Функциональное состояние мышц тазовых конечностей у собак в условиях экспериментальной модели перелома седалищной кости

Автор: Сайфутдинов М.С., Антонов Н.И., Сизова Т.В.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 1, 2009 года.
Бесплатный доступ
ЭМГ-обследования 35 собак при моделировании переломов тела и ветви седалищной кости показали снижение активационной способности мышц тазовой конечности экспериментальных животных, которая восстанавливалась быстрее и полнее в условиях чрескостного остеосинтеза, чем в контроле при консервативном лечении. Механизмами изменения функционального состояния мышц при травме таза являются денервационно-реиннервационный процесс и защитный интерорецептивный рефлекс.
Электронейромиография, травма таза, внешний остеосинтез, денервационно-реиннервационный процесс
Короткий адрес: https://sciup.org/142121164
IDR: 142121164 | УДК: 619:616-073.97:616.718.2-001.5-089.84
The functional state of canine pelvic limb muscles in the experimental model of ischial bone fracture
EMG-studies of 35 dogs while modeling ischial bone body and ramus have revealed decrease of the activation ability of pelvic limb muscles of experimental animals, which was restored more rapidly and more completely under transosseous osteosynthesis, than in control under conservative treatment. Both the denervation-and-reinnervation process and the protective interoreceptive reflex are mechanisms of change in muscle functional state in case of pelvis injury.
Текст научной статьи Функциональное состояние мышц тазовых конечностей у собак в условиях экспериментальной модели перелома седалищной кости
Переломы костей таза часто (до 30 % случаев) сочетаются с поражением периферических нервов, что приводит к парезам и атрофиям мышц нижних конечностей [1]. При переломах седалищной кости сравнительно часто наблюдается повреждение седалищного нерва [2]. Трудность лечения больных с данной патологией отмечают многие исследователи, особенно при множественных и сочетанных повреждениях. В связи с вышесказанным была разработана экспериментальная модель перелома седалищной кости и проведены неврологические и электрофизиологические исследования при консервативном и оперативном методах лечения переломов тела и ветви седалищной кости у собак.
Целью настоящего исследования была оценка влияния моделирования комбинированной травмы бедра и таза на функциональное состояние мышц тазовых конечностей экспериментальных животных и степень восстановления их активационной способности в условиях чреско-стного остеосинтеза.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперименты проведены на 35 собаках 1 обоего пола в возрасте от года до пяти лет, весом 6-27 кг. В первой серии (18 животных -основная группа) осуществляли лечение травмы с помощью аппарата внешней фиксации [3]; во второй серии (17 животных – контроль) прово-
М.С. Сайфутдинов - ведущий научный сотрудник научного клинико-экспериментального отдела физиологии, к.б.н.;
Н.И. Антонов - научный сотрудник экспериментального отдела травматологии и ортопедии;
Т.В. Сизова - научный сотрудник научного клиникоэкспериментального отдела физиологии.
дили только консервативное лечение.
Стимуляционная электромиография (ЭМГ) по общепринятой методике [4, 5] в модификации, предложенной А.П. Шеиным, проводилась под внутривенным барбитуровым наркозом. ЭМГ покоя и вызванную биоэлектрическую активность (M-ответы) m. gastrocnemius, m. tibialis anterior и m. semitendinossus справа и слева регистрировали с помощью ЭМГ-системы «DISA-1500» (DAN-TEC, Дания). В первом случае отведение биопотенциалов было биполярным с использованием игольчатого электрода 13К13 (DANTEC, Дания), а во втором – монополярное, электродом с модифицированными отводящими поверхностями в виде игл. Активный полюс вводили в брюшко тестируемой мышцы, а индифферентный - под кожу в области сухожилия. М-ответы получали в результате раздражения седалищного нерва через игольчатый электрод прямоугольными импульсами длительностью 1 мс с заведомо супрамакси-мальной интенсивностью. Измеряли амплитуду М-ответа (от вершины максимального негативного до вершины максимального позитивного пика), его латентный период (время от артефакта раздражения до начала генерации суммарного потенциала действия мышцы) и длительность (отрезок времени от начала до окончания генерации суммарного потенциала действия мышцы). Оценка характера статистического распределения проводилась по нескольким критериям [6]: по степени соответствия вида гистограммы кривой нормального распределения, по степени совпадения значений среднего арифметического, моды и медианы анализируемой выборки данных, по величине асимметрии и эксцесса распределения, по величи- не соотношения среднего арифметического и стандартного отклонения. Поскольку в соответствии с используемыми критериями тип статистического распределения анализируемых данных соответствовал нормальному, рассчитывали среднее значение (М) и ошибку средней (m) изучаемого параметра и показатели его вариативности: стандартное отклонение (σ) и коэффициент вариации (KV) как отношение стандартного отклонения к среднему арифметическому [7, 8]. Степень различий средних значений по группам и в разные сроки восстановительного периода оценивалась с помощью t-критерия Стьюдента. Статистическую обработку данных проводили с помощью вычислительных средств Miсrosoft Excel-2000.
