Фотосинтетическая функция генотипов пшеницы с различными донорно-акцепторными отношениями при водном дефиците

Автор: Ахмедова Ф.А., Абдулбагиева С.А., Ибрагимова И.Г.
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 9 т.9, 2023 года.
Бесплатный доступ
Представлены результаты исследований изучения интенсивности фотосинтеза генотипов пшеницы при различных донорно-акцепторных отношениях. Полевые опыты проводились в оптимальном режиме полива и в условиях засухи, объектом исследований служили генотипы пшеницы: Баракатли 95, Вугар, Гарагылчыг 2 и Гырмызы бугда. Искусственное изменение донорно-акцепторных соотношений в растении в совокупности с факторами внешней среды, в том числе действием засухи, оказывает определенное влияние на интенсивность фотосинтеза. При засухе интенсивность фотосинтеза закономерно снижается в листьях. Удаление одного яруса листьев приводит к увеличению интенсивности фотосинтеза в листьях другого яруса, а при удалении половины колоса, наоборот, наблюдается снижение интенсивности фотосинтеза как в 7, так и в 8 листьях.
Пшеница, генотип, засуха, колос
Короткий адрес: https://sciup.org/14128693
IDR: 14128693 | УДК: 633.11:633.112 | DOI: 10.33619/2414-2948/94/12
Photosynthetic function of wheat genotypes with different donor-acceptor ratio in water deficiency
The article presents the results of studies of the intensity of photosynthesis of wheat genotypes at various donor-acceptor ratios. Field experiments were carried out in the optimal irrigation regime and under drought conditions, the object of research was wheat genotypes: Barakatli 95, Vugar, Garagylchyg 2 and Gyrmyzy bugda. An artificial change in the donor-acceptor ratios in the plant, in combination with environmental factors, including the effect of drought, has a certain effect on the intensity of photosynthesis. When exposed to drought, the intensity of photosynthesis naturally decreases in both leaves. The removal of one layer of leaves leads to an increase in the intensity of photosynthesis in the leaves of the other layer, and when half the ear is removed, on the contrary, a decrease in the intensity of photosynthesis is observed both in the 7th and 8th leaves.
Текст научной статьи Фотосинтетическая функция генотипов пшеницы с различными донорно-акцепторными отношениями при водном дефиците
Бюллетень науки и практики / Bulletin of Science and Practice
Процесс фотосинтеза является основным поставщиком органических веществ, необходимых для роста, развития и продуктивности растений. Продуктивность растений в основном связана с процессом фотосинтеза в листьях, и сухая биомасса образуется за счет продуктов фотосинтеза [1].
Фотосинтез протекает во всех зеленых частях растения, преимущественно в листьях, поэтому эти органы можно считать донорами ассимилятов. Транспорт и распределение продуктов фотосинтеза в онтогенезе постепенно изменяются в зависимости от степени развития листьев и акцепторная активность листа полностью заменяется донорной. С усилением донорской функции листьев в онтогенезе происходят существенные изменения и в образовании конечных продуктов фотосинтеза [2].
Роль каждого листа в общем фотосинтезе растения сначала возрастает, а затем уменьшается. У многих видов растений при удалении определенной части листовой поверхности транспорт ассимилятов из оставшихся листьев ускоряется, в это время количество крахмала в листьях снижается, а это свидетельствует о том, что происходящий в листьях фотосинтез не может компенсировать повышенный отток ассимилятов, в результате чего происходит мобилизация резервных продуктов фотосинтеза [3].
Ослабление акцепторной силы органов, потребляющих ассимиляты, вызывает торможение транспорта ассимилятов из листа или его направление к другим органам, нуждающимся в ассимилятах. Удаление части потребляющих ассимиляты органов при сохранении общей листовой поверхности у растения, иными словами торможение оттока ассимилятов из листьев, вызывает снижение интенсивности фотосинтеза в листьях [4].
Уменьшение общей листовой поверхности растения (при сохранении органов, нуждающихся в ассимилятах) стимулирует фотосинтез за счет увеличения фотосинтетической нагрузки оставшихся листьев [5].
