Функциональная структура и устойчивость растительных сообществ нагорно-байрачных ландшафтных комплексов

Автор: Невский С.А., Давиденко О.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 1 т.9, 2007 года.
Бесплатный доступ
Статья посвящена оценке функционального состояния нагорно-байрачных растительных сообществ юга Саратовского Правобережья. Применен методический подход, предусматривающий построение стандартных распределений видов (геометрических рядов и Мак-Артура) и их сравнение с помощью нормированного индекса Шеннона. Выявлены сообщества разной степени устойчивости и дана экологическая интерпретация наблюдаемых закономерностей.
Короткий адрес: https://sciup.org/148197904
IDR: 148197904 | УДК: 581,524
Functional structure and stability of vegetation communities in upland-ravine habitat types
The article is devoted to the estimation of functional structure and stability vegetation communities in upland- ravine habitat in the south part of Volga right bank in Saratov region. A standard distributions of plants species (geometric rows and MacArthur), corresponding to the experimental one, and their comparison with the help of normalized Shannon index were applied. Different groups of stability were established and its ecological interpretation was performed.
Текст научной статьи Функциональная структура и устойчивость растительных сообществ нагорно-байрачных ландшафтных комплексов
УÄÊ 581.524
ФУНКЦИОНАЛЬНАЯ СТРУКТУРА И УСТОЙЧИВОСТЬ РАСТИТЕЛЬНЫХ СООБЩЕСТВ НАГОРНО-БАЙРАЧНЫХ
ЛАНДШАФТНЫХ КОМПЛЕКСОВ
Саратовский государственный университет им. Н.Г. ×ернышевского, г. Саратов
Статья посвящена оценке функционального состояния нагорно-байрачных растительных сообществ юга Саратовского Правобережья. Применен методический подход, предусматривающий построение стандартных распределений видов (геометрических рядов и Мак-Артура) и их сравнение с помощью нормированного индекса Шеннона. Выявлены сообщества разной степени устойчивости и дана экологическая интерпретация наблюдаемых закономерностей.
Bʙeäение
Оценка состояния лесных и лесостепных экосистем и прогноз их грядущих изменений необходимы для разработки экологических основ сохранения и воспроизведения лесных ресурсов, а также формирования сети охраняемых и рекреационных территорий [4]. При этом подходы к определению функционального состояния лесных растительных сообществ и установлению времени стабилизации лесных фитоценозов после нарушений на основании индексов энтропии остаются недостаточно проработанными. Вместе с тем энтропийно-информационный анализ в экологии зарекомендовал себя в качестве надежного метода оценки состояния экосистем [5, 8]. Целью работы была оценка степени функциональной структурированности наиболее распространенных в южной части Приволжской возвышенности растительных сообществ, сформированных в нагорно-байрачных комплексах типов местообитаний, нa основе определения их энтропии.
Материал и метоäы
Исследования проводились в 2003-2006 гг. на территории южной части Приволжской возвышенности, в Красноармейском районе Саратовской области. Район исследования относится к степной зоне (подзона типчаково-ковыльных степей) [11], лесные сообщества приурочены главным образом к нагорно-байрачным ландшафтным комплексам.
В основу работы положены геоботанические описания 17 наиболее распространенных в районе исследования растительных сообществ, в том числе 12 лесных ассоциаций, имеющих естественное, двух - искусственное происхождение и
3 степных фитоценозов. Bсе естественные сообщества являются условно ненарушенными, т.е. не испытывают заметного антропогенного воздействия. Геоботанические описания выполнены по стандартной методике [14] на площадках 400 м2 в лесных и 100 м2 - в степных сообществах. Экологическую структуру сообществ оценивали при помощи индексов увлажнения, трофности и освещенности, рассчитанных для каждого фитоценоза через проективное покрытие входящих в состав сообщества экологических групп видов [6, 13].
В качестве меры энтропии сообщества использовали индекс Шеннона, рассчитанный через распределение относительных проективных покрытий видов [7, 15, 16]. Был применен методический подход, предусматривающий построение стандартных распределений геометрических рядов и Мак-Артура, соответствующих экспериментальному, и их последующее сравнение с помощью нормированного индекса Шеннона. Этот подход был широко апробирован на примере различных лесных ассоциаций Средней Сибири [2, 3, 9, 10]. Соответствие распределения экспериментальных данных геометрическому ряду предполагает сформированность сообщества в результате конкурентной борьбы между видами, ведущей к максимальной упаковке экологических ниш. Альтернативой такому распределению выступает распределение Мак-Артура, при котором захват экологических ниш видами признается случайным [7]. Таким образом, по близости экспериментального индекса Шеннона конкретного растительного сообщества к той или иной теоретической модели распределения можно судить о степени сформированности и функциональной устойчивости сообщества.