Содержание, кормление животных, операции и ЭМГ-обследования, а также выведение из опыта осуществляли в соответствии с Европейской конвенцией по защите позвоночных животных, используемых для экспериментальных и других научных целей [9], а также согласно приказу МЗ СССР № 775, 1977 года. 1
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Биоэлектрическая активность покоя тестированных мышц в большинстве наблюдений была в норме (табл. 1). В случае повышенного уровня фоновой ЭМГ, она была представлена комбинацией потенциалов фибрилляции и потенциалов фас-цикуляции. В ранние сроки эксперимента они сохраняют постоянную интенсивность, либо медленно и часто не до конца затухают. В контрольной группе частота встречаемости повышенного уровня ЭМГ покоя выше, чем в основной, она более интенсивна и разнообразно представлена в ранние сроки эксперимента. В поздние сроки наблюдения спонтанная ЭМГ встречается значительно реже, исключительно в контрольной группе. Она представлена в основном потенциалами фасцикуляции, которые возникают в ответ на введение электрода и быстро затухают. У животных основной группы в поздние сроки наблюдения спонтанная ЭМГ полностью нормализуется.
Результаты тестирования вызванной биоэлектрической активности мышц нижней конечности экспериментальных животных представлены в таблице 2 и на рисунке 1. В ранние сроки (до 1 месяца) после травмы средние значения амплитуды М-ответа мышц на стороне вмешательства снижены на 14,3-46,8 % статистически значимо (p<0,05) относительно соответствующих величин контралатеральной стороны. При этом данное снижение более выражено в контрольной группе (на 30,0-46,8 %), чем в основной (14,3-25,3 %), поскольку на стороне вмешательства средние значения амплитуды М-ответа всех тестированных мышц выше, чем в контроле, причём для m. gastrocnemius и m. semitendinossus данные раз- личия статистически значимы (p<0,05). Средние значения латентных периодов М-ответов тестированных мышц основной группы на стороне вмешательства и на контралатеральной конечности короче (p>0,05), чем в контроле, за исключением m. tibialis anterior. Данный ЭМГ-параметр в обеих группах несущественно больше (p<0,05) на стороне вмешательства, чем на контралатеральной. Средние значения длительности М-ответов мышц на стороне вмешательства в основной группе больше, чем в контрольной, но различия статистически значимы (p<0,05) только для m. semitendinossus. В контрольной группе средняя длительность М-ответов m. gastrocnemius и m. semitendinossus чуть ниже, а для m.tibialis anterior – чуть выше по сравнению с контралатеральной конечностью, в то время как в основной группе М-ответы мышц на стороне вмешательства имеют большие значения длительности, чем на контралатеральной.2
В сроки, превышающие три месяца после вмешательства, средняя амплитуда М-ответа мышц оперированной конечности остаётся сниженной относительно уровня контралатеральной, у животных основной группы на 9,6-32,9 % (для m. semitendinossus снижение статистически значимо, p<0,05), а у животных контрольной группы на 17,5-32,5 % (p<0,05). По сравнению с ранними сроками эксперимента данный ЭМГ-параметр на оперированной конечности у животных основной группы увеличился на 24,3 % (p<0,05), 20,1 (p>0,05) и 6,8 (p>0,05), соответственно для m. gastrocnemius, m. tibialis anterior и m. semitendinossus. В контрольной группе он увеличился на 11,3 % (p>0,05) для m. gastrocnemius и m. semitendinossus, а для m. tibialis anterior практически не изменился. На контралатеральной стороне в контрольной группе средняя амплитуда М- ответа снижалась для всех тестированных мышц на 6,7-12,2 % (p>0,05), в то время как в основной группе в поздние сроки обследования средняя амплитуда М-ответа увеличивалась на 17,8 % (p<0,05), 13,1 % (p>0,05) и 19,0 % (p>0,05) соответственно для m. gastrocnemius, m. tibialis anterior и m. semitendinossus.
