Фрагменты гомологии эндогенных ретровирусов в геномах растений и животных

Автор: Елькина М.А., Глазко В.И.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Молекулярная биология, вирусология
Статья в выпуске: 5 т.49, 2014 года.
Бесплатный доступ
Использование последовательностей мобильных генетических элементов в целях маркирования полиморфизма участков их инсерций может оказаться наиболее эффективным подходом при выявлении специфических особенностей генофондов для различных групп организмов, контроля их динамики, что особенно важно в работе с сельскохозяйственными видами. В царстве растений широко распространен Sireviruses ( Pseudoviridae ) - одни из древнейших LTR-ретротранспозонов. Наиболее крупный и подробно изученный ретротранспозон - SIRE-1. Его анализ показывает, что инсерции указанного ретроэлемента (в частности, в геном кукурузы) произошли недавно. Мы изучили полиморфизм фрагментов ДНК, фланкированных участками LTR-ретротранспозона SIRE-1 (IRAP-PCR маркеры), в геномах у различных таксономических групп. Объектами исследования были однодольные растения мягкой пшеницы Triticum aestivum (озимые сорта Московская 39 и Мироновская 808, яровой сорт Омская 36), двудольные растения Glycine soja (пять дикорастущих популяций из Приморского края) и G. max (сорнополевая форма сои, Китай), а также представители заводских и аборигенных пород крупного рогатого скота Bos taurus - черно-пестрого голштинизированного, айширского, якутского и красного эстонского (97 гол.). В качестве праймера в IRAP-PCR был выбран терминальный участок SIRE-1 (GCA-GTT-ATG-CAA-GTG-GGA-TCA-GCA). Полученные данные свидетельствуют о том, что полилокусное генотипирование по IRAP-PCR маркерам с использованием фрагмента ретротранспозона LTR SIRE-1 в качестве праймера позволяет надежно дифференцировать не только представителей однодольных и двудольных растений, но и их сорта. В исследованиях представителей разных пород крупного рогатого скота по IRAP-PCR маркерам получен спектр фрагментов ДНК (от 13 до 16 в зависимости от породы) в диапазоне длин 330-1470 п.н. Наибольший полиморфизм фрагментов ДНК наблюдался в средней части спектра (760-980 п.н.) у айширских и черно-пестрых коров. На дендрограмме одна из групп черно-пестрых коров и айширский скот оказались ближе друг к другу, тогда как другая вместе с красным эстонским и якутским скотом обособились в отдельный подкластер. Выявленные различия по гетерогенности в двух исследованных группах черно-пестрого скота из разных хозяйств, возможно, связаны с особенностями проводимой там селекционной работы либо с иными факторами искусственного и/или естественного отбора. Обсуждается возможность применения LTR-ретротранспозонов в качестве молекулярно-генетических маркеров для полилокусного генотипирования растений и животных.
Инвертированные терминальные сайты ltr-ретротранспозонов, полиморфное информационное содержание, доля полиморфных локусов
Короткий адрес: https://sciup.org/142133540
IDR: 142133540 | УДК: 636.082.12+631.52]:577.2:575.174.015.3
The fragments of homologies of endogenous retroviruses in the genomes of plants and animals
Using sequences of mobile genetic elements in order to mark polymorphic sites in their insertion might be the most effective approach to identify specific features of gene pools of different groups of organisms and control their dynamics, which is especially important in dealing with agricultural species. Sireviruses ( Pseudoviridae ) one of the oldest LTR-retrotransposons are widespread throughout the plant kingdom. SIRE-1 is one of the largest and most detailed studied retrotransposon. Its analysis showed that insertions of the retroelement (particularly in the genome of maize) have occurred recently. We studied the polymorphism of DNA fragments flanked by LTR-retrotransposon SIRE-1 (IRAP-PCR markers) in the genomes of different taxonomic groups. The objects of our study were Triticum aestivum (Moscovskaya 39, Mironovskaya 808, Omskaya 36 varieties), Glycine soja (five wild populations of Primorskii region of Russia) and G. max (China), as well as representatives of the factory and indigenous breeds of cattle - Black-and-White cattle improved by Holstein cattle, Ayrshire, Yakut and Red Estonian cattle (97 animals in total). A terminal site of SIRE-1 was chosen as primer IRAP-PCR (GCA-GTT-ATG-CAA-GTG-GGA-TCA-GCA). The data indicate that the multiloci genotyping by IRAP-PCR using retrotransposon LTR SIRE-1 as a marker reliably differentiates not only representatives of the monocotyledonous and dicotyledonous plants, but also their varieties. Spectrum of DNA fragments (13 to 16 depending on the breed) obtained in studies different breeds of cattle using IRAP-PCR markers was in the length range of 330-1470 bp. The highest polymorphism of DNA fragments was observed in the middle part of the spectrum (760-980 bp) in Ayrshire and Black-and-White cows. According to the obtained dendrogram one of the groups of Black-and-White cows and Ayrshire cattle were closer to each other, while the other with Red Estonian and Yakut cattle became isolated in a separate subcluster. Identified differences in heterogeneity in the two studied groups of Black-and-White from different farms possibly could be explained by the peculiarities of breeding work carried out or with other factors of artificial and/or natural selection. The possibility of using LTR-retrotransposons as molecular genetic markers for polyloci genotyping plants and animals is discussed.