Результаты и обсуæäение
На характер пространственного распределения растительности в районе исследования значительное влияние оказывает рельеф, перераспределяющий важнейшие экологические факторы и трансформирующий тепловой и гидрологический режимы [1]. На изученной территории наблюдается общая закономерность приуроченности естественных лесов к днищам балок и их теневым склонам. Световые склоны большей частью остепнены. Плакорные участки частично распаханы, частично используются под выпас и сенокошение, небольшие территории заняты смешанными искусственными посадками древесных пород.
В пределах естественных лесных сообществ наблюдаются свои закономерности распределения по элементам рельефа, проявляющиеся в основном в смене доминантов и содоминантов напочвенного покрова, а при значительном изменении условий - и в смене лесообразующих дре- весных пород.
К днищам суходольных балок приурочен липняк снытевый, теневые склоны заняты преимущественно дубравами ландышево-дубравномятликовой и ландышевой, липняком дубравномятликовым. В верхних частях теневых склонов распространены более ксерофильные ассоциации -липняк мезофитно-разнотравный и дубрава разнотравно-злаковая. На световых склонах встречается дубрава остепненная, но чаще эти местообитания остепнены.
В днищах балок с проточным увлажнением формируются вязовники, липо-осинники и ольшаники, в травяном покрове которых абсолютно доминируют сныть обыкновенная и крапива. На теневых склонах распространены липняки и дубо-липняки с мезофильным и гигромезофильным разнотравьем. В нижних частях световых склонов -дубравы ландышевая и разнотравно-злаковая [12].
На состав и структуру растительности влияют также профильные характеристики балок. Если русло их широкое и пологое, то получают
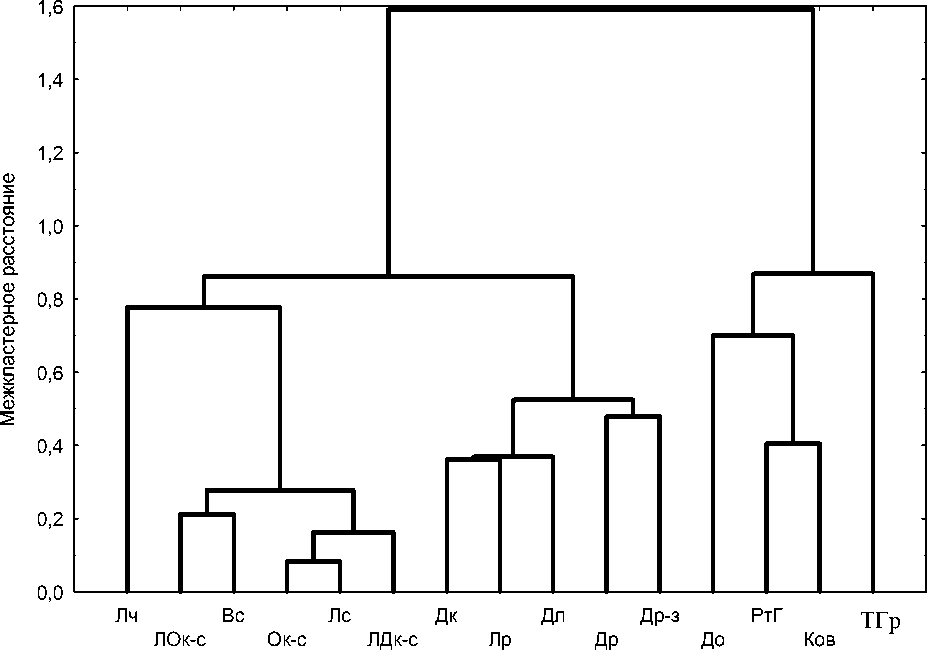
Рис. 1. Результаты оценки экологического сходства сообществ методом кластерного анализа.