Таблица 1
Количество наблюдений повышенного уровня биоэлектрической активности покоя мышц голени экспериментальных животных
|
Мышцы |
M. gastrocnemius |
M. tibialis anterior |
||||||||||
|
Группы |
контроль |
основная |
контроль |
основная |
||||||||
|
Срок эксперимента (дни) |
N |
N1 |
N2 |
N |
N1 |
N2 |
N |
N1 |
N2 |
N |
N1 |
N2 |
|
14 дней |
2 |
2 |
0 |
2 |
2 |
0 |
2 |
2 |
0 |
2 |
2 |
0 |
|
28 дней |
14 |
11 |
3 |
15 |
14 |
1 |
15 |
10 |
5 |
15 |
14 |
1 |
|
33 дня |
0 |
0 |
0 |
1 |
1 |
0 |
0 |
0 |
0 |
1 |
1 |
0 |
|
35 дней |
2 |
1 |
1 |
0 |
0 |
0 |
2 |
1 |
1 |
0 |
0 |
0 |
|
65 дней |
1 |
1 |
0 |
3 |
3 |
0 |
1 |
1 |
0 |
3 |
3 |
0 |
|
72 дня |
2 |
2 |
0 |
0 |
0 |
0 |
2 |
2 |
0 |
0 |
0 |
0 |
|
125 дней |
3 |
3 |
0 |
0 |
0 |
0 |
3 |
3 |
0 |
0 |
0 |
0 |
|
132 дня |
2 |
2 |
0 |
0 |
0 |
0 |
2 |
2 |
0 |
0 |
0 |
0 |
|
215 дней |
5 |
4 |
1 |
6 |
6 |
0 |
5 |
5 |
0 |
6 |
6 |
0 |
|
305 дней |
2 |
2 |
0 |
3 |
3 |
0 |
2 |
2 |
0 |
3 |
3 |
0 |
|
400 дней |
2 |
1 |
1 |
3 |
0 |
0 |
2 |
2 |
0 |
3 |
3 |
0 |
Примечание: N – общее количество наблюдений; N1 – количество наблюдений, где ЭМГ-покоя в норме; N2 – количество наблюде- ний с повышенным уровнем ЭМГ-покоя.
Таблица 2
Характеристики вызванной биоэлектрической активности мышц задних конечностей экспериментальных животных в разные сроки после травмы
|
Срок |
Мышца |
Амплитуда (мВ) |
LP (мс) |
T (мс) |
|||||||||||
|
n |
M±m |
Σ |
KV |
n |
M±m |
σ |
KV |
n |
M±m |
Σ |
KV |
||||
|
в о У и S § сЗ 02 О & S & с о |
M. gastrocnemius |
К |
15 |
18,1±2,2* |
8,4 |
46,6 |
14 |
3,5±0,2 |
0,7 |
20,4 |
14 |
26,8±1,9 |
7,0 |
26,2 |
|
|
О |
16 |
22,7±1,3*” |
5,3 |
23,6 |
15 |
3,0±0,1 |
0,4 |
14,1 |
14 |
28,3±1,2* |
4,5 |
15,8 |
|||
|
M. tibialis anterior |
К |
15 |
16,5±1,9* |
7,3 |
44,3 |
15 |
3,5±0,2 |
0,7 |
19,3 |
14 |
28,0±1,3 |
4,9 |
17,3 |
||
|
О |
16 |
19,2±2,0* |
7,8 |
40,9 |
15 |
3,5±0,1 |
0,5 |
14,8 |
15 |
30,4±1,4* |
5,4 |
17,7 |
|||
|
M. semitendinossus |
К |
13 |
5,5±0,7* |
2,6 |
46,9 |
11 |
2,4±0,2 |
0,7 |
29,4 |
8 |
18,4±2,7 |
7,7 |
41,9 |
||
|
О |
16 |
7,9±1,0*” |
4,1 |
52,4 |
13 |
2,1±0,3 |
0,9 |
44,7 |
9 |
30,5±4,4*” |
13,3 |
43,7 |
|||
|
M. gastrocnemius |
К |
15 |
20,2±1,8 |