Текст научной статьи Фрагменты гомологии эндогенных ретровирусов в геномах растений и животных
В последние годы разработано множество методов маркирования полиморфизма различных геномных участков (RFLP — restriction fragment length polymorphism, SNP — single nucleotide polymorphism, AFLP — amplified fragment length polymorphism, SSR-PCR — simple sequence repeats PCR и др.), успешно применяющихся для генотипирования как сортов культурных растений (1-4), так и пород сельскохозяйственных видов животных (5-7). Среди них особое место занимают мобильные элементы, первое и, вероятно, самое главное отличие которых от других структурно-функциональных элементов генома — это способность к перемещению. Высокая скорость транспозиций позволяет предполагать их существенную роль в генерации генетической изменчивости (8).
У растений LTR (long terminal repeat) ретротранспозоны занимают значительную часть генома: у Arabidopsis thaliana — чуть более 7 % (9), у риса — 50 %, у пшеницы — 90 % (10), у кукурузы — 75 % (11).
Sireviruses — одни из древнейших LTR-ретротранспозонов, получивших широкое распространение внутри царства растений. Это единственный представитель семейства Pseudoviridae, члены которого предположительно могут содержать ген, близкий к кодирующему оболочечные бел- ки вирусных частиц (env-подобный ген). К наиболее крупным по размеру (около 11 тыс. п.н.) и подробно изученным элементам Sireviruses относится SIRE-1. Анализ последовательности SIRE-1 показывает, что инсерции указанного ретроэлемента (в частности, в геном кукурузы) произошли недавно (12).
Использование последовательностей мобильных генетических элементов в целях маркирования полиморфизма участков их инсерций может оказаться наиболее эффективным подходом для выявления специфических особенностей генофондов для различных групп организмов, контроля их динамики, что особенно важно в работе с сельскохозяйственными видами.
В этой связи целью работы было изучение возможности использовать терминальные участки эндогенных ретровирусов для выявления сортоспецифичных характеристик у культурных растений, а также генетических особенностей у разных пород крупного рогатого скота.
Методика . Исследования выполняли на однодольных растениях Triticum aestivum и двудольных растениях Glycine soja и G. max . Пшеница была представлена двумя озимыми сортами (Московская 39 — мягкая озимая, Мироновская 808 — мягкая озимая, выведена из яровой) и одним яровым (Омская 36 — мягкая яровая), соя — пятью популяциями вида дикорастущая уссурийская ( G . soja , Приморский край) и сорнополевой формой сои ( G . max , Китай). В анализ также включили представителей заводских и аборигенных пород крупного рогатого скота (всего 97 гол.) — две группы черно-пестрых голштинизированных коров (I группа — с животноводческой фермы «Степаньково», Московская обл., II — из вивария Российского государственного аграрного университета), коровы айширской породы (МКЗ № 1, Московская обл.), якутский (Республика Саха) и красный эстонский скот (СПК «Заря», Псковская обл.).