Сообщества: Лч – липняк чистотеловый, ЛОк-с – липо-осинник крапивно-снытевый, Вс – вязовник снытевый, Ок-с – ольшаник крапивно-снытевый, Лс – липняк снытевый, ЛДк-с – липо-дубрава крапивно-снытевая, Дк – дубрава кирказоновая, Лр – липняк разнотравный, Дл – дубрава ландышевая, Др – дубрава разнотравная, Др-з – дубрава рaзнотравно-злаковая, До – дубрава остепненная, РтГ – разнотравно-грудницевый фитоценоз, Ков – ковыльный фитоценоз, ТГр – типчаково-грудницевый фитоценоз
распространение дубравы и липо-дубравы (сны-тевая, кирказоновая, чистотеловая и др.). П о мере углубления балки и увеличения крутизны ее скло-нов происходит повышение затененности и увлаж-нения местообитаний - формируются липовые сообщества с доминированием в напочвенном покрове сныти, чрапивы и других влаголюбивых видов. П о мере уменьшения глубины балки в верхней ее части происходит иссушение место -обитаний - снова встречаются дубравы и липо-дубравы от снытевой до разнотравно-злаковой.
На рис. 1 чредставлены результаты кластеризации естественных сообществ на основании сходства их экологических индексов влажности, трофности и освещенности, оычисленных для напочвенного покрова. П о обобщенной оценке структурной организации сообщества четко раз -деляются на три группы, в чоду которых идет снижение коэффициентов влажности и увеличе-ние освещенности. Инчекс трофности почв дает менее выраженный градиент в этом направле-нии и варьирует в пределах каждой из групп. Липняк чистотеловый по обобщенной оценке эко -логических индексов стоит несколько обособлен -но в группе фитоценозов наиболее увлажненных местообитаний. В ксерофитных сообществах подобное же положение занимает типчаково-грудницевый фитоценоз.
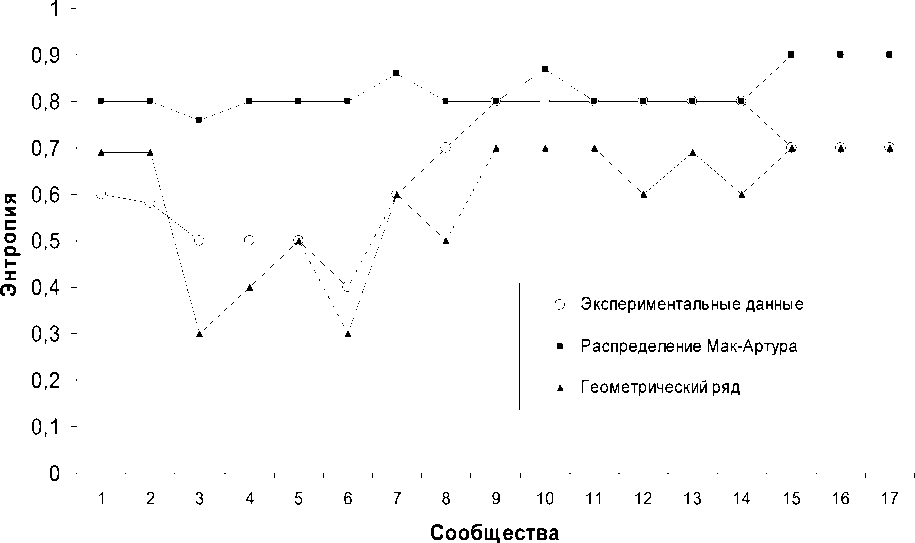
Оценка устойчивости изученных раститель-ных сообществ (рис. 2) показала, что в пределах каждой экологической группы фитоценозов пред -ставлены сообщества разной степени функцио-нальной сформированное™ и стабильности.
Энтропия всех степных фитоценозов полное -тью совпадает с таковой, рассчитанной для распределения геометрических рядов, что подтверждает наиболее полное соответствие данных со -обществ экологическим условиям местообита -ния и свидетельствует о структуре этих сооб-ществ, кчк основанной на дифференциации экологических ниш. Среди лесных ассоциаций наибольшей функциональной устойчивостью (распределение видов совпадает с геометрическим рядом) характеризуются дубрава ландышевая и липо - дубрава крапивно-снытевая. члизкими к данному состоянию являются липняк и вязовник снытевые.
