Функциональная активность эндотелия роговицы свиньи

Автор: Батурина Г.С., Каткова Л.Е., Соленов Е.И., Пальчикова И.Г., Искаков И.А.
Журнал: Саратовский научно-медицинский журнал @ssmj
Рубрика: Глазные болезни
Статья в выпуске: 2 т.16, 2020 года.
Бесплатный доступ
Цель: исследовать возможность диагностики функциональной сохранности трансплантатов роговицы человека на основе оценки транспорта воды и ионов натрия клетками эндотелия роговицы свиньи. Материал и методы. Исследование проводилось на дисках роговицы свиньи. Оценка динамики объема клеток выполнялась методом, основанным на эффекте гашения флуоресценции красителя Calcein белками цитоплазмы. Содержание внутриклеточного натрия исследовалось методом флуоресцентной микроскопии с использованием специфического для натрия красителя Sodium Green. Для исследования динамики десвеллинга матрикса роговицы применяли микрофлуидную проточную камеру. Результаты. Повышение температуры с 20 до 37°С активирует Na/K-АТФазу, что запускает снижение объема клеток эндотелия роговицы. Экспоненциальная аппроксимация профиля флуоресценции дает траекторию, отражающую снижение клеточного объема с характерным временем 73,5±12,6 секунды (п=6). Экспоненциальная аппроксимация профиля флуоресценции красителя Sodium Green, отражающая изменение относительной концентрации внутриклеточного натрия, соответствовала снижению концентрации этого иона с характерным временем 130,0±26,3 секунды (п=5). Эксперименты с температурной активацией насосной активности клеток эндотелия позволили определить начальную скорость снижения объема матрикса: 0,36±0,006 um/s (п=5). Заключение. Оценка динамики изменения объема клеток эндотелия и содержания в них натрия, динамика десвеллинга после прекращения холодовой консервации
Внутриклеточный натрий, десвеллинг, объем клетки, холодовая консервация, эндотелий роговицы
Короткий адрес: https://sciup.org/149135442
IDR: 149135442 | УДК: 617.7.13
Corneal endothelium functional activity in pigs
The purpose of the study is to determine the potential for diagnostics of human corneal endothelial cells functional activity based on a study of the transport of water and sodium ions by the endothelial cells of a pig's cornea. Material and Methods. The study was conducted on the disks of pig's cornea. Evaluation of cell volume dynamics was studied with a method based on fluorescence quenching effect of Calcein dye by cytoplasmic proteins. The intracellular sodium concentration was investigated by fluorescence microscopy method using sodium-specific dye Sodium Green. Micro-fluidic flow chamber was used to study the dynamics of corneal matrix deswelling. Results. Increase of temperature from 20 to 37° С activates Na/K-ATPase and leads to a decrease in the volume of corneal endothelium cells (characteristic time 73.5±12.6 s) (n=6). The relative concentration of intracellular sodium decreased with the characteristic time 130.0±26.3 s (n=5). Studies with temperature activation of endothelial cell pumping activity allowed us to estimate the initial rate ofthe matrix volume reduction: 0.36±0.006 um/s (n=5). Conclusion. The study of the dynamics of endothelial cells' volume and intracellular content of sodium in it during cold preservation allows to improve the quality of assessment of viability of a preserved cornea.
Текст научной статьи Функциональная активность эндотелия роговицы свиньи
ведения и поступления воды и осмолитов в матрикс. Эндотелий представляет собой монослой клеток с плотными соединениями, разделяющий матрикс и жидкость передней камеры глаза. Интенсивность процессов водно-электролитного обмена в роговице в значительной мере определяется транспортными механизмами клеток эндотелия. Эти процессы часто описывают с помощью так называемого механизма “pump-leak” транспорта осмолитов и воды. Поступление воды в матрикс (leak) происходит пассивно согласно проницаемости эндотелия для воды и осмолитов и градиенту осмотического давления между матриксом и жидкостью в передней камере глаза. Удаление воды из матрикса (pump) обусловлено Na, K-ATФазой и системой, включающей транспорт HCO3", Cl", а также активность карбоангидраз клеток эндотелия. Оба процесса определяются функцией клеток эндотелия роговицы. Поступление воды в матрикс зависит от водной проницаемости монослоя клеток эндотелия, а формирование градиента осмотического давления определяется интенсивностью энергозависимого транспорта в этих клетках.
Цель: исследовать возможность диагностики функциональной сохранности трансплантатов роговицы человека на основе оценки транспорта воды и ионов натрия клетками эндотелия роговицы свиньи.