Влияние донорно-акцепторных отношений на интенсивность фотосинтеза связано как с эпигенетическими процессами, так и с факторами внешней среды. Все фазы фотосинтеза как сложного физиологического процесса подвержены стрессу [6].
Засуха значительно снижает эффективность фотосинтеза, устьичную проводимость, площадь листьев и эффективность использования воды злаков, в том числе пшеницы [7–9].
В условиях водного стресса снижается скорость транспирации и фотосинтеза, устьичная проводимость, выделение кислорода, повышается концентрация углекислотного газа в межклеточных пространствах. Снижение количества Xl (a+b) в условиях засухи отмечено так же, как один из лимитирующих факторов фотосинтеза [10].
При различных водных режимах (80%, 60%, 45%, 35% от полевой влагоемкости) интенсивность фотосинтеза (Pn) во флаговом листе снижается с усилением почвенной засухи в фазе требования у генотипов мягкой пшеницы [11].
В фазах цветения и налива зерна водный стресс вызывал снижение скорости фотосинтеза на 30,2%, транспирации на 59,0% и устьичной проводимости на 44,2% [12]. Во время засухи при высоких температурах снижением интенсивности фотосинтеза замедляется процесс роста и снижается урожайность, при очень сильной засухе на 75%, при сильной засухе на 70%, при умеренной засухе на 65% [13].
Водный стресс зависит от различных факторов — от фазы развития и его продолжительности, глубины стресса, а также генотипа.
Материалы и методы
Полевые опыты проводились на опытном участке экспериментальной базы НИИ земледелия, расположенном на Апшеронском полуострове. Объектами исследования были взяты генотипы пшеницы отличающиеся по морфофизиологическим признакам. Опыты проводились в двух вариантах, орошаемом и неорошаемом.
В обоих вариантах у исследуемых генотипов донорно-акцепторные отношения были искусственно изменены у изучаемых сортов. Для этого у данного генотипа были взяты 10–20 образцов и удалены все листья 7 яруса, а оставлены листья 8 яруса и колос.
В другом варианте были удалены листья 8 яруса и оставлены листья 7 яруса и колос.
В другом варианте была удалена половина колоса и оставлены все листья. Для измерения интенсивности углекислотного газообмена был применен инфракрасный газоанализатор URAS-2T фирмы Хартман и Браун, Германия.
Результаты и обсуждение
Исследуемые генотипы реагируют на изменение донорно-акцепторных соотношений со дня удаления листьев, при этом наблюдается изменение интенсивности фотосинтеза по сравнению с контролем. В Таблице 1 представлены изменения интенсивности фотосинтеза при удалении листьев и половины колоса в условиях нормального водообеспечения и засухи в фазу колошения-цветения.
Как видно из Таблицы 1, максимальное значение интенсивности фотосинтеза в фазу цветения-колошения у генотипа Баракатли-95 наблюдается на 10 день удаления листьев. Разница между контрольным и опытным вариантами составляет 4,1 мг СО 2 /дм2ч в листьях 8го яруса и 4,6 мг СО 2 /дм2ч в листьях 7 яруса, которое составляют соответственно 19,3% и 22,4%. Минимальная разница между контрольным и опытным вариантами приходится на день удаления и составляет 3,6% и 3,9%. Изменение донорно-акцепторного соотношения в растении, т. е. соотношения между ассимилирующими и потребляющими органами, во всех случаях приводит к изменению интенсивности фотосинтеза. Удаление 7-го листа приводит к увеличению интенсивности фотосинтеза в 8-м листе и наоборот. Теоретически это можно объяснить тем, что при удалении определенного листа изменяется донорно-акцепторное отношения в транспортной системе целого растения и для сохранения темпа роста и развития фотосинтетическую функцию удаленного листа может брать на себя другой лист, этим обеспечивая сохранение жизненных процессов на определенном уровне [2].
