Фотосинтетические пигменты фитопланктона высоких широт в условиях полярной ночи

Автор: Водопьянова Вероника Владимировна, Макаревич Павел Робертович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Океанология
Статья в выпуске: 1-2 т.19, 2016 года.
Бесплатный доступ
Впервые в Баренцевом море в период отсутствия фотосинтетически активной радиации исследован пигментный состав и вертикальное распределение хлорофилла фитопланктона. Приведена оценка функционального состояния фитопланктонного сообщества в условиях полярной ночи. Выявлено растущее влияние тепловодной атлантической флоры на фитопланктонное сообщество Баренцева моря.
Хлорофилл, фитопланктон, фотосинтетические пигменты
Короткий адрес: https://sciup.org/14294888
IDR: 14294888 | УДК: 574.583 | DOI: 10.21443/1560-9278-2016-1/2-249-257
Photosynthetic pigments in phytoplankton living in high latitudes under conditions of the polar night
Pigment composition and the vertical distribution of phytoplankton chlorophyll have been studied for the first time in the Barents Sea during the period when there is no photosynthetically active radiation. The paper describes the functional status of the Barents Sea phytoplankton community under conditions of the polar night. Studies of the Barents Sea phytoplankton during the polar night have identified an increasing influence of the warm-water Atlantic flora on the Barents Sea phytoplankton community.
Текст научной статьи Фотосинтетические пигменты фитопланктона высоких широт в условиях полярной ночи
Традиционно считается, что экосистема пелагиали северных морей функционирует в жестких экологических условиях. В период полярной ночи (декабрь – январь) количество солнечной радиации в этих водоемах составляет 0,002 (кал/см 2 )/мин, тогда как минимальная интенсивность света, необходимая для роста практически всех микроводорослей, должна достигать порядка 0,01 (кал/см 2 )/мин [1]. Содержание в фитопланктоне основного пигмента фотосинтеза – хлорофилла а – считается универсальным экологофизиологическим показателем, который отражает обилие и фотосинтетическую активность альгоценозов. Адаптация водорослей к низкому уровню освещенности может выражаться в составе и активности дополнительных пигментов фотосинтетического аппарата [2]. Дополнительные пигментные характеристики планктона характеризуют функциональное состояние и протекающие в фитопланктонном сообществе продукционные процессы. Наибольший интерес для функциональной оценки фитопланктона в осеннезимний период представляют пигментный индекс и соотношение суммарной концентрации каротиноидов к хлорофиллу а [3–7].
Исследования видового состава, биомассы и численности фитопланктона Баренцева моря ведутся в течение продолжительного времени [8–10]. Характеристика пигментного состава фитопланктона арктических морей представлена в основном сведениями о концентрации хлорофилла а , полученными в период активной вегетации микроводорослей. При этом не учитывается фотосинтетическая активность альгоценозов в зимний период и не раскрываются данные о содержании дополнительных пигментов фотосинтетического аппарата микрофитопланктона [11–14]. Для Баренцева моря наиболее полно представлена информация о концентрации фотосинтетических пигментов альгосообществ пелагиали прибрежья в период весенне-летней и летне-осенней сукцессии [7; 15].
В последние годы большой интерес исследователей вызывают возможности спутникового (дистанционного) зондирования концентраций пигмента. Полученные данные, однако, необходимо сравнивать с данными натурных измерений хлорофилла [16; 17]. Для акватории Баренцева моря получение спутниковых данных о распределении полей хлорофилла затруднено из-за высокой облачности, особенно в осенне-зимний период [18]. Получаемые на основе биооптических характеристик поверхности моря региональные карты распределения хлорофилла свидетельствуют, что самые высокие его концентрации наблюдаются летом к югу от полярного фронта [17]. Тем не менее необходим мониторинг концентраций данного пигмента в толще воды, так как глубинный максимум хлорофилла может вносить существенный вклад в первичную продукцию Мирового океана, но, как правило, не учитывается при использовании спутниковых данных и данных поверхностных измерений in situ [19].