Для полилокусного генотипирования использовали IRAP-PCR маркеры (IRAP-PCR — inter-retrotransposon amplified polymorphism PCR) (13). Геномную ДНК выделяли с использованием коммерческого набора «ДНК-экстран 1» («Синтол», Россия). ПЦР проводили на амплификаторе «Терцик» (Россия) с применением смеси ПЦР-РВ («Синтол», Россия). Условия и стадии ПЦР: первоначальная денатурация — 94 °С, 2 мин; денатурация — 94 °С, 30 с; отжиг — 55 °С, 30 с; элонгация — 72 °С, 2 мин; заключительная элонгация — 72 °С, 10 мин; 35 циклов. Праймерами служили терминальные участки LTR-ретротранспозона SIRE-1 (GCA-GTT-ATG-CAA-GTG-GGA-TCA-GCA). Результаты амплификации оценивали с помощью электрофореза в агарозном геле (1,5 %) с применением в качестве маркера молекулярных масс ДНК GeneRulerTM 100 bp DNA Ladder Plus («MBI Fermentas», США) для оценки длины продуктов. Визуализацию продуктов ПЦР-амплификации после окрашивания бромистым этидием проводили под УФ-светом. Для того чтобы избежать грубых ошибок, анализировали область спектра с размером фрагментов ДНК менее 1500 п.н.
Математическую обработку данных осуществляли с использованием компьютерной программы TFPGA. Расчет индекса PIC (polymorphic information content) выполняли по формуле для диаллельных локусов: PIC = 2f (1 - f), где f — частота одного из двух аллелей (14).
Поиск гомологичных последовательностей осуществляли в GenBank с использованием баз данных секвенированных участков, применяя для этих целей набор алгоритмов BLASTn.
Результаты . В связи с широкой представленностью LTR-ретротранспозонов (в том числе SIRE-1) в геномах растений для полилокусного генотипирования в качестве праймера в IRAP-PCR нами была выбрана терминальная область этого мобильного элемента.
В результате IRAP-PCR с праймером к терминальному участку ретроэлемента LTR SIRE-1 мы получили отчетливо воспроизводимые спектры фрагментов ДНК как у сои, так и у пшеницы, причем такие фрагменты находились в одном размерном диапазоне: суммарно до 22 локусов длиной 350-1240 п.н., 26 локусов — 220-1450 п.н. (рис. 1, 2).
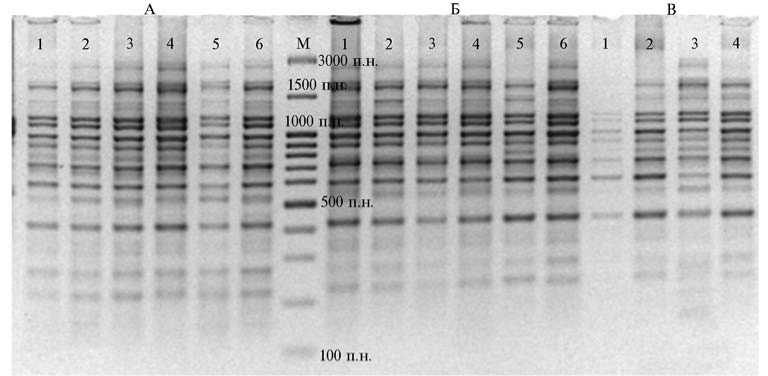
Рис. 1. Электрофоретические спектры, полученные в IRAP-PCR с ДНК пшеницы Triticum aestivum озимых (А — Московская 39, Б — Мироновская 808) и ярового (Омская 36) сортов при использовании в качестве праймера терминального участка ретротранспозона LTR SIRE-1: М — маркер молекулярных масс GeneRuler TM 100 bp DNA Ladder Plus («MBI Fermentas», США).
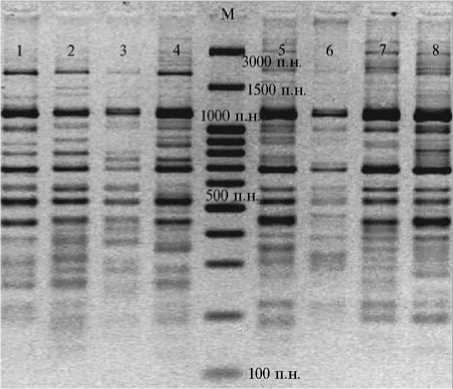
Рис. 2. Электрофоретические спектры, полученные в IRAP-PCR с ДНК дикорастущей сои Glycine soja разных групп при использовании в качестве праймера терминального участка ретротранспозона LTR SIRE-1: 1-4 —
I группа, 5-8 — II группа; М — маркер молекулярных масс GeneRuler TM 100 bp DNA Ladder Plus («MBI Fermentas», США).