Из-за воздействия засухи происходит естественное снижение интенсивности фотосинтеза в обоих листьев, однако данные, полученные в контрольном варианте мало отличаются от опытных, что может свидетельствовать о засухоустойчивости данного генотипа. На 10 день удаления листьев разница между вариантами максимальные и составляют у 8 и 7 листьев, соответственно 16,7% и 21,0%, а в тот же день удаления минимальные — 3,3% и 4,1%. Видно, что в контрольном и в опытном вариантах 7 лист больше реагирует на изменение донорно-акцепторных отношений при засухе.
Показатели изменения интенсивности фотосинтеза в листьях при удалении половины колоса у сорта Баракатли-95 представлены в Таблице 1. Если удаление листьев одного яруса приводила к увеличению интенсивности фотосинтеза других ярусов, то при удалении половины колоса, наоборот, наблюдается снижение интенсивности фотосинтеза как в 7, так и 8 ярусов. У сорта Баракатли-95 минимальное снижение интенсивности фотосинтеза наблюдается в день удаления половины колоса и составляет 4,2.
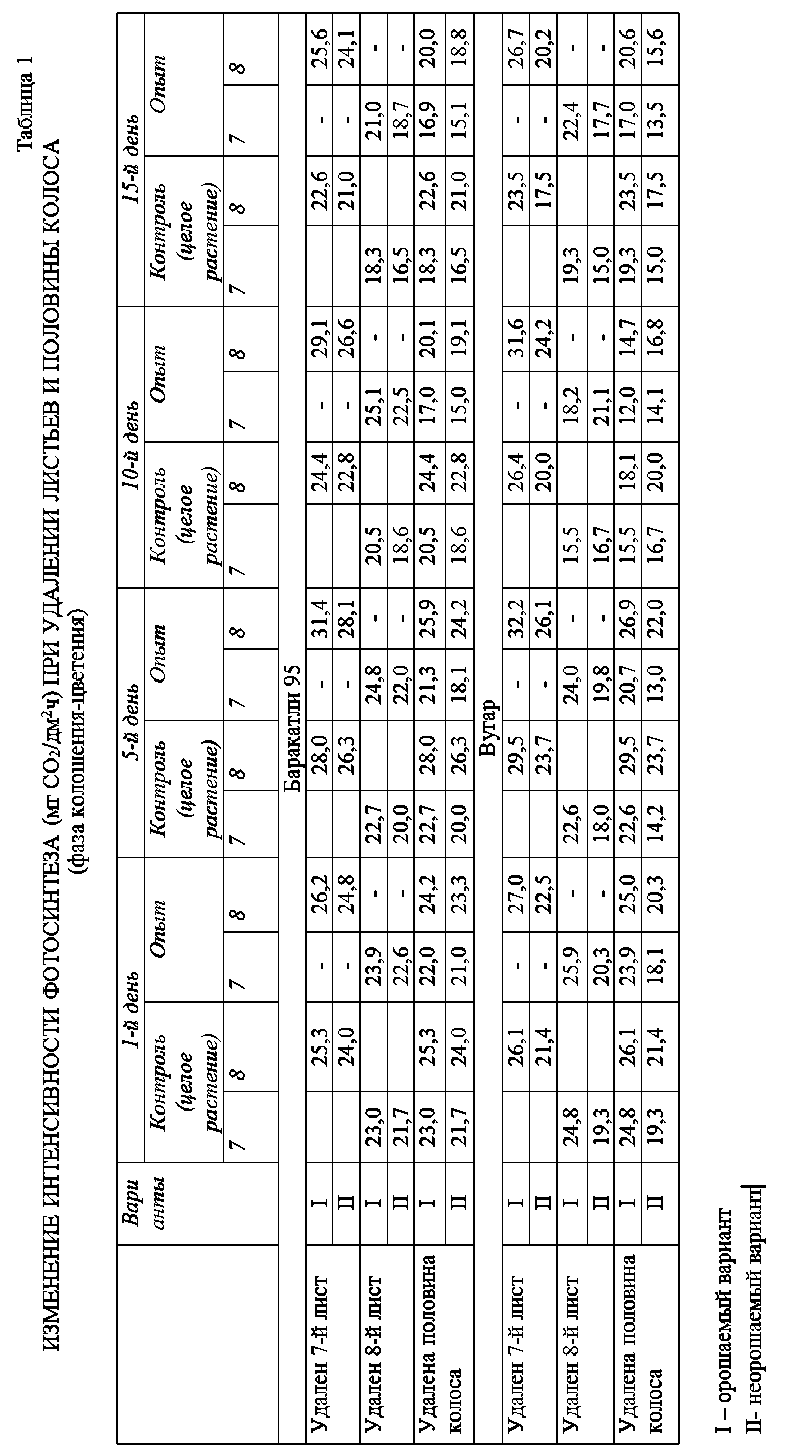
В 10 день удаления половины колоса снижение интенсивности фотосинтеза достигает максимального значения у листьев 8 яруса — 4,3 мг СО 2 /дм2ч, а у листьев 7 яруса — 3,5 мг СО 2 /дм2ч, что составляет 17,6% и 11,1%, соответственно. На 15 день снижение интенсивности фотосинтеза ослабляется и доходит примерно до уровня 5 дня удаления колоса 11,5% и 7,7%, соответственно.
