Функционирование малатдегидрогеназного компекса в мезофилле и обкладке кукурузы в условиях солевого стресса

Автор: Епринцев А.Т., Федорина О.с
Журнал: Журнал стресс-физиологии и биохимии @jspb
Рубрика: Original article
Статья в выпуске: 2 т.2, 2006 года.
Бесплатный доступ
Индуцированные солью изменения в работе ферментов малатдегидрогеназной системы вносят существенный вклад в формирование адаптации клетки к действию стрессора. Наибольший интерес в этом вопросе представляют ферментные системы С4-растений в связи с их лучшей приспособленностью к условиям окружающей среды. В данной работе исследована роль отдельных компонентов малатдегидрогеназного комплекса мезофилла и обкладки кукурузы в формировании адаптивной реакции в стрессовых условиях. На первом этапе воздействия соли в мезофилле кукурузы наблюдалась активация всех ферментов малатдегидрогеназной системы, с последующим снижением их активности. В обкладке данные показатели отличались от контрольных менее существенно. Было также обнаружено индуцированное солью быстрое накопление пирувата в клетках обкладки и малата в обеих исследуемых тканях. Дальнейшее засоление приводило к падению концентрации указанных интермедиатов. В обкладке содержание пирувата было сначала ниже контрольного уровня, а к концу экспозиции повышалось. Результаты исследований свидетельствуют об индукции цикла трикарбоновых кислот хлоридом натрия в клетках мезофилла и обкладки кукурузы и указывают на интенсификацию цикла Хэтча-Слейка в стрессовых условиях. Обнаруженные отличия в функционировании малатдегидрогеназной системы мезофилла и обкладки листьев кукурузы при засолении главным образом заключаются в том, что активность ферментов изучаемого комплекса в обкладке подвержена минимальным изменениям по сравнению с мезофиллом. На основании полученных данных обсуждается роль МДГ ферментной системы в механизмах адаптивной реакции различных тканей кукурузы к засолению
Засоление, малатдегидрогеназный комплекс, мезофилл/обкладка
Короткий адрес: https://sciup.org/14323455
IDR: 14323455
Function of malatdehydrogenase complex of maize mesophyll and bundle sheath cells under salt stress condition
Salt-induced changes in malatdehydrogenase system activity make the essential contribution to cell adaptation to stress condition. The enzyme systems of C4-plants are most interesting due to their ability for adaptation to environment conditions. The role of separate components of malatdehydrogenase complex of mesophyll and bundle sheath cells of com in formation of adaptive reaction in stressful conditions is investigated in presented work. The activation of all enzymes of malatdehydrogenase system and the subsequent decrease in their activity was observed in mesophyll durring the first stage of adaptation to salt influence. In bundle sheath cells such parameters are differed from control less essentially. Fast accumulation of piruvate in cells and malate in both investigated tissues was induced. The further salinity led to falling of concentration this intermediate. The concentration of piruvate was below control level, and it was raised by the end of an exposition. The results show that sodium chloride causes induction of Krebs-cycle in mesophyll and bundle sheath cells of com and intensification of Hatch-Slack cycle. The described differences in function malatdehydrogenase systems of mesophyll and bundle sheath cells of leaves of com under salinity mainly consist of the activity of enzymes of a studied complex in bundle sheath cells is subject to the minimal changes in comparison with mesophyll. Role of this enzyme system in mechanisms of adaptive reaction of various tissues of com to salt stress is discussed.
Текст обзорной статьи Функционирование малатдегидрогеназного компекса в мезофилле и обкладке кукурузы в условиях солевого стресса
Одним из наиболее распространенных стрессоров, оказывающих влияние на организм растения, является соль. Проблемы адаптации растений к солевому стрессу особенно актуальны в связи с ухудшением экологии почв. Засоление среды вызывает нарушение осмотического и ионного гомеостаза растительных клеток, а также накопление в них токсичных веществ (Балнокин, 2005). В метаболизме подверженных действию стрессора клеток происходят значительные изменения, направленные на нейтрализацию указанных негативных эффектов. Изменяется работа ионно-транспортных систем, происходит накопление осмолитов и осмопротекторв:
пролина, аланина, бетаина, манитолла и д.р (Балнокин, 2005).