Материал и методы. Исследование проводилось на дисках роговицы свиньи диаметром 8,0 мм (трепанация через 6–8 часов после забоя животных), сохраняемых 2 суток при 4°С во влажной камере в консервационной среде Eusol-C (Alchimia, Италия).
Исследование содержания натрия и изменения относительного объема клетки. Исследование содержания внутриклеточного натрия. Изменения концентрации внутриклеточного натрия исследовали флуоресцентным методом с использованием красителя Sodium Green. Клетки эндотелия загружали красителем 5,0 μM Sodium Green AM (Molecular Probes, США) при 37°С 30 минут, затем помещали в проточную камеру микроскопа (рис. 1) в изотоническом растворе фосфатно-солевого буфера (PBS: 138 мM NaCl, 4,7 мМ Na2HPO4, 2,7 мМ KCl, 1,5 мМ KH2PO4, 0,5 мМ MgCl2, 5,5 мM глюкоза, 0,1 мМ CaCl2) с нормальной концентрацией натрия и уравновешивали в течение минимум 60 секунд. Флуоресценцию клеток эпителия регистрировали с помощью описанной установки.
Динамику изменения внутриклеточной концентрации натрия в клетках эндотелия записывали через 6-8 часов после забоя животных и после гипотермической консервации (4°C) препаратов роговицы глаза длительностью 2 суток.
Определение изменения относительного объема клетки. Для изучения изменений объема клетки применяли метод, основанный на эффекте гашения флуоресцентного красителя Calcein белками цитоплазмы [4]. Загрузка клеток красителем (Calcein AM, Sigma, Германия) производилась в среде L-15 Leibovitz с конечной концентрацией Calcein AM 10 мкМ, DMSO 1%. Время инкубации 15 минут при 37°С. Установка собрана на микроскопе Axiovert 40 (Zeiss, Германия). Для регистрации флуоресценции использовали комплект фильтров Zeiss #09, объектив х40, 0,5 N.A. Проточную камеру микроскопа и растворы термостатировали при 20°С или 37°С.
Эмитируемый свет регистрировали с помощью фотоприемника на основе ФЭУ-71, оборудованного полевой диафрагмой для измерения интенсивности флуоресценции в интересующей группе клеток. Измерения производили с помощью цифрового осциллографа АСК-3102 (Актаком ® , Россия) с записью на компьютер.
Ранее нами было показано, что изменения относительной флуоресценции (F/F0) линейно пропорциональны изменениям относительного объема клеток (V/V0) [3]. Изменения объема клетки выражали в относительных величинах флуоресценции.
Исследование влияния интегральной функции эндотелия на объем матрикса роговицы. Для исследования влияния интегральной “pump-leak” функции эндотелия фрагмент препарата роговицы помещали в проточную камеру, в которой поверхность препарата формирует микроканальную систему. В данной системе контакт препарата роговицы с протекающей средой осуществляется таким образом,
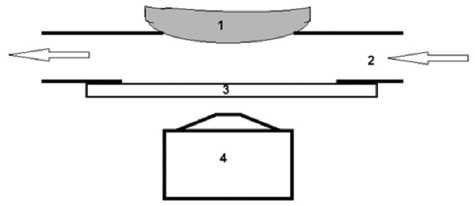
Рис. 1. Схема микроканальной проточной камеры:
1 — препарат роговицы; 2 — поток среды с флуоресцентным красителем; 3 — покровное стекло; 4 — микроскоп что водно-электролитный обмен между средой и матриксом происходит через эндотелий (см. рис. 1). Среда представляет собой PBS, содержащий 10–6 М Calcein в ионизированной форме, препятствующей его проникновению в клетки. Микроканальная система монтируется на стол инвертированного флуоресцентного микроскопа (Axiovert 40, Zeiss, объектив x10, 0.25 N. A.) и представляет собой канал с глубиной 500 и шириной 2000 микрометров. Среда протекает со скоростью 0,1 мл/мин. Дно канала выполнено из покровного стекла, противоположную сторону формирует поверхность образца роговицы. В этой связи глубина канала не постоянна и зависит от степени набухания препарата. В наших экспериментах она изменялась в интервале 200–400 микрометров. Амплитуда флуоресцентного сигнала в такой системе зависит от глубины протекающей среды, содержащей флуоресцентный краситель.
Для оценки вариабельности измеряемых величин рассчитывали стандартную ошибку среднего на основе несмещенной оценки дисперсии. Все величины выражены как М±т, n — количество препаратов роговицы. Процедура исследования соответствовала стандартам этического комитета, изложенным в Хельсинкской декларации 1975 г. и ее пересмотре 1983 г.