Целью настоящей работы являлось изучение хлорофилла и дополнительных пигментов фотосинтетического аппарата арктического фитопланктона Баренцева моря в переходный период между осенним и зимним гидрологическими сезонами в условиях недостаточной освещенности.
Материалы и методы
Фотосинтетические пигменты микрофитопланктонного сообщества Баренцева моря исследовали в ходе рейса НИС "Дальние Зеленцы" с 10 по 25 ноября 2013 г. Научные изыскания проводились на стандартном разрезе "Кольский меридиан" (станции 1, 3, 7, 11, 15, 17, 23) и участке западнее от него (станция 41) (рис. 1). Продолжительность светового времени в ходе выполнения наблюдений убывала от 4.5 часов на широте 70° с. ш. до 0.0 часов на широте 74° с. ш. и севернее, таким образом, исследования на станциях 15, 17, 23 и 41 выполнялись уже в условиях наступившей полярной ночи.
Рис. 1. Схема расположения станций на разрезе "Кольский меридиан", НИС "Дальние Зеленцы", ноябрь 2013 г.
В соответствии с рекомендациями ГОСТа 1 было отобрано 45 проб морской воды для дальнейшего определения концентрации фотосинтетических пигментов: хлорофилла а ( С ха ), b ( С b ), с ( С с1 + с2 ), феофитина а ( С фа ), каротиноидов ( С k ) и пигментного индекса сообщества ( I 430/664 ). Пробы отбирали с шести горизонтов: поверхностный слой, глубины 10, 25, 50, 100 м и придонный слой (глубины от 150 до 310 м). Объем проб достигал 5 л. С момента отбора проб на пигментный состав и до процесса фильтрации воды проходило не более часа. Фильтрацию осуществляли под вакуумом через мембранные фильтры Владипор МФАС-ОС-4 (диаметр пор 0.4 мкм) продолжительность фильтрования не превышала 60 мин. Фильтры, сложенные пополам осадком внутрь, хранили в морозильной камере до обработки в стационарных условиях не более 1 месяца.
В лаборатории фильтры гомогенизировали для разрушения клеток фитопланктона и экстрагировали пигменты. Экстракцию проводили 90%-м ацетоном в течение 1 часа. Для удаления светорассеивающей взвеси полученный экстракт центрифугировали на центрифуге Опн-8 20 мин при 8 000 об./мин. Спектрофотометрирование очищенного экстракта осуществляли дважды (до и после подкисления экстракта раствором соляной кислоты) на UV-Visible спектрофотометре Nicolett Evolution 500 "Spectronic Unicam", использовали кюветы, имеющие рабочую длину 1 см. Концентрацию хлорофилла а и других пигментов (мг/м3) вычисляли по стандартным формулам, приведенным в ГОСТ 17.1.4.02-90.
Результаты и обсуждение
Концентрация хлорофилла а в толще воды на большинстве станций изменялась в незначительном для применяемой методики диапазоне (0.05–0.21 мг/м 3 ), за исключением станции 23, где были зарегистрированы самые высокие концентрации исследуемого пигмента и придонный максимум содержания хлорофилла (0.50 мг/м 3 ) (рис. 2).