гментов составили соответственно 0,449 и
Анализируемые группы Glycine обладали высоким полиморфизмом (PICср. = 0,414, P = 91 %) по сравнению с T. aestivum (PICср. = 0,120, P = 65 %; PICср. — усредненное значение индекса PIC по всему спектру ампликонов). С использованием баз данных GenBank был выполнен поиск участков гомологии к фрагменту LTR SIRE-1, послужившему праймером, в се-квенированных последовательностях геномов T. aestivum и G. max и выявлены соответственно 122 и 102 таких участка (табл. 1). У Glycine наиболее полиморфными оказались фрагменты длиной от 350 до 490 п.н. и от 1010 до 1240 п.н. Значения PICср. для этих фра-0,364. У T. aestivum, наобо рот, высокий полиморфизм наблюдался в зоне средних длин (от 520 до
720 п.н. и от 760 до 990 п.н., соответственно PICcp . — 0,196 и 0,155). Все тяжелые фрагменты были мономорфными и встречались у 100 % исследованных образцов пшеницы (табл. 2).
1. Число участков, гомологичных терминальным сайтам ретротранспозона LTR SIRE-1, в геномах сои Glycine max и пшеницы Triticum aestivum и их некоторые генетические характеристики по результатам IRAP-PCR
|
Показатель | |
G . max |
| T. aestivum |
|
Длина генома, млрд п.н.1 |
1,10 |
1,78 |
|
Число: |
||
|
найденных гомологичных участков1 |
122 |
102 |
|
фрагментов ДНК в спектре |
15-22 |
26 |
|
PIC усредненный по праймеру |
0,414 |
0,120 |
|
P, % |
91 |
65 |
Примечание. 1 — по данным ; IRAP-PCR — inter-retrotransposon amplified polymorphism PCR, PIC — индекс полиморфного информационного содержания локуса, P — доля полиморфных локусов.
2. Частота ампликонов (ЧА) в спектрах, полученных в IRAP-PCR с праймером, гомологичным терминальному участку ретротранспозона LTR SIRE-1, и значения индекса PICл0K у Cа у сортов пшеницы Triticum aestivum
|
Длина фрагмента, п.н. |
Московская 39 |
Мироновская 808 |
Омская 36 |
|||
|
ЧА |
1 PIC „окуcа |
ЧА |
1 PIC „окvcа |
ЧА |
1 PIC „окvcа |
|
|
1450 |
0 |
0 |
0,2 |
0,159 |
1 |
0 |
|
1420 |
1 |
0 |
1 |
0 |
1 |
0 |
|
1370 |
1 |
0 |
1 |
0 |
0,4 |
0,349 |
|
1340 |
0,8 |
0,483 |
0 |
0 |
0,4 |
0,349 |
|
1210 |
1 |
0 |
1 |
0 |
1 |
0 |
|
1130 |
1 |
0 |
1 |
0 |
1 |
0 |
|
990 |
1 |
0 |
1 |
0 |
1 |
0 |
|
900 |
1 |
0 |
1 |
0 |
1 |
0 |
|
810 |
1 |
0 |
0,3 |
0,300 |
1 |
0 |
|
790 |
1 |
0 |
0 |
0 |
0 |
0 |
|
760 |
0 |
0 |
0,8 |
0,483 |
0 |
0 |
|
720 |
1 |
0 |
1 |
0 |
1 |
0 |
|
680 |
1 |
0 |
1 |
0 |
1 |
0 |
|
630 |
0,500 |
0,414 |
0,800 |
0,483 |
1 |
0 |
|
590 |
1 |
0 |
1 |
0 |
1 |
0 |
|
570 |
0 |
0 |
0,3 |
0,3 |
0 |
0 |
|
550 |
0 |
0 |
1 |
0 |
1 |
0 |
|
540 |
1 |
0 |
0 |
0 |
0,2 |
0,189 |
|
520 |
0,300 |
0,300 |
0 |
0 |
0 |
0 |
|
460 |
1 |
0 |
0,700 |
0,488 |
0 |
0 |
|
420 |
1 |
0 |
1 |
0 |
1 |
0 |
|
360 |
1 |
0 |
1 |
0 |
1 |
0 |
|
320 |
1 |
0 |
1 |
0 |
1 |
0 |
|
270 |
1 |
0 |
1 |
0 |
1 |
0 |
|
250 |
0,500 |
0,414 |
0,300 |
0,300 |
0,200 |
0,189 |
|
220 |
1 |
0 |
1 |
0 |
1 |
0 |
|
р, % |
15 |
27 |
15 |
|||
|
Примечание. |
IRAP-PCR |
— inter-retrotransposon amplified polymorphism PCR, |
PIC — индекс по- |
|||
|
лиморфного информационного содержания локуса, P — доля полиморфных локусов. |
||||||
У Glycine был обнаружен только один мономорфный локус длиной 700 п.н., в среднем же доля полиморфных локусов по спектру составила 93 %, PICcp . = 0,414. Это свидетельствует об относительно высоком генетическом разнообразии исследованных групп как внутри рода Glycine , так и внутри одного вида G. soja. При этом у G. max отсутствовал локус длиной 680 п.н., тогда как тот же локус встречался у представителей G. soja.