Результаты. Повышение температуры с 20 до 37°С активирует Na/K-АТФазу, что запускает снижение объема клеток, определяемого как изменение относительной флуоресценции красителя Calcein (F/F0), в клетках эндотелия роговицы с характерным временем снижения клеточного объема 73,5±12,6 секунды (n=6) и начальной скоростью снижения 0,0017±0,0004 секунды-1 (n=6) (рис. 2а).
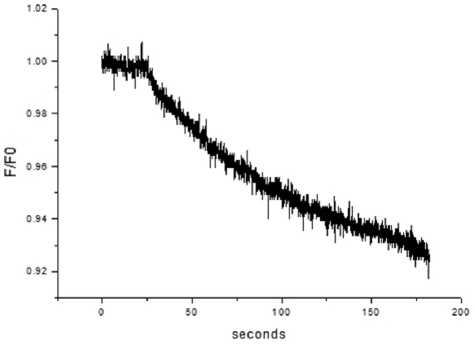
Рис. 2а. Типичный график флуоресценции красителя Calcein при повышении температуры препаратов в эндотелиальных клетках роговицы свиньи с 20 до 37°С.
Оси: абсцисс — секунды, ординат — относительная флуоресценция
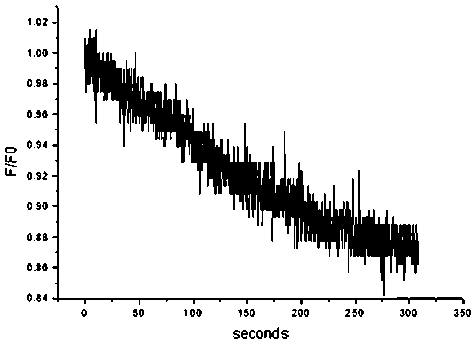
Рис. 2б. Типичный график флуоресценции красителя Sodium Green при повышении температуры препаратов в эндотелиальных клетках роговицы свиньи с 20 до 37°С.
Оси: абсцисс — секунды, ординат — внутриклеточная концентрация натрия, усл. ед.
Относительная концентрация внутриклеточного натрия, определяемая как величина относительной флуоресценции красителя Sodium Green (F/F0), при экспоненциальной аппроксимации профиля флуоресценции (рис. 2б) снижалась с характерным временем экспоненты 130±26,3 секунды (n=5).
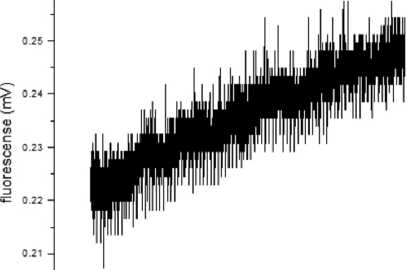
Эксперименты с температурной активацией насосной активности клеток эндотелия позволили сделать оценку начальной скорости снижения объема матрикса (десвеллинга) (рис. 3). Определение скорости снижения объема матрикса (0,36 ± 0,006 pm/s, n=5) позволяет сделать заключение, что кинетика этого процесса отражает эффективность интегральной насосной функции эндотелия.
Обсуждение. Основным источником энергии для векторного транспорта ионов через плазматическую мембрану является Na/К-АТФаза, создающая градиент электрохимического потенциала натрия, что является вторичным источником энергии для транспортных процессов в клетке и выполнения клетками насосной функции (pump) [5, 6]. В состоянии холодовой консервации Na/К-АТФаза не активна и клетки находятся в деполяризованном состоянии, следствием чего является набухание клетки в результате входа ионов натрия, поскольку в охлажденной, деполяризованной клетке устанавливается равновесная со средой концентрация ионов. В настоящей работе мы исследовали кинетику снижения относительного объема эндотелиальных клеток в результате выведения ионов натрия из клетки при температурной активации Na/K-АТфазы. Снижение клеточного объема в процессе выведения ионов натрия из клетки определяли по снижению флуоресцентного сигнала красителя Calcein. В препарате роговицы свиньи (2 суток после забоя животных), помещенном в изотонический раствор PBS, происходило снижение клеточного объема при повышении температуры. Эти результаты указывают на интенсивные процессы транспорта ионов натрия, приводящие к снижению клеточного объема (см. рис. 2а).