По-видимому, это связано с тем, что со временем после удаления транспортная система целого растения адаптируется к новым условиям и происходит медленное восстановление прежнего уровня фотосинтеза листьев, однако не полностью, а частично. Как видно из таблицы, под влиянием засухи, у сорта Баракатли 95 в фазе колошения-цветения снижается абсолютная величина фотосинтеза, а относительная величина, т. е. процентное значение, существенно не изменяется.
У генотипа Вугар в фазе колошения-цветения интенсивность фотосинтеза листьев 7 и 8 ярусов в 10 день удаления увеличивается на 19,7% и 23,3%, а в фазе налива зерна на 23,7% и 24,6%. Однако, сравнение показателей, полученных в условиях нормального водообеспечения и засухи, показывает, что у генотипа Вугар во все фазы развития интенсивность фотосинтеза листьев 7 и 8 ярусов снижается больше, чем у других генотипов, что указывает на его относительно слабую засухоустойчивость.
Как и у других генотипов, у генотипа Вугар удаление половины колоса во всех случаях приводит к снижению интенсивности фотосинтеза в листьях. В это время максимальное снижение наблюдается так же на 10 день после удаления. У генотипа Гарагылчыг-2 максимальная величина снижения интенсивности фотосинтеза при удалении половины колоса наблюдается на 10 день, а минимальная — на 1 день (Таблица 2). В этом генотипе абсолютные и относительные величины фотосинтеза схожи.
У генотипов Гарагылчыг-2 и Гырмызы бугда в засушливом варианте интенсивность фотосинтеза снижается больше, чем у генотипов Баракатли-95 и Вугар. У этих генотипов, особенно у генотипа Гарагылчыг-2, при воздействии засухи интенсивность фотосинтеза снижается в обоих ярусах листьев примерно одинаково.
Максимальное увеличение интенсивности фотосинтеза, в отличие от низкорослых генотипов, у генотипа Гырмызы бугда наблюдается на 15 день удаления, которое составляет у листьев 8 яруса 35,7%, у листьев 7 яруса 37,2%, в условиях засухи соответственно, 27,7% и 23, 5%. Это можно объяснить тем, что у высокорослых генотиповов в связи чрезмерным ростом стебля и других генеративных органов увеличивается запрос на ассимиляты и удаление 7-го листа сокращает площадь ассимилирующих органов. Для обеспечения нормального роста и развития в целом растении и для непрерывного обеспечения ассимилятами остальных органов, листья 8 яруса в некоторой степени выполняют функцию и удаленного листа путем увеличения интенсивности фотосинтеза.
У генотипа Гырмызы бугда засуха также вызывает снижение интенсивности фотосинтеза. Изменения интенсивности фотосинтеза при удалении листьев или колоса у генотипа Гырмызы бугда в фазе колошения-цветения представлено в Таблице 2.
В листьях 7 и 8 ярусов максимальное значение интенсивности фотосинтеза наблюдалось на 10 и 15 дни удаления листьев в обоих вариантах, а минимальное — в этот же день и которые составляют, соответственно 22,0% и 20,2%, 20,5% и 21,0%, а также 2,3% и 4,3%.