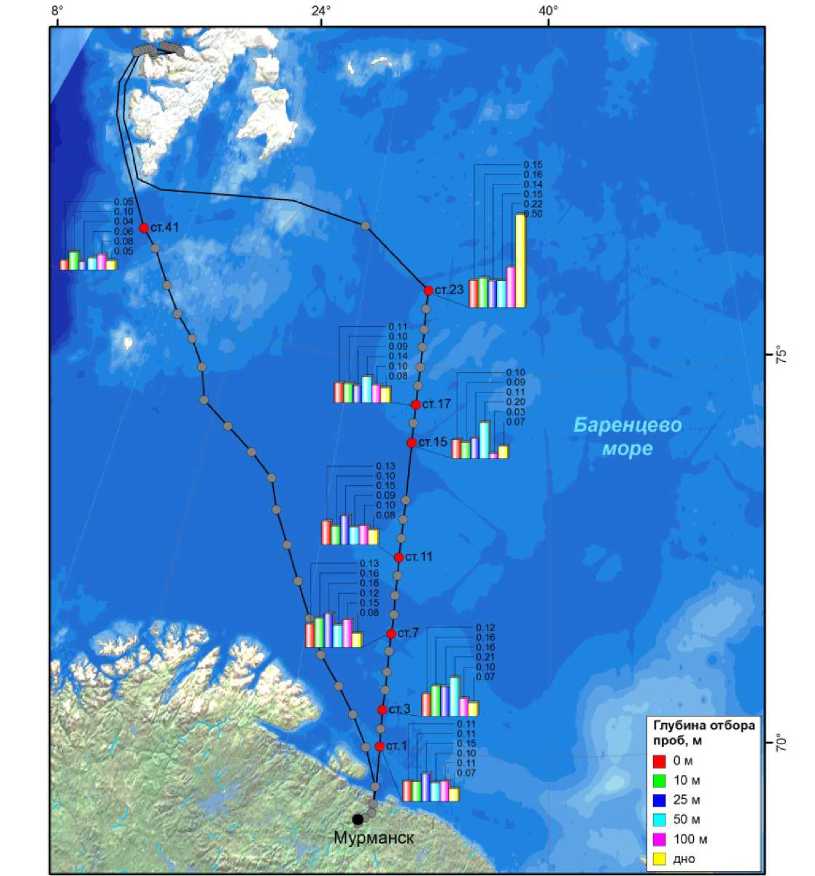
Рис. 2. Концентрация хлорофилла а , мг/м 3 , на станциях разреза "Кольский меридиан", ноябрь 2013 г.
Равномерное вертикальное распределение хлорофилла а на всех станциях, несомненно, связано с отсутствием стратификации водных масс в период проведения исследований. Условия освещенности в период проведения научных изысканий были характерными для осенне-зимнего гидрологического периода, часть станций (выше 72° 34') находилась в условиях полярной ночи.
Анализ полученных с помощью СТД-зонда (SEACAT SBE 19plus) термохалинных характеристик показал, что южный участок разреза (станции 1, 3) находился в зоне влияния Мурманского прибрежного течения, а центральный (станции 7, 11) и северный (станции 15, 17, 23) участки – в зоне воздействия атлантических вод (в пределах Мурманского и Нордкапского течений соответственно). Прибрежные воды являлись более прогретыми и менее солеными по сравнению с атлантическими водами (табл. 1). На исследуемой акватории переход к зимнему гидрологическому режиму сопровождался разрушением сезонного термоклина, в результате чего в верхнем 50-метровом слое наблюдалась гомотермия, далее температура плавно понижалась с глубиной.
Диапазоны значения температуры ( Т , °С) и солености ( S , ‰) воды на участках разреза "Кольский меридиан", ноябрь 2013 г.
Таблица 1
|
Участок |
Южный (станции 1, 3) 69°30'–70°30' с. ш. |
Центральный (станции 7, 11), 71°30'–72°30' с. ш. |
Северный (станции 15, 17, 23), 74°00'–76°00' с. ш. |
|||
|
Слой |
0–50 м |
100 м – дно |
0–50 м |
100 м – дно |
0–50 м |
100 м – дно |
|
Т , °С |
6.03–7.28 |
3.90–6.62 |
5.23–7.00 |
1.62–5.98 |
3.72–5.08 |
0.11–4.79 |
|
S , ‰ |
34.31–34.42 |
34.57–34.81 |
34.39–34.94 |
34.72–35.07 |
35.05–35.07 |
34.98–35.09 |
Глубинный максимум хлорофилла а , зафиксированный на единственной станции (0.50 мг/м 3 ), указывает на активное развитие микроводорослей. В последнее время отмечается повышение температуры вод арктических морей [20], что может привести к изменению таксономического состава фитопланктонного сообщества [10].