Поскольку снижение объема клетки при температуре 37°С в основном связано с активацией Na, K-АТФазы и выведением из клеток эндотелия ионов натрия, мы провели исследование динамики выхода натрия из клетки с помощью специфического для натрия флуоресцентного красителя Sodium Green. Согласно полученным результатам, харак-
0.26 1
0 100 200 300 «00
seconds
Рис. 3. Динамика флуоресцентного сигнала в микроканаль-ной камере в процессе снижения объема матрикса роговицы при повышении температуры с 20 до 37°С
терное время снижения относительного клеточного объема при экспоненциальной аппроксимации профиля флуоресценции начального этапа позволяет дать оценку потенциальной эффективности эндотелия трансплантата в процессе послеоперационного восстановления функции (см. рис. 2б). Скорость снижения относительной концентрации внутриклеточного натрия меньше, но близка по порядку величины к тем значениям, которые были получены в экспериментах по снижению объема, что согласуется с предположением об определяющем влиянии натрия на изменение клеточного объема. Таким образом, динамика внутриклеточной концентрации натрия может являться параметром, отражающим жизнеспособность и функциональную активность клеток эндотелия трансплантата роговицы.
Наши ранние исследования процесса выведения ионов натрия из клеток эндотелия показывают, что холодовая консервация в зависимости от длительности снижает интенсивность выведения натрия, а в сочетании с повышением проницаемости плазматической мембраны для этого иона ведет к установлению более высокого стационарного уровня внутриклеточного натрия в клетках эндотелия [7]. Результаты исследования интегральной “pump-leak” функции эндотелия фрагмента препарата роговицы, проведенные в экспериментальной микроканальной системе, также позволяют говорить о перспективности такого подхода. Эксперименты с температурной активацией насосной активности клеток эндотелия позволили сделать оценку скорости снижения объема матрикса (десвеллинг) (см. рис. 3). Кинетика снижения объема матрикса, представленная на рис. 3, характерна, по-видимому, для образцов с не поврежденным клеточным монослоем и отражает максимальную эффективность эндотелия. Такое заключение можно сделать на основании наших предыдущих исследований насосной функции клеток, из которых следует, что применяемый метод консервации приводит к постепенному снижению интенсивности, но тем не менее позволяет сохранять функцию клеточного транспорта более 10 дней [8]. В этой связи можно предположить, что в образцах с нулевой или низкой кинетикой десвеллинга доминирующим фактором является «утечка» (leak), т. е. поток воды через разрывы в эндотелии шунтирует и делает неэффективной работу клеточных насосов. Важным свойством такого подхода является его требователь- ность к целостности клеточного монослоя и плотных соединений между клетками, поскольку проницаемость эндотелия для успешности такого теста (leak) должна быть достаточно низкой. Резюмируя, можно отметить, что высокая кинетика десвеллинга в таком тесте с высокой вероятностью свидетельствует о высоком качестве образца как материала для трансплантации.
Заключение. Исследование динамики снижения клеточного объема эндотелия роговицы, содержания внутриклеточного натрия в клетках эндотелия, дес-веллинга матрикса роговицы при восстановлении функции после холодовой консервации, дает объективное представление о транспортной компетентности этих клеток и пригодности консервированной роговицы для трансплантации.
Список литературы Функциональная активность эндотелия роговицы свиньи
- Maurice DM. The location of the fluid pump in the cornea. J Physiol 1972; 221 (1): 43-54.
- Bonanno JA. Molecular Mechanisms Underlying the Corneal Endothelial Pump. Exp Eye Res 2012; 95 (1): 2-7.
- Zarogiannis SG, et al. Regulatory volume decrease of rat kidney principal cells after successive hypo-osmotic shocks. Math Biosci 2013; 244 (2): 176-87.
- Solenov E, et al. Sevenfold-reduced osmotic water permeability in primary astrocyte cultures from AQP-4-deficient mice, measured by a fluorescence quenching method. Am J Physiol Cell Physiol 2004; 286 (2): 426-32.
- Hoffmann EK, Lambert IH, Pedersen SF. Physiology of cell volume regulation in vertebrates. Physiol Rev 2009; 89 (1): 193-277.
- Wehner F, et al. Hypertonic activation of a non-selective cation conductance in HeLa cells and its contribution to cell volume regulation. FEBS Lett 2003; 551 (11): 20-4.
- Батурина Г.С., Пальчикова И.Г., Конев А.А. и др. Исследование влияния гипотермической консервации на уровень натрия в клетках эндотелия трансплантата роговицы. Вавиловский журнал генетики и селекции 2018; 22 (4): 433-7.
- Батурина Г.С., Каткова Л.Е., Пальчикова И.Г., Соленов Е.И., Искаков И.А. Новые подходы к исследованию функциональной активности клеток эндотелия препаратов роговицы глаза. Современные технологии в офтальмологии 2019; 5 (30): 262-5.
