль квантов/м2с по вышеуказанной схеме.
Зависимость фотосинтеза от освещенности описывали уравнением Михаэлиса-Ментен с модификациями [13, 14]:
Pn=[aI/(I + b)]–Rd (1), где Pn – скорость нетто-фотосинтеза; Rd – скорость темнового дыхания; I – интенсивность ФАР; а и b – константы.
Результаты и их обсуждение
Определения СО2-газообмена при температуре +20о С, близкой к среднесуточной в июле, показали, что фотосинтетическая активность листьев гибрида кукурузы Дорка возрастала с увеличением интенсивности ФАР (рис. 1 и 2). Наибольшую скорость видимого поглощения СО2 (Pn) регистрировали при интенсивности ФАР 1600 мкмоль кван-тов/м2с. У молодых растений (фаза четырех-пяти листьев) величина Pn составляла около 14 мкмоль СО2/м2с. У зрелых сформировавшихся растений этот показатель был на 40 % меньше, что свидетельствует о снижении способности листьев верхнего яруса использовать свет высокой интенсивности.
Полученные данные хорошо описывались уравнением (1) (r = 0,97), позволяющим оценить максимальную величину видимого поглощения СО2 (Pnмах) и интенсивность ФАР, при которой скорость поглощения СО2 достигает 50 % Pnмах. Как видно из табл. 1, величина Pnмах (коэффициент «а» в уравнении (1)) листьев молодых растений гибрида Дорка может достигать 24 мкмоль СО2/м2с, что на 40 % выше, чем получено нами при 1600 мкмоль квантов/м2с. Интенсивность ФАР, при которой возможно достижение
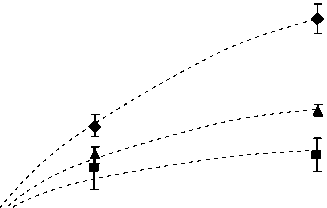
Таблица 1
Параметры кривых СО2-газообмена листьев верхнего яруса кукурузы сорта Дорка по модели [13, 14]
Table 1
Parameters of CO2-gas exchange curves in the upper-tier leaves of the Dorka hybrid corn according to the model [13, 14]
|
Фаза развития |
а, мкмоль СО2/м2с |
b, мкмоль квантов /м2с |
Rd, мкмоль СО2/м2с |
|
5 листьев |
23,8±1,0а |
902±83a |
1,5±0,1a |
|
10 листьев |
10,5±0,6b |
244±42b |
1,6±0,2a |
|
Выметывание метелки |
13,6±0,6b |
463±52c |
1,0±0,1b |
-2
ФАР, мкмоль/м2 с
Рисунок 1. Зависимость СО2-газообмена листьев верхнего яруса гибрида кукурузы Дорка от освещенности в разные сроки вегетации: 1 – фаза 5 листьев; 2 – фаза 10 листьев; 3 –фаза выметывания метелки. Условные обозначения. Здесь и на рис. 2: пунктиром обозначены кривые по модели [13, 14].
Figure 1. Light dependence of CO2-gas exchange of the upper leaves of Dorka hybrid corn at different periods of vegetation: 1– the phase of five leaves; 2 – the broom sweeping phase; 3 – the phase of ten leaves.
Key. Here and in Figure 2: The dotted lines indicate the curves according to the model [13, 14].
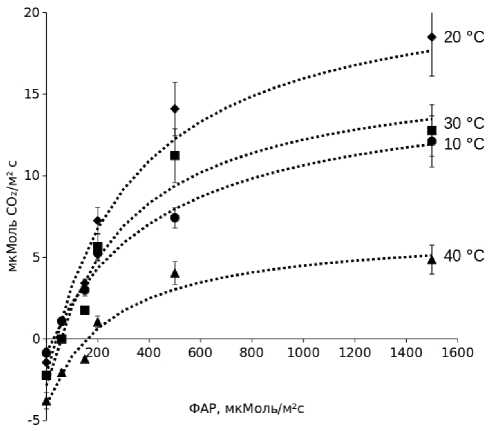
Рисунок 2. Зависимость СО2-газообмена листьев верхнего яруса гибрида кукурузы Уральский 150 от освещенности при разной температуре.