В результате IRAP-PCR с использованием фрагмента ретротранспозона LTR SIRE-1 в качестве праймера были получены уникальные для каждого из сортов пшеницы спектры фрагментов ДНК. Так, у сорта пшеницы Московская 39 фрагмент ДНК длиной 790 п.н. присутствовал у всех исследованных образцов этого сорта, тогда как у сортов Мироновской 808 и Омской 36 фрагментов такой длины не обнаружили. И наоборот, локус, соответствующий фрагменту размером 550 п.н., не встречался только у сорта Московская 39, у остальных сортов по этому локусу PIC = 1.
Показано, что инбридинг в популяциях растений и дрозофилы, например, приводит к физиологическим и фенотипическим изменениям, которые, как предполагается, обусловлены эпигенетическими эффектами, связанными с попарным взаимодействием хромосом (15). Подобные эпигенетические изменения могут приводить к активации мобильных генетических элементов и, как следствие, к усилению процессов, вызывающих кариотипические модификации, к повышению генетического разнообразия и к эпигенетическим перестройкам, что служит важным источником фенотипической изменчивости (15).
Предложенной гипотезой может частично объясняться высокий полиморфизм внутри локуса у сортов T . aestivum с учетом того, что пшеница — самоопылитель. Известно также, что на долю растений с перекрестным опылением у этого самоопылителя приходится до 3 %, а иногда до 10 % популяции. Кроме того, выявлены формы с повышенной частотой переопыления (от 2,6 до 5 % в зависимости от сорта), и к ним, в частности, относятся сорта озимой мягкой пшеницы мироновской селекции (16). Следовательно, высокая внутрисортовая гетерогенность (высокая доля полиморфных локусов и увеличение индекса PIC) (см. табл. 2) у сорта Мироновская 808 по сравнению с двумя другими сортами может быть обусловлена этим фактом.
В связи с неодинаковой распространенностью у разных сортов наибольший интерес в случае пшеницы представляют локусы, соответствующие фрагментам 790 и 550 п.н., сои — локусы 790, 680 и 550 п.н.
На основании значений генетических дистанций (DN), рассчитанных по методу M. Nej (1972) исходя из частоты ампликонов разной длины в полученных спектрах фрагментов ДНК, фланкированных инвертированными участками фрагмента ретротранспозона LTR SIRE-1, нами была построена дендрограмма (рис. 3). На ней выделяются два отдельных крупных кластера однодольных и двудольных растений. В кластере однодольных в одну группу обособляются сорта пшеницы Мироновская 808 и Омская 36
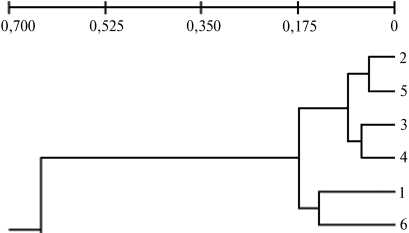
Рис. 3. Дендрограмма филогенетического родства между изученными группами растений, построенная на основании частоты ампликонов в IRAP-PCR с праймером, гомологичным терминальному участку ретротранспозона LTR SIRE-1: 1 — Glycine max (Китай); 2-6 — G . soja (Приморский край); 7-9 — Triticum aestivum , сорта соответственно Московская 39, Мироновская 808 и Омская 36.