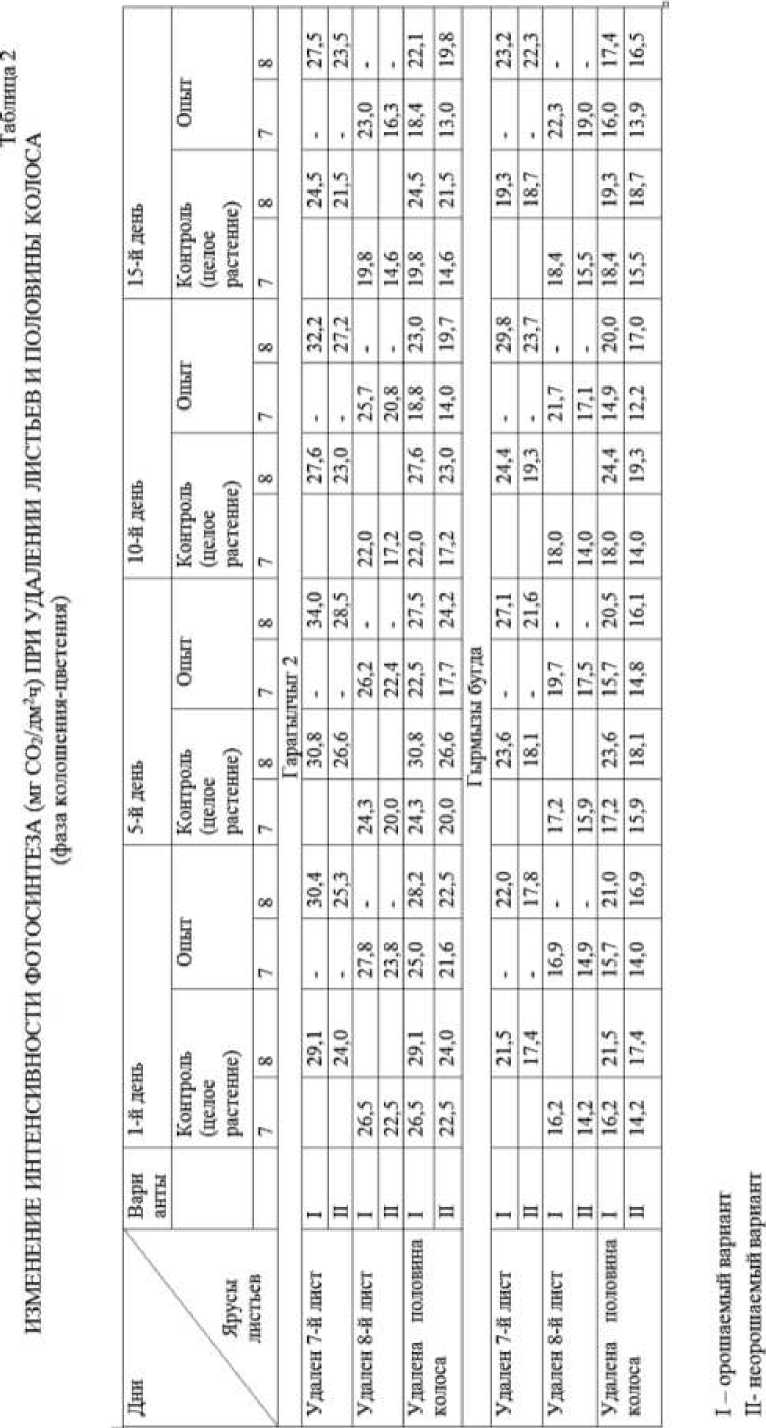
У генотипа Гырмызы бугда снижение интенсивности фотосинтеза при удалении половины колоса в фазе колошения-цветения 10 день наибольшая, однако в отличие от других генотипов у листьев этого генотипа интенсивность фотосинтеза в 15 день удаления не восстанавливается, а остается примерно на уровне 10 дня.
У генотипа Гырмызы бугда интенсивность фотосинтеза в листьях под влиянием засухи снижается больше, по сравнению с контрольным вариантом, что может характеризовать слабую адаптацию листьев к условиям засухи.