Figure 2. Light dependence of CO2-gas exchange of the upper leaves of Uralskiy 150 hybrid corn at different temperatures.
50 % Pnмах (коэффициент «b» в уравнении (1)), составляет около 1000 мкмоль квантов/м2с, что свидетельствует о высокой требовательности молодых растений к освещенности. На более поздних этапах развития растений (кривые 2 и 3) полученные по модели величины Pnмах были заметно (в два раза) ниже. Прямые определения и данные моделирования показывают, что с возрастом растений способность листьев верхнего яруса эффективно использовать свет высокой интенсивности для ассимиляции СО2 снижается, и выход световой кривой фотосинтеза на плато осуществляется при более низкой освещенности. Возможно,
Примечание. Здесь и в табл. 2: разными буквенными символами обозначены достоверные различия.
Note. Here and in Table 2: Different letter symbols indicate true differences. что такая реакция была обусловлена переходом растений к генеративному развитию.
Дыхание листьев кукурузы варьировало в пределах 1,0-1,5 мкмоль СО2/м2с и составляло от 6 до 15 % максимальной величины нетто-фотосинтеза. Следует заметить, что нетто-фотосинтез листьев кукурузы, возделываемой в более теплом климате, например, на севере Италии, достигал 40 мкмоль СО2/м2с [15], что почти вдвое выше величин, полученных нами для изученных гибридов.
На рис. 2 приведены данные, характеризующие влияние температуры на световую зависимость СО2-газооб-мена листьев гибрида кукурузы Уральский 150. Максимальное нетто-поглощение было зарегистрировано при температуре +20о С. Более низкие и высокие температуры снижали нетто-ассимиляцию, особенно заметно в области высокой плотности потока радиации. Причем, температура +10о С слабее влияла на процесс ассимиляции, чем температура +40о С. Проведенные нами ранее модельные эксперименты с выдерживанием растений гибрида Дорка в течение ночи при +6о С показали более слабое последействие пониженной температуры на фотосинтез, чем на дыхание листьев [16]. Эти данные свидетельствуют о способности раннеспелых холодостойких гибридов поддерживать сравнительно высокую функциональную активность при умеренных температурах.
Аппроксимация экспериментальных данных уравнением Михаэлиса-Ментен [13, 14] подтвердила наблюдаемые закономерности. Максимальные величины нетто-ассимиляции отмечены при температуре +20о С. Супероптимальная температура +40о С снижала величину Pnмах в два раза. Высокие температуры активировали темновое выделение СО2 (величина Rd), связанное с дыханием. Учитывая тот факт, что локализованный в клетках мезофилла листьев фермент фосфоенолпируваткарбок-силаза (ФЕПК) имеет более высокое сродство к диоксиду углерода, чем рибулезобифосфаткарбоксилаза (РБФК), восстанавливающая СО2, выделенный при декарбоксилировании С4-кислот в клетках, окружающих проводящие пучки, мы сочли возможным оценить величину гросс-фо-тосинтеза (Pg) листьев как сумму Pnмах (величина «а» в уравнении (1)) и Rd. Полученные таким способом величины Pg существенно не повлияли на характер зависимости фотосинтетической активности от температуры. Можно отметить незначительное сокращение разницы между величинами фотосинтеза, измеренными при +20о и +40о С. Если доля Pnмах (величина «а» в уравнении (1)) при +40о С составляла 44 % от величины Pnмах при +20о С, то разница между соответствующими величинами Pg была меньше из-за повышения темнового дыхания.