7,1 |
35,0 |
15 |
3,3±0,2 |
0,6 |
18,4 |
15 |
30,7±1,2 |
4,6 |
15,0 |
||
|
О |
13 |
28,2±1,6 |
5,9 |
20,8 |
13 |
3,2±0,1 |
0,5 |
14,4 |
13 |
30,5±1,4 |
5,0 |
16,5 |
|||
|
M. tibialis anterior |
К |
15 |
16,2±1,9 |
7,5 |
46,2 |
14 |
3,6±0,2 |
0,6 |
15,9 |
14 |
30,9±1,7 |
6,2 |
20,1 |
||
|
О |
13 |
23,0±1,8 |
6,3 |
27,5 |
13 |
3,7±0,1 |
0,5 |
13,6 |
13 |
30,2±1,2 |
4,2 |
13,9 |
|||
|
M. semitendinossus |
К |
15 |
6,1±0,8 |
3,3 |
53,8 |
14 |
2,1±0,2 |
0,7 |
32,6 |
8 |
19,3±1,3 |
3,7 |
18,9 |
||
|
О |
13 |
8,4±1,3* |
4,6 |
54,9 |
10 |
1,9±0,1 |
0,4 |
21,8 |
9 |
24,2±1,1” |
3,4 |
14,2 |
|||
|
g § У S § 5 & 5 § Э |
M. gastrocnemius |
К |
16 |
26,2±1,7 |
6,9 |
26,5 |
16 |
3,3±0,2 |
0,8 |
23,2 |
16 |
27,3±1,0 |
3,9 |
14,3 |
|
|
О |
16 |
26,4±1,3 |
5,3 |
20,2 |
16 |
3,1±0,1 |
0,5 |
15,4 |
16 |
25,4±1,3 |
5,2 |
20,6 |
|||
|
M. tibialis anterior |
К |
16 |
23,6±1,7 |
6,7 |
28,5 |
16 |
3,3±0,2 |
0,7 |
22,3 |
16 |
27,6±1,1 |
4,4 |
16,0 |
||
|
О |
16 |
23,5±1,0 |
4,1 |
17,3 |
16 |
3,4±0,1 |
0,5 |
14,2 |
16 |
25,6±1,2 |
4,9 |
19,1 |
|||
|
M. semitendinossus |
К |
14 |
10,3±1,1 |
4,2 |
40,4 |
13 |
2,2±0,2 |
0,8 |
37,3 |
13 |
19,7±2,0 |
7,3 |
37,4 |
||
|
О |
16 |
10,5±0,8 |
3,2 |
30,6 |
16 |
1,8±0,1 |
0,4 |
22,4 |
13 |
22,2±1,4 |
4,9 |
22,3 |
|||
|
M. gastrocnemius |
К |
15 |
24,5±1,9 |
7,3 |
29,9 |
15 |
3,4±0,2 |
0,6 |
19,1 |
15 |
28,8±1,4 |
5,4 |
18,6 |
||
|
О |
13 |
31,2±1,8 |
6,4 |
20,5 |
13 |
3,2±0,1 |
0,5 |
14,5 |
13 |
27,9±1,3 |
4,6 |
16,5 |
|||
|
M. tibialis anterior |
К |
15 |
21,9±1,9 |
7,5 |
34,3 |
15 |
3,6±0,2 |
0,6 |
17,0 |
15 |
28,3±1,1 |
4,3 |
15,1 |
||
|
О |
13 |
26,5±1,7 |
6,1 |
22,9 |
13 |
3,5±0,1 |
0,5 |
14,8 |
13 |
28,8±1,3 |
4,8 |
16,7 |
|||
|
M. semitendinossus |
К |
14 |
9,1±1,0 |
3,7 |
41,1 |
14 |
1,9±0,2 |
0,8 |
40,9 |
11 |
19,7±3,0 |
10,1 |
51,1 |
||
|
О |
13 |
12,5±1,5 |
5,5 |
44,3 |
13 |
2,0±0,1 |
0,5 |
26,9 |
11 |
23,8±2,4 |
8,1 |
34,1 |
|||
Примечание: звёздочкой отмечено статистически значимое (p<0,05) отличие средних значений ЭМГ-параметра от уровня контралатеральной конечности; кавычками отмечено статистически значимое (p<0,05) отличие средних значений ЭМГ-параметра от уровня контрольной группы.