I (см. рис. 3). Подобная диффе-•---9 ренциация сортов отражает как
7 их фенотипические особенности, так и происхождение. На пример, для сортов Мироновская 808 и Омская 36 общая фенотипическая характеристика — белый безостый неопушенный колос (17), тогда как Московская 39 отличается от них белым остистым неопушенным колосом . При этом озимый сорт Мироновская 808 был выведен из яровой мягкой пшеницы методом группового и массового отбора морфологически однородных растений (18).
Полученные данные свидетельствуют о том, что полилокусное генотипирование по IRAP-PCR маркерам с использованием фрагмента ретротранспозона LTR SIRE-1 в качестве праймера позволяет надежно дифференцировать представителей не только однодольных и двудольных растений, но и разных принадлежащих к ним сортов.
Результаты выполненного нами поиска в GenBank участков гомологии к фрагменту LTR ретротранспозона SIRE-1 с использованием алгоритмов BLASTn свидетельствовали о присутствии таких последовательностей в геномах не только растений, но и млекопитающих. Например, в се-квенированных последовательностях генома крупного рогатого скота выявляется 165 участков с такой гомологией.
Для оценки возможности применения фрагментов ретротранспозонов растений с целью полилокусного генотипирования и контроля генофондов пород сельскохозяйственных животных мы выполнили анализ групп некоторых пород крупного рогатого скота (айширский, красный эстонский, якутский скот, две группы черно-пестрых коров из разных хозяйств) по IRAP-PCR маркерам с использованием терминального сайта ретротранспозона LTR SIRE-1 в качестве праймера. При этом были получены спектры фрагментов ДНК (от 13 до 16 в зависимости от породы) в диапазоне длин 330-1470 п.н. (рис. 4).
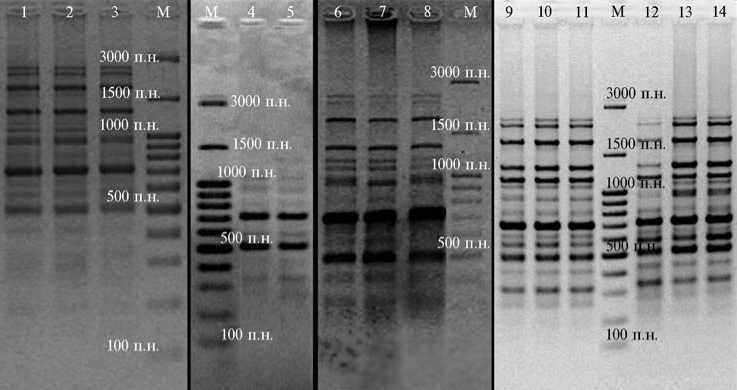
Рис. 4. Электрофоретические спектры, полученные в IRAP-PCR с ДНК крупного рогатого скота Bos taurus разных пород при использовании в качестве праймера терминального участка ретротранспозона LTR SIRE-1 : 1-3 — айширский скот, 4, 5 — красный эстонский скот, 6-8 — черно-пестрый скот (I группа), 9-11 — черно-пестрый скот (II группа), 12-14 — якутский скот; М — маркер молекулярных масс GeneRulerTM 100 bp DNA Ladder Plus («MBI Fermentas», США).
3. Значения индекса PICл0K у Cа в спектрах, полученных в IRAP-PCR с праймером, гомологичным терминальному участку ретротранспозона LTR SIRE-1, у крупного рогатого скота Bos taurus
|
Длина фрагмента, п.н. |
Айширский |
Черно-пестрый |
Якутский |
Красный эстонский |
|
|
II группа |
I группа |
||||
|
1470 |
0,325 |
0,486 |
0 |
0,488 |
0,452 |
|
1300 |
0,500 |
0 |
0,465 |
0 |
0 |
|
1200 |
0 |
0 |
0 |
0 |
0 |
|
1100 |
0 |
0 |
0 |
0 |
0 |
|
980 |
0,325 |
0,416 |
0,494 |
0 |
0,470 |
|
950 |
0,325 |
0 |
0 |
0 |
0 |
|
820 |
0,483 |
0 |
0,349 |
0 |
0 |
|
800 |
0,496 |
- |
0 |
0,380 |
0 |
Продолжение таблицы 3
|
760 |
0,121 |
- |
0,349 |
- |
0 |
|
720 |
0 |
0 |
0 |
0 |
0 |
|
640 |
0 |
0,330 |
0,432 |
0 |
0 |
|
590 |
0 |
0 |
0 |
0 |
0 |
|
510 |
0 |
0 |
0 |
0 |
0 |
|
440 |
0 |
0 |
0 |
0 |
0 |
|
380 |
0,361 |
- |
0,097 |
- |
0,137 |
|
330 |
0,457 |
0 |
0 |
0 |
0 |
Примечание. IRAP-PCR — inter-retrotransposon amplified polymorphism PCR, PIC — индекс полиморфного информационного содержания локуса. Прочерки означают, что локус отсутствует.