Экспериментально измеренные величины нетто-по-глощения СО2 при разной температуре и умеренной плотности потока ФАР (см. рис. 2), аппроксимировали уравнением параболы (рис. 3, табл. 2). Статистические показатели свидетельствуют об удовлетворительном соответствии экспериментальных данных модели. Используя уравнение (1), можно найти точки перехода кривой через ось х (т. е. нули функции). Эти значения представляют собой температуры, при которых Pn=0. В области низких положительных температур величина х равна 3,29, а в области высоких – 42,94. Следовательно, температуры, близкие к +3,3 и +43о С, можно рассматривать как критические для нетто-фотосинтеза листьев кукурузы. О способности кукурузы сохранять положительный газообмен при низких
Таблица 2
Параметры кривых СО2-газообмена листьев верхнего яруса гибрида кукурузы Уральский 150 по модели [13, 14]
Table 2
Parameters of CO2-gas exchange curves in the upper-tier leaves of the Uralskiy 150 hybrid corn according to the model [13, 14]
|
Температура, оС |
а, мкмоль СО2/м2с |
b, мкмоль квантов /м2с |
Rd, мкмоль СО2/м2с |
Pg, мкмоль СО2/м2с |
|
10 |
16,5±1,4a |
437±116a |
0,83±0,6 a |
17,3±2,0a |
|
20 |
24,4±1,9b |
347±86a |
2,1±0,9b |
26,5±2,8c |
|
30 |
19,6 ±1,9c |
301±91a |
2,9±0,9b |
22,5±2,8c |
|
40 |
10,7±0,9d |
259±73a |
4,0±0,5c |
14,7±1,4b |
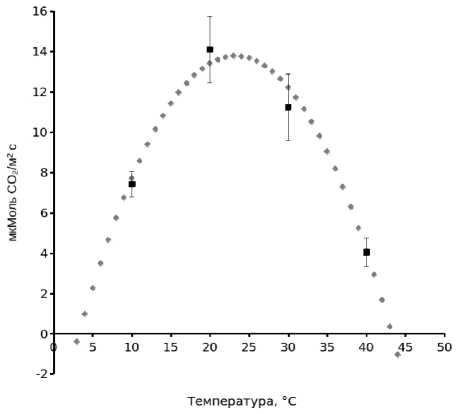
Рисунок 3. Зависимость СО2-газообмена листьев верхнего яруса гибрида кукурузы Уральский 150 от температуры при плотности потока ФАР 500 мкмоль квантов/м2с.
Условные обозначения. Пунктиром обозначена кривая аппроксимации экспериментальных данных уравнением параболы y=-0,03467914 x² + 1,603938 x + -4,888032, где у – скорость нетто-поглощения СО2, х – температура (R2=0,93; F=20,9; p=0,153).
Figure 3. The dependence of CO2-gas exchange in the upper-tier leaves of the Uralskiy 150 hybrid corn on the temperature at a PAR flux density of 500 μmol quanta/m2s.
Key. The dotted line represents the approximation of the experimental data by the parabolic equation y = -0,03467914 x² + 1,603938 x + -4,888032, where y is the net CO2 absorption rate and x – the temperature (R2 = 0.93; F = 20.9; p = 0.153).
положительных температурах свидетельствуют полученные нами ранее данные [17]. Скорость фотосинтеза листьев молодых растений гибрида Уральский 150 при +6° C составляла около 2 мкмоль СО2 /м2с. Оптимальными для нетто-поглощения СО2 считаются температуры, при которых скорость Pn снижается не более чем на 10-15 % от наибольшей величины на кривой, описывающей температурную зависимость данного показателя. На кривой (рис. 3) зона температурного оптимума Pn довольно широкая и находится в пределах +18...+28о С.
Заключение
В условиях центрального агроклиматического района РК раннеспелые и холодостойкие гибриды кукурузы накапливают за вегетацию 350-500 ц/га зеленой массы благодаря способности поддерживать сравнительно высокую фотосинтетическую активность при умеренной температуре. На примере двух гибридов кукурузы выявлены закономерности изменения видимого фотосинтеза листьев в зависимости от освещенности и температуры. Показано, что при температуре +20º С и близкой к полной солнечной интенсивности света кукуруза способна ассимилировать со скоростью 20-25 мкмоль СО2/м2с. Зона температурного оптимума фотосинтеза находится в пределах +18...+28º С.