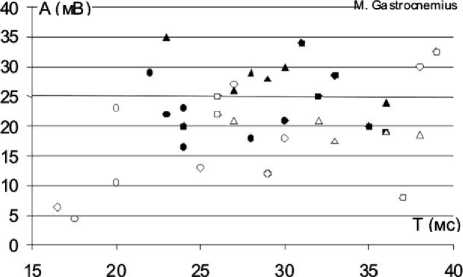
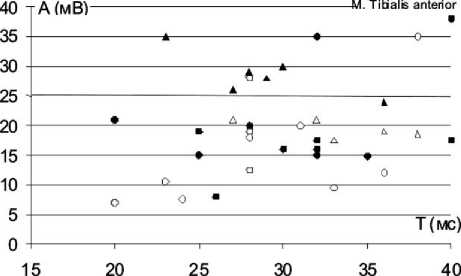
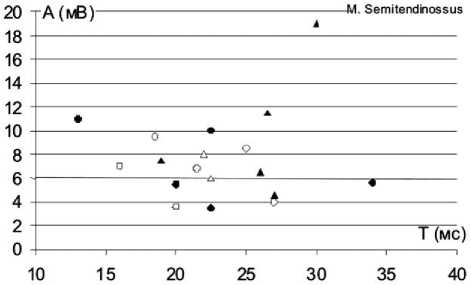
О - контрольная группа, 28 дней после вмешательства
Д - контрольная ipynna, более 3 месяцев после вмешательства ф- основная группа, 28 дней после вмешательства
А - основная группа, более 3 месяцев после вмешательства
Рис. 1. Индивидуальные значения характеристик вызванной биоэлектрической активности мышц нижних конечностей у экспериментальных животных в разные сроки восстановительного периода: ось абсцисс – значения длительности М-ответа (Т), выраженные в милисекундах (мс), ось ординат – амплитуда М-ответа (А), выраженная в миливольтах (мВ)
Средние значения латентных периодов М-ответов мышц оперированной и контралатеральной конечностей в основной и контрольной группах имеют близкие значения (p>0,05). Степень изменения этого ЭМГ-параметра относительно уровня, наблюдаемого в ранние сроки эксперимента, также несущественна (p>0,05). Средние значения длительности М-ответов мышц голени на стороне вмешательства в основной и контрольной группах очень близки, в то время как для m. semitendinossus в основной группе сохранилось преобладание более высокой, чем в контроле длительности (p<0,05). Средние значения длительности большинства М-ответов мышц контралатеральной конечности чуть меньше (p<0,05), чем на стороне вмешательства. По сравнению с ранними сроками эксперимента средняя длительность М-ответов тестированных мышц контрольной группы на оперированной и контралатеральной конечностях увеличивалась, соответственно на 5,114,7 % и 0,4-5,4 %. Для m. gastrocnemius на стороне вмешательства это увеличение статистически значимо (p<0,05). В основной группе незначительное увеличение длительности имеет место для m. gastrocnemius и m. semitendinossus контралатеральной конечности. Для m. tibialis anterior прирост длительности статистически значим. На стороне вмешательства длительность М-ответа m. gastrocnemius также незначительно увеличивается, в то время как для m. tibialis anterior и m. semitendinossus длительность сокращается, в последнем случае статистически значимо (p<0,05).