Согласно литературным данным [7; 12; 15] концентрации хлорофилла а выше 0.10 мг/м 3 являются высокими для района исследований в осеннее-зимний период. Кроме того, столь высокие концентрации не соответствуют биомассе и численности "сетного" микрофитопланктона. В период наших исследований общая численность микроводорослей размерной группы более 20 мкм составляла 1×10 1 кл./л. По составу доминантов и уровню обилия сообщество фитопланктона являлось типичным для южной части Баренцева моря в осенне-зимний период, были отмечены известные формы динофлагеллят – индикаторы атлантических водных масс. Кроме того, при изучении видового состава микрофитопланктонного сообщества впервые для изучаемой акватории зафиксированы тепловодные бореальные виды динофлагеллят, ранее не встречающиеся в Баренцевом море: Amphidoma caudata Halldal, Dinophysis hastata Stein, Dinophysis nasuta Parke & Dixon, Dinophysis ovata Claparéde & Lachmann, Heterodinium milneri Kofoid, Mesoporos perforatus Lillick, Oxytoxum milneri Murray & Whitting, Podolampas palmipes Stein, Protoperidinium brochii Balech, Pyrophacus horologicum Stein.
Известно, что для олиготрофных вод Атлантики максимальный вклад (до 80 %) в первичную продукцию и содержание хлорофилла а в фитопланктонном сообществе вносит пикопланктон [21; 22], для эвтрофных вод – ультрананопланктон [23]. Доля хлорофилла, соответствующая размерной группе фитопланктеров 0.45–0.85 мкм, может достигать 49 % [22]. В арктических экосистемах на долю малых клеток может приходиться до 84 % хлорофилла и до 49 % первичной продукции [24]. Как показали наши исследования, видовой состав микрофитопланктонного сообщества Баренцева моря в осенне-зимний период формировался за счет атлантической тепловодной флоры. Предположительно, повышенные концентрации хлорофилла а в период изысканий могут указывать на то, что микрофитопланктонное сообщество Баренцева моря представлено не только фитопланктоном размерной фракции более 20 мкм, но и видами более мелких размерных фракций (пикопланктон, ультрананопланктон), характерных для атлантических вод в больших количествах и вносящих основной вклад в первичную продукцию.
Доля хлорофилла а в сообществе фотосинтетиков поверхностного слоя в период исследований не превышала (43.44 ± 5.26) % (в % от суммы всех хлорофиллов), хлорофиллов b и с 1 + с 2 (24.64 ± 4.11) % и (31.94 ± 5.53) % соответственно. Для всех слоев содержание хлорофилла а в фитопланктоне составило (39.27 ± 8.45) %, хлорофилла b (25.36 ± 5.68) %, хлорофиллов с 1 + с 2 (35.37 ± 6.16) %. Такую картину с незначительным преобладанием доли хлорофилла а над другими пигментами мы наблюдали в осеннезимний сукцессионный период в фитопланктоне прибрежья Баренцева моря, в то время как в период активной вегетации микроводорослей в весенне-летний период, доля хлорофилла а достигала 70 % [7]. Таким образом, в условиях полярной ночи, при сниженной продукционной активности микрофитопланктона процессы деструкции не преобладают над автотрофными, адаптивные механизмы фотосинтетиков выражаются в увеличении доли дополнительных фотосинтетических пигментов.
Для оценки физиологического состояния фитопланктона и продукционных процессов, происходящих в сообществе фотосинтетиков, нами традиционно используются следующие характеристики: пигментный индекс ( I 430/664 ), отношение суммарной концентрации хлорофиллов с 1 +с 2 к концентрации хлорофилла а ( С с / C ха ), отношение суммарной концентрации каротиноидов к концентрации хлорофилла а ( С k / C ха ) и доля в сообществе феофитина а – продукта распада хлорофилла а (табл. 2).
Таблица 2
Оценка физиологического состояния фитопланктона и продукционных процессов, происходящих в сообществе фотосинтетиков, выполненная в ходе исследований на станциях разреза "Кольский меридиан", ноябрь 2013 г.