Наибольший полиморфизм фрагментов ДНК наблюдался в средней части спектра (760-980 п.н.) у черно-пестрых (I группа) и айширских коров. У остальных пород наиболее полиморфным оказался локус, соответствующий фрагменту размером 1470 п.н. В спектре ампликонов ДНК у черно-пестрых (II группа) и якутских коров отсутствовали локусы длиной 380 и 760 п.н., в то же время они встречались у других пород (табл. 3).
4. Характеристика полиморфизма локусов, выявленных в IRAP-PCR с праймером, гомологичным терминальному участку ретротранспозона LTR SIRE-
1, у крупного рогатого скота Bos taurus
|
Порода, группа скота |
1 PIC локусов |
1 P, % |
|
Айширский |
0,212 |
56 |
|
Черно-пестрый: |
||
|
I группа |
0,137 |
38 |
|
II группа |
0,094 |
23 |
|
Якутский |
0,062 |
14 |
|
Красный эстонский |
0,066 |
19 |
Спектры ампликонов, полученные в PCR с праймером LTR SIRE-1 суммарно по всем локусам, для представителей якут- ской, красной эстонской и чернопестрой (II группа) пород крупного рогатого скота существенно не различались по доле полиморфных локусов (P) и индексу PICср. (соответственно 14, 19 и 23 %; 0,062, 0,066 и 0,094). Наиболее гетерогенной при этом оказалась популяция айширского скота (PICср. = 0,212, P = 56 %) (табл. 4).
Примечание. IRAP-PCR — inter-retrotransposon amplified polymorphism PCR, PIC — индекс полиморфного информационного содержания локуса, P — доля полиморфных локусов. Описание групп см. в разделе «Методика».
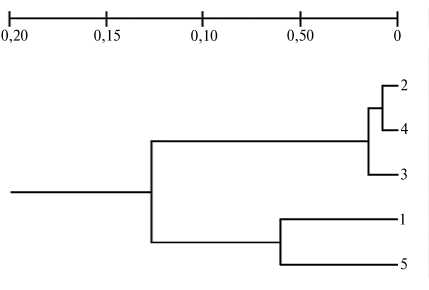
Рис. 5. Дендрограмма филогенетического родства между изученными породами крупного рогатого скота Bos taurus , построенная на основании частоты ампликонов в IRAP-PCR с праймером, гомологичным терминальному участку ретротранспозона LTR SIRE-1: 1 — айширский скот, 2 — черно-пестрый скот (I группа), 3 — якутский скот, 4 — красный эстонский скот, 5 — черно-пестрый скот (II группа).
На основании оценок полиморфизма ампликонов построили дендрограмму для исследуемых групп животных (рис. 5). Чернопестрый (I группа) и айширский скот оказались ближе друг к другу, тогда как черно-пестрый (II группа), красный эстонский и якутский обособились в отдельный подкластер. Необходимо отметить, что генетическая дистанция между черно-пестрыми и якутскими коровами была крайне мала (DN = 0,0076). Две группы черно-пестрых коров по генетическим характеристикам (см. табл. 4) имели значительные различия. Б о льшую гетерогенность отмечали у I группы (P = 38 %, PICср . = 0,137). Возможно, это связано с различиями в проводимой селекционной работе или иными факторами как искусственного, так и естественного отбора.
Таким образом, полученные данные свидетельствуют о том, что участки мобильных элементов, в частности LTR SIRE-1, могут применяться в исследованиях генетической структуры у групп и сортов как двудольных (Glycine), так и однодольных (Triticum) растений. Кроме того, такое полилокусное генотипирование фрагментов ДНК, полученных в результате IRAP-PCR с праймером, гомологичным терминальнаму сайту ретротранспозона LTR SIRE-1, позволяет выявлять генетические особенности пород крупного рогатого скота.