У всех исследованных генотипов снижение интенсивности фотосинтеза в листьях при удалении половины колоса, указывает на то, что снижается потребность в ассимилятах и акцепторная способность колоса. В результате эндогенной регуляции процесс фотосинтеза в листьях несколько замедляется [14].
Таким образом, один из путей адаптации фотосинтетической деятельности растений пшеницы при изменении донорно-акцепторных отношений в условиях засухи может быть тот факт, что фотосинтетическая деятельность листьев непосредственно связана с потребностью колоса в ассимилятах и контролируется его аттрагирующей способностью.
Список литературы Фотосинтетическая функция генотипов пшеницы с различными донорно-акцепторными отношениями при водном дефиците
- Ashraf M., Harris P. J. C. Photosynthesis under stressful environments: An overview // Photosynthetica. 2013. V. 51. P. 163-190. https://doi.org/10.1007/s11099-013-0021-6
- Aliyev J. A. Photosynthesis, photorespiration and productivity of wheat and soybean genotypes // Physiologia Plantarum. 2012. V. 145. №3. P. 369-383. https://doi.org/10.1111/j.1399-3054.2012.01613.x
- Мокроносов А. Т. Интеграция функций роста и фотосинтеза // Физиология растений. 1983. Т. 30. №5. С. 868-880.
- Hussain M., Waqas-ul-Haq M., Farooq S., Jabran K., Farroq M. The impact of seed priming and row spacing on the productivity of different cultivars of irrigated wheat under early season drought // Experimental Agriculture. 2016. V. 52. №3. P. 477-490. https://doi.org/10.1017/S0014479716000053
- Чиков В. И. Фотосинтез и транспорт ассимилятов. М.: Наука, 1987. 185 с.
- Bogale A., Tesfaye K., Geleto T. Morphological and physiological attributes associated to drought tolerance of Ethiopian durum wheat genotypes under water deficit condition // Journal of biodiversity and environmental sciences. 2011. V. 1. №2. P. 22-36.
- Changhai S., Baodi D., Yunzhou Q., Yuxin L., Lei S., Mengyu L., Haipei L. Physiological regulation of high transpiration efficiency in winter wheat under drought conditions // Plant, Soil and Environment. 2010. V. 56. №7. P. 340-347. https://doi.org/10.17221/220/2009-PSE
- Lamaoui M., Jemo M., Datla R., Bekkaoui F. Heat and drought stresses in crops and approaches for their mitigation // Frontiers in chemistry. 2018. V. 6. P. 26. https://doi.org/10.3389/fchem.2018.00026
- Zandalinas S. I., Mittler R., Balfagón D., Arbona V., Gómez‐Cadenas A. Plant adaptations to the combination of drought and high temperatures // Physiologia plantarum. 2018. V. 162. №1. P. 2-12. https://doi.org/10.1111/ppl.12540
- Hall A. J., Brady C. J. Assimilate source-sink relationships in Capsicum annuum L. II. Effects of fruiting and defloration on the photosynthetic capacity and senescence of the leaves // Functional Plant Biology. 1977. V. 4. №5. P. 771-783. https://doi.org/10.1071/PP9770771
- Shan C., Tang J., Yang W., Zhao X., Ren X., Li Y. Comparison of photosynthetic characteristics of four wheat (Triticum aestivum L.) genotypes during jointing stage under drought stress // African Journal of Agricultural Research. 2012. V. 7. №8. P. 1289-1295.
- Farooq M., Hussain M., Ul-Allah S., Siddique K. H. Physiological and agronomic approaches for improving water-use efficiency in crop plants // Agricultural Water Management. 2019. V. 219. P. 95-108. https://doi.org/10.1016/j.agwat.2019.04.010
- Калинин Н. И. Влияние экстремальных гидротермических условий на темпы развития яровой пшеницы // Бюллетень ВНИИ растениеводства. 1982. №116. С. 62-67.
- Моргун В. В., Киризий Д. А., Шадчина Т. М. Экофизиологические и генетические аспекты адаптации культурных растений к глобальным изменениям климата // Физиология и биохимия культурных растений. 2010.