|
Номер станции |
2 |
3 |
7 |
11 |
15 |
17 |
23 |
41 |
Среднее по горизонту |
|
|
I430/664 |
||||||||||
|
н о к Си о 1-н |
0 м |
3.12 |
3.57 |
2.23 |
3.10 |
4.01 |
4.03 |
2.78 |
6.89 |
3.72 |
|
10 м |
4.28 |
3.10 |
2.00 |
3.91 |
4.92 |
4.54 |
3.52 |
3.89 |
3.77 |
|
|
25 м |
3.20 |
3.37 |
2.46 |
3.61 |
4.62 |
4.79 |
4.01 |
7.48 |
4.19 |
|
|
50 м |
3.12 |
2.11 |
3.00 |
3.40 |
3.05 |
4.40 |
4.93 |
5.70 |
3.71 |
|
|
100 м |
3.40 |
4.57 |
2.33 |
4.20 |
8.15 |
3.81 |
5.70 |
5.58 |
4.72 |
|
|
Дно |
3.65 |
4.53 |
3.42 |
3.17 |
4.37 |
3.93 |
7.90 |
7.50 |
4.81 |
|
|
Схс/Сха |
||||||||||
|
н о к Си о 1-н |
0 м |
0.64 |
0.84 |
0.61 |
0.50 |
0.65 |
0.71 |
0.84 |
1.29 |
0.76 |
|
10 м |
0.75 |
1.05 |
1.08 |
0.39 |
0.85 |
0.62 |
1.04 |
1.33 |
0.89 |
|
|
25 м |
0.40 |
0.91 |
0.97 |
0.53 |
0.67 |
0.74 |
1.02 |
0.70 |
0.74 |
|
|
50 м |
0.70 |
0.76 |
0.74 |
1.38 |
1.14 |
0.77 |
1.00 |
1.09 |
0.95 |
|
|
100 м |
0.64 |
1.51 |
1.12 |
0.46 |
1.20 |
1.02 |
1.28 |
1.12 |
1.04 |
|
|
Дно |
0.86 |
1.66 |
1.54 |
1.38 |
1.08 |
1.45 |
1.97 |
1.62 |
1.45 |
|
|
Сk/Cха |
||||||||||
|
н о к Си о 1-н |
0 м |
2.18 |
1.72 |
1.26 |
1.48 |
1.84 |
1.89 |
1.39 |
3.29 |
1.88 |
|
10 м |
1.96 |
1.64 |
1.42 |
1.70 |
2.35 |
2.15 |
1.75 |
2.15 |
1.89 |
|
|
25 м |
1.00 |
1.67 |
1.53 |
1.65 |
2.13 |
2.16 |
2.04 |
3.28 |
1.93 |
|
|
50 м |
2.00 |
1.26 |
1.65 |
2.12 |
1.68 |
2.13 |
2.28 |
2.62 |
1.97 |
|
|
100 м |
2.00 |
2.31 |
1.59 |
1.81 |
3.79 |
1.85 |
2.60 |
3.15 |
2.39 |
|
|
Дно |
2.86 |
2.34 |
2.16 |
1.59 |
2.31 |
2.30 |
1.30 |
4.01 |
2.23 |
|
|
Феофитин а, % |
||||||||||
|
н о к Си о 1-н |
0 м |
39 |
50 |
62 |
57 |
63 |
44 |
34 |
24 |
47 |
|
10 м |
53 |
51 |
54 |
49 |
35 |
38 |
54 |
73 |
51 |
|
|
25 м |
40 |
60 |
53 |
27 |
42 |
52 |
67 |
64 |
51 |
|
|
50 м |
44 |
52 |
36 |
66 |
53 |
40 |
70 |
55 |
52 |
|
|
100 м |
39 |
45 |
51 |
20 |
55 |
54 |
64 |
49 |
47 |
|
|
Дно |
50 |
57 |
56 |
61 |
58 |
57 |
63 |
53 |
57 |
|
Пигментный индекс микрофитопланктонного сообщества на большинстве станций находился в диапазоне 2.00 > I 430/664 < 4.92 (табл. 2). Однако на станциях, расположенных в самых северных точках и в придонном горизонте, наблюдалось существенное увеличение этого показателя (5.58–8.15) (табл. 2). Ранее нам не доводилось регистрировать столь высокие значения пигментного индекса в прибрежье или открытых водах Баренцева моря, максимально в зимний период (по результатам многолетнего мониторинга) I 430/664 достигал 5.14 [7], а для фитопланктонного сообщества Атлантики пигментный индекс может принимать значения до 9.0 [25]. Значения пигментного индекса I 430/664 < 4.00 (табл. 2) следует рассматривать как индикатор пограничного состояния фитопланктонного сообщества между стадией активной вегетации и стадией затухания его сукцессионного цикла; повышение этого показателя I 430/664 > 4.00 (табл. 2) свидетельствует о низкой фотосинтетической активности микрофитопланктона [7].