При наличии части свободных ресурсов мес-тообитания, не распределенных между видами, значение энтропии стремится к нулю. Это характерно для наиболее влажных вариантов место -обитаний и сформированных здесь ассоциаций -
. 2.
-
1 - , 2 - - , 3,
-
4 , 5 - - . 6,
7 , 8 , 9,
10 , 11 - , 12,
13 , 14 , 15 -,
16 , 17- ольшаника крапивно-снытевого и липо-осинника крапивно-снытевого. Есть основание предположить, что подобные значения энтропии для данных сообществ связаны с тем, что в изученной ситуации они являются ненасыщенными в видовом отношении из-за отсутствия возможности заноса зачатков растений, приспособленных к условиям избыточного увлажнения. В таком случае проявляется абсолютное доминирование одного или двух видов, а доля участия остальных минимальна.
Примером сообществ, которые сформировались в результате случайного захвата экологических ниш видами и не пришли еще к устойчивому равновесию между компонентами, являются искусственные посадки, а также пограничные со степью участки леса - дубравы разнотравнозлаковая и остепненная (распределение видов этих фитоценозов полностью соответствует значениям распределения Мак-Артура). Дубравы кирказоновая и разнотравная занимают промежуточное положение между двумя теоретическими моделями распределения видов в сообществе, чуть больше тяготея к модели Мак-Артура. Липняк чистотеловый и вязовник снытевый также являются сообществами со средней степенью сформированности структуры, но со смещением в сторону геометрических рядов.
Список литературы Функциональная структура и устойчивость растительных сообществ нагорно-байрачных ландшафтных комплексов
- Болдырев В.А. Естественные леса Саратовского Правобережья. Эколого-ценотический очерк. Саратов, 2005.
- Зырянова О.А., Абаимов А.П., Бугаенко Т.Н. Оценка видового разнообразия коренных лиственничных ассоциаций криолитозоны и его послепожарной динамики на основе информационного индекса Шеннона//Сибир. экол. журн. 2004. № 5.
- Зырянова О.А., Бугаенко Т.Н., Бугаенко Н.Н. К вопросу изучения видового разнообразия коренных лесов криолитозоны Сибири//Исследовано в России (электронный журнал). 2002. №198; http://zhurnal.ape.relarn.ru/articles/2002/198.pdf.
- Коломыц Э.Г. Эмпирическая имитация регионального биоклиматического тренда экосистемами локального уровня//Изв. Самар. НЦ РАН. 2005. Т. 7, № 1.
- Крамаренко С.С. Метод использования энтропийно-информационного анализа для количественных признаков//Изв. Самар. НЦ РАН. 2005. Т. 7, № 1.
- Матвеев Н.М. Биоэкологический анализ флоры и растительности (на примере лесостепной и степной зоны): учебное пособие. Самара, 2006.
- Мэгарран Э. Экологическое разнообразие и его измерение. М., 1992.
- Савинов А.Б. Метод биоиндикации экосистем по соотношению адаптивных и инадаптивных потенциалов популяций и биоценозов (информационно-энтропийный аспект)//Методы популяционной биологии: Сб. материалов VII Всерос. популяционного семинара. Сыктывкар, 2004.
- Тараканова Ю.В. Использование нормированного индекса Шеннона для индикации коренных и условно-коренных сообществ//Тез. 7-й Пущинской шк.-конф. мол. ученых. Пущино, 2003.
- Тараканова Ю.В. Применение индекса Шеннона-Уивера для индикации структуры растительных сообществ//Тез. 6-й Пущинской шк-конф. мол. ученых. Пущино, 2002.
- Тарасов А.О. Основные географические закономерности растительного покрова Саратовской области. Саратов, 1977.
- Филиппова С.А., Невский С.А. Особенности структуры лесных сообществ в зависимости от положения в рельефе//Вопросы биологии, экологии, химии и методики обучения: Сб. науч. ст. Вып. 7. Саратов, 2004.
- Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М., 1983.
- Юннатов А.А. Типы и содержание геоботанических исследований. Выбор пробных площадей и заложение экологических профилей//Полевая геоботаника. М.; Л., 1964. Т. 3.
- Gatson K.J. Biodiversity-measurement. Progress in physical geography. 1994. 18 (4).
- Peet R.K. The measurement of species diversity//Ann. review of ecology and systematics. 1974. № 5.