На рисунке 1 представлено распределение индивидуальных значений ЭМГ-характеристик тестированных мышц задних конечностей экспериментальных животных в разные сроки восстановительного периода. На рисунке видно, что множество значений анализируемых ЭМГ-параметров всех тестированных мышц вытянуто вдоль оси абсцисс. В ранние послеоперационные сроки множество значений ЭМГ-параметров m. gastrocnemius в основной группе охватывает более широкий диапазон длительностей М-ответа, а наблюдения, относящиеся к основной группе, расположены более компактно в центральной части графика. Для m. tibialis anterior области распределения ЭМГ-данных основной и контрольной групп практически совпадают, однако видно, что наблюдения, относящиеся к основной группе, в большей степени группируются в центре и в правом верхнем углу графика, а случаи, относящиеся к контрольной группе, находятся преимущественно в нижней части графика, т.е. у животных основной группы чаще встречаются М-ответы с большей, чем в контроле, амплитудой и длительностью, что соответствует соотношению средних значений данных ЭМГ-характеристик. Для m. semitendinossus наблюдается картина, обратная той, что отмечена для m. gastrocnemius.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Снижение амплитуды М-ответа мышц на стороне вмешательства при некотором увеличении их латентных периодов и разнонаправленном изменении длительности суммарного потенциала действия мышцы свидетельствует о наличии в двигательных единицах тестированных мышц реактивных изменений, вызванных травмой и снижающих общее функциональное состояние нейромоторно-го аппарата травмированной конечности. Данные реактивные изменения могут быть связаны с сосудистой реакцией на травму и с денервационно-реиннервационным процессом, на что указывает наличие потенциалов фибрилляции в ранние сроки наблюдения. Снижение амплитуды М-ответов на стороне вмешательства более выражено у контрольных животных, что свидетельствует о том, что реактивные изменения у них преобладают над основной группой. Наличие тенденции к увеличению латентных периодов и сокращению длительности также указывает на присутствие в этих изменениях денервационного компонента [10]. При моделировании травмы тазовых костей возникает обширная гематома, способная оказать на седалищный нерв механическое воздействие и влияние на биохимическом уровне. Кроме того, в ряде случаев дополнительное механическое действие могут оказать и костные отломки. Под влиянием механической альтерации и посттравматических изменений внутренней среды частично нарушается барьерная функция периневральной оболочки, что приводит к отклонению в составе интранев-ральной жидкости и появлению реактивных изменений в нервных волокнах [11]. Следует отметить, что у собак реактивные изменения в нервных волокнах, вызванные альтерирующим воздействием на нервный ствол, даже при высоком уровне его альтерации, достигали с током аксоплазмы соответствующей мышцы значительно быстрее, чем у человека. Так, при создании тотальных дефектов седалищного нерва денервационная активность высокой интенсивности наблюдалась уже на пятые сутки после операции [12, 13]. Поэтому на 1429-е сутки после моделирования травмы последствия альтерации седалищного нерва были в полной мере выражены не только в m. semitendinossus, но и в мышцах голени.
Поскольку альтерирующее воздействие на седалищный нерв при моделировании травмы и уровень гипокинезии в раннем послеоперационном периоде у животных контрольной и ос- новной групп были примерно одинаковы, то меньшее, чем в контроле, снижение средней амплитуды М-ответа и тенденция к увеличению его длительности у животных основной группы в срок 14-29 дней эксперимента можно объяснить более интенсивным протеканием у них восстановительных процессов и, в частности, реиннервации, на что указывает тенденция к возрастанию длительности М-ответов [10] и меньшая частота встречаемости спонтанной ЭМГ в ранние сроки, при её полном исчезновении в позднем периоде наблюдения у животных основной группы. Это согласуется с данными о высоком уровне регенераторной способности нервно-мышечного аппарата у собак, полученными в экспериментах по созданию значительных дефектов седалищного нерва [14], когда были выявлены электрофизиологические и морфологические признаки спонтанной реиннервации мышц голени у экспериментальных животных без замещения дефекта нерва.
Однако следует помнить, что наличие потенциалов фасцикуляции может быть не только признаком реиннервации, но и реакцией на наркоз [15]. Тем не менее разница в частоте встречаемости фасцикуляций в основной и контрольной группах свидетельствует о том, что влияние наркоза на генерацию потенциалов фасцикуляции в анализируемой нами выборке незначительно. Присутствие потенциалов фасцикуляции может быть связано также с развитием защитного тормозного рефлекса на травму, который проявляется в повышении спонтанной активности некоторой части тонических двигательных единиц [16], потенциалы действия которых регистрируются в виде фасцикуляций. Этим может объясняться сохранение фасцикуляций у контрольных животных в поздние сроки эксперимента из-за недостаточно стабильного по сравнению с основной группой, уровня фиксации костных отломков, что является источником дополнительного раздражения инте-рорецепторов, активирующих защитный рефлекторный механизм [17].
Таким образом, ЭМГ-тестирование мышц нижних конечностей экспериментальных животных в условиях комбинированной травмы бедра и таза выявляет снижение функционального состояния нервно-мышечного аппарата повреждённой конечности, менее выраженное у животных основной группы.
ВЫВОДЫ
-
1. Моделирование комбинированной травмы бедра и таза приводит к снижению функционального состояния нервно-мышечного аппарата повреждённой конечности.
-
2. Использование метода чрескостного ос-
- теосинтеза для лечения комбинированной травмы бедра и таза у экспериментальных животных способствует более быстрому, чем у контрольных животных восстановлению активационной способности мышц нижних конечностей.