Отношение суммарной концентрации хлорофиллов с 1 + с 2 к концентрации хлорофилла а ( С хс /С ха ) увеличивается при затухании развития планктонных водорослей и снижении ими фотосинтетической активности [25]. Нами отмечено, что средние значения соотношения С хс /С ха увеличиваются с глубиной от 0.76 до 1.45 (табл. 2), а также по направлению к самым северным точкам – для всех станций (табл. 2). В фотическом слое, за исключением станций 23 и 41, этот показатель не превышал 1 (табл. 1), что характеризует сообщество как находящееся в состоянии функциональной (фотосинтетической) активности. Известно, что при старении фитопланктона соотношение С хс /С ха может принимать значения 3.6–9.6, в активно развивающемся сообществе этот показатель варьирует от 0.0 до 0.8 [25].
Отношение суммарной концентрации каротиноидов к концентрации хлорофилла а (Сk/Cха) характеризует физиологическое состояние фитопланктонного сообщества. Повышенные значения Сk/Cха свидетельствуют о старении сообщества и угнетенном состоянии микроводорослей [3–5; 26]. Значения Сk/Cха <2 свидетельствуют о функциональной активности фитоценоза Баренцева моря, повышение этого соотношения (Сk/Cха >2) можно рассматривать как сигнал о старении сообщества [7]. В период нашего исследования значения Сk/Cха изменялись в диапазоне 1.00 > Сk/Cха <4.01 (табл. 2), наибольшие показатели отмечены на самой северной станции, однако они далеки от характерных зимних величин (до 9.28) [7]. Для средних значений анализируемого параметра увеличение Сk/Cха отмечалось по направлению от поверхности к придонному слою (табл. 2). Можно заключить, что сообщество планктонных фотосинтетиков Баренцева моря в период проведения исследований находилось в функционально активном состоянии. Ожидаемо менее активно автотрофные процессы протекали в придонных слоях и на станции, расположенной в самой северной точке.
Доля феофитина а в толще воды изменялась в диапазоне от 20 до 70 % (средние показатели 47–57 %) (табл. 2). Содержание в фитопланктонном сообществе Баренцева моря в осенне-зимний период дериватов хлорофилла менее 70 % рассматривается как невысокое [7; 27] и свидетельствует о доминирующей роли активных форм фотосинтетических пигментов в продукционных процессах. Подобный факт подтверждают литературные данные, свидетельствующие об обнаружении функционально активного хлорофилла в клетках микрофитопланктона при минимальном уровне солнечной радиации во время полярной ночи и активной вегетации сообщества в подледных условиях [1; 28–31].
В пигментном составе фитопланктонного сообщества Баренцева моря в период полярной ночи выявлено увеличение доли дополнительных фотосинтетических пигментов (хлорофиллов b и с 1 + с 2 ) и лишь незначительное преобладание хлорофилла а . Согласно дополнительным пигментным характеристикам автотрофные процессы в микрофитопланктоне доминировали над деструкционными, сообщество планктонных фотосинтетиков находилось в функционально активном состоянии.
Заключение
Вертикальное распределение хлорофилла а в водной толще на большинстве станций было равномерным, что вероятно связано с условиями недостаточной освещенности и отсутствием стратификации водных масс в период проведения исследований. Концентрации хлорофилла а были высокими для периода научных изысканий и не соответствовали биомассе и численности фитопланктона размерной группы более 20 мкм, что указывает на присутствие в пробах более мелких размерных групп фитопланктеров, характерных для атлантических вод. Соотношения дополнительных фотосинтетических пигментов микрофитопланктона и продуктов их распада характеризуют фитопланктонное сообщество Баренцева моря в период полярной ночи как находящееся в функционально активном состоянии, и хотя продукционная деятельность фитопланктона невелика, она не прекращается полностью даже в условиях полярной ночи, т. е. в условиях недостаточной освещенности. Адаптивные механизмы сообщества фотосинтетиков к условиям полярной ночи выражаются в увеличении доли дополнительных фотосинтетических пигментов (хлорофиллов b и с 1 + с 2 ) и снижении доли основного пигмента фотосинтеза – хлорофилла а .
Таким образом, полученные результаты свидетельствуют о сохранении пелагическим сообществом микроводорослей незамерзающей части Баренцева моря фотосинтетической активности в период полярной ночи, что в итоге оказывается одним из факторов, определяющих высокую биологическую продуктивность экосистем в Западном арктическом бассейне.
Выражаем благодарность сотруднику ММБИ Калинке О. П., оказавшей помощь в отборе проб морской воды в сложных полевых условиях.
Список литературы Фотосинтетические пигменты фитопланктона высоких широт в условиях полярной ночи
- Макаревич П. Р., Ларионов В. В., Дружкова Е. И. Динамика фитопланктона в эстуарных областях северных морей в период полярной ночи//Альгология. 2004. Т. 14, № 2. С. 137-142.
- Злобин В. С. Первичная продукция и культивирование морского фитопланктона. М.: Пищевая промышленность, 1976. 247 с.
- Humphrey G. F. Seasonal variations in plankton pigments in waters off Sydney, Austral//J. Marine Freshwater Res. 1963. V. 14, N 1. P. 24-36.
- Margaleff R. Valeur indicatrice de la composition des pigments du phytoplankton sur la productivite, composition taxonomique et proprietes dynamiques des populations//Comm. Int. Explor. Sci. Mer. Medit., Rapp. Proc. Verb. 1960. V. 15, N 2. P. 277-281.
- Parsons T. R., Stevens K., Strickland J. D. H. On the chemical composition of eleven species of marine phytoplankters//J. Fish. Res. Bd. Can. 1961. V. 18, N 6. P. 1001-1016.
- Бульон В. В. Содержание феопигментов в планктоне (обзор)//Гидробиол. журн. 1978. Т. 14, № 3. С. 62-69.
- Макаревич П. Р., Водопьянова В. В., Олейник А. А. Фитоценозы пелагиали Кольского залива. Структура и функциональные характеристики/отв. ред. С. Л. Дженюк. Ростов н/Д.: Изд-во ЮНЦ РАН, 2015. 192 с.
- Макаревич П. Р. Структура и функционирование планктонных альгоценозов эстуарных экосистем шельфовых морей: автореф. дис. … д-ра биол. наук. Мурманск, 2004. 45 с.
- Макаревич П. Р., Дружкова Е. И. Сезонные циклические процессы в прибрежных планктонных альгоценозах северных морей. Ростов н/Д: Изд-во ЮНЦ РАН, 2010. 280 с.
- Олейник А. А. Новые динофлагеллаты (Dinophyceae) планктона Баренцева моря//Ботан. журн. 2014. Т. 99, № 7. С. 28-37.
- Sakshang E. Food webs and primary production in the Barents area//Proc. NIPR Symp. Polar. Biol. 1991. N 8. P. 1-8.
- Планктон морей Западной Арктики/отв. ред. Г. Г. Матишов. Апатиты: Изд-во КНЦ РАН, 1997. 352 с.
- Ведерников В. И., Гагарин В. И. Первичная продукция и хлорофилл в Баренцевом море в сентябре -октябре 1997 г.//Океанология. 1998. Т. 38, № 5. С. 710-718.
- Вирусы в пелагиали Баренцева моря/М. П. Венгер, Т. И. Широколобова, П. Р. Макаревич, В. В. Водопьянова//Докл. акад. наук. 2012. Т. 446, № 3. С. 345-349.
- Кузнецов Л. Л., Шошина Е. В. Фитоценозы Баренцева моря (физиологические и структурные характеристики). Апатиты: Изд-во КНЦ РАН, 2003. С. 48-75.
- Ветров А. А., Романкевич Е. А. Межгодовая изменчивость первичной продукции и потоков органического углерода в арктических морях России//Океанология. 2008. Т. 48, № 3. С. 371-380.
- Копелевич О. В., Буренков В. И., Шеберстов С. В. Разработка и использование региональных алгоритмов для расчета биооптических характеристик морей России по данным спутниковых сканеров цвета//Современные проблемы дистанционного зондирования Земли из космоса. 2006. Т. II. С. 99-105.
- Водопьянова В. В. Содержание хлорофилла в фитопланктоне Баренцева моря в осенний период//Гидробиологические и экосистемные исследования морей Европейского Севера: мат. ХХVIII конф. молодых ученых Мурман. морского биолог. ин-та, посвящ. 100-летию со дня рождения М. М. Камшилова (г. Мурманск, май 2010 г.). Мурманск, 2010. С. 35-40.
- Parameterization of vertical chlorophyll a in the Arctic Ocean: Impact of the subsurface chlorophyll maximum on regional, seasonal and annual primary production estimates/M. Ardyna, M. Babin, M. Gosselin, E. Devred, S. B´elanger, A. Matsuoka, J.-E´. Tremblay//Biogeosciences Discuss. 2013. 10 (1345-1399). URL: www.biogeosciences-discuss.net/10/1345/2013/. Doi: DOI: 10.5194/bgd-10-1345-2013
- Fifty years of meteo-glaciological change in Toll Glacier, Bennett Island, De Long Islands, Siberian Arctic/K. Konya, T. Kadota, H. Yabuki, T. Ohata//Polar Science. 2014. V. 8, N 2. P. 86-95.
- Platt T., Subba Rao D. V., Irwin B. Photosynthesis of picoplankton in the oligotrophic ocean//Nature. 1983. V. 301, N 5902. P. 702-704.
- Крупаткина Д. К., Лопухин А. С., Каменир Ю. Г. Размерное распределение фитопланктона Тропической Атлантики и его влияние на оценку первичной продукции и хлорофилла а в водах разной трофности//Океанология. 1987. Т. ХХVII. Вып. 3. С. 470-474.
- Стельмах Л. В. Вклад пикопланктона в первичную продукцию и содержание хлорофилла а в эвтрофных водах на примере Севастопольской бухты//Океанология. 1988. Т. 28. Вып. 1. С. 127-132.
- Wassmann P., Carroll J., Bellerby R. G. J. Carbon flux and ecosystem feedback in the northern Barents Sea in an era of climate change: An introduction//Deep-Sea Research. 2008. V. 55, N 2. P. 2143-2153.
- Балоде М. Я. Результаты пигментного анализа фитопланктона Балтийского моря в сентябре -октябре 1980 г.//Планктон Балтийского моря/отв. ред. Г. П. Андрушайтис. Рига, 1990. С. 92-101.
- Travers M. Recherches sur le phytoplancton du Golfe de Marseille. II. Etude quantitative des populations phytoplanctoniques du Golfe de Marseille (I)//Resuelt Trav. Station Mar. D'Endoume. 1962. V. 41, N 26. P. 70-139.
- Мордасова Н. В., Федосов М. В. О содержании хлорофилла и феофитина в воде и льдах Баренцева моря//Тр. ВНИРО. 1972. Т. 75. Вып. 1. С. 80-87.
- Быков О. Д. Фотосинтез и темновая фиксация СО2 литоральных макроводорослей Баренцева моря в условиях полярной ночи//Ботан. журн. 2003. Т. 88, № 12. С. 68-73.
- High-resolution observations of aggregate flux during a sub-polar North Atlantic spring bloom/N. Briggs, M. J. Perry, I. Cetinic, C. Lee, E. D’Asaro, A. M. Gray, E. Rehm//Deep-Sea Research. 2011. V. 58, N 1. P. 1031-1039.
- Laney S. R., Sosik H. M. Phytoplankton assemblage structure in and around a massive under-ice bloom in the Chukchi Sea//Deep-Sea Research. 2014. V. 105, N 2. P. 30-41.
- Lowry K. E., Dijken G. L., Arrigo K. R. Evidence of under-ice phytoplankton blooms in the Chukchi Sea from 1998 to 2012//Deep-Sea Research. 2014. V. 105, N 2. P. 105-117.