Известно, что малат играет существенную роль в поддержании внутренних физиологических условий растительных клеток (Пинейру де Карвалью и др., 1991). Двойной путь утилизации малата с помощью малатдегирогеназ и малик-энзимов позволяет организму уменьшить зависимость от гликолиза при синтезе энергии и углеродных скелетов. Растительная МДГ (малатдегидрогеназная) система представляет собой динамическое равновесие белков, способное четко реагировать на физиологическое состояние и потребности организма, а также на изменения окружающей среды (Пинейру де Карвалью и др., 1991). Малатдегидрогеназный комплекс представлен четырьмя дегидрогеназами, две из которых обладают оксидоредуктазной активностью, а две другие – декарбоксилирующие МДГ (Иванищев, Курганов, 1992). Благодаря работе данной ферментной системы, осуществляется стыковка и сопряжение отдельных метаболических процессов в клетках.
Наиболее изученным ферментом МДГ комплекса является НАД+-зависимая оксидоредуктазная МДГ (К.Ф.1.1.1.37), катализирующая превращение малата в оксалоацетат в цикле Кребса. Кроме того, данная МДГ отвечает за синтез малата и оксалоацетата (Пинейру де Карвалью и др., 1991; Епринцев, Попов, 1999)
НАД+-зависимый малик-энзим (К.Ф.1.1.1.39) в присутствии НАД+ превращает малат в пируват. За счет работы этого фермента протекает обходной путь окисления малата в тах условиях, когда оксидоредуктазная дегидрогеназа (К.Ф.1.1.1.37) блокируется высоким уровнем ОАА, низким рН или недостаточной концентрацией ацетил-КоА (Пинейру де Карвалью и др., 1991; Иванищев, Курганов, 1992).
НАДФ+-зависимая декарбоксилирующая МДГ (малик-энзим) (К.Ф.1.1.1.40) кукурузы участвует в механизме фиксации и транспорта СО 2 и в регенерации НАДФН для биосинтетических процессов (Пинейру де Карвалью и др., 1991).
НАДФ+- зависимая оксидоредуктазная МДГ (К.Ф.1.1.1.82) играет роль при переносе в цитоплазму восстановительных эквивалентов, синтезированных в ходе фотофосфорилирования (Пинейру де Карвалью и др., 1991; Иванищев, Курганов, 1992).
Индуцированные солью изменения в работе ферментов малатдегидрогеназной системы вносят существенный вклад в формирование адаптации клетки к действию стрессора (Косулина и др., 1993).
Известно, что растения, относящиеся к С 4 -типу (кукуруза, сорго, амарант), имеют более сложную морфо-физиологическую структуру и лучше приспособлены к современным условиям окружающей среды, чем С 3 -растения, в том числе к засолению (Балнокин, 2005). В связи с этим, целью нашей работы было изучение роли отдельных компонентов малатдегидрогеназного комплекса мезофилла и обкладки кукурузы в формировании адаптивной реакции к условиям солевого стресса.
МАТЕРИАЛЫ И МЕТОДЫ
Объектом наших исследований служили 10-ти дневные проростки кукурузы сорта
Воронежская-76, выращенные гидропонным методом. Растения подвергали в течение 6 часов воздействию раствора хлорида натрия (концентрация 150мМ), ионы которого обладают низкой физиологической активностью в растительной клетке. Контролем служили образцы, экспонированные в воде. Первую пробу брали перед внесением соли, затем пробы снимали через 1, 3 и 6 часа экспозиции.
Проводили разделение мезофилла и обкладки по методу Клечковского (Епринцев и др., 2005) Чистоту разделенных тканей опредиляли по активности маркерных ферментов – ФЕП-карбоксилазы и НАДФзависимой-глицеральдегидфосфатдегидрогеназы, о которой судили по изменению оптической плотности, обусловленному утилизацией или накоплением НАДН. Активность малатдегидрогеназ также определяли спектрофотометрически при длине волны 340 нм (Землянухин, Землянухин, 1996). За единицу ферментативной активности принимали количество фермента, превращающего 1мкмоль субстрата за 1 минуту при стандартных условиях.
Для изучения содержания малата и пирувата, в полученные гомогенаты тканей добавляли 3% HClO4 в соотношении 1:5 и центрифугировали 5 минут при 7тыс.g. Супернатант нейтрализовали NaHCO3 и снова центрифугировали. Для определения содержания малата использовали гидразин-глициновый буфер, рН 9.5, 0.01М NAD+ и коммерческий препарат МДГ аналитической чистоты. Для определения пирувата брали 0.3М трис-HCl-буфер, рН 7.6, 0,005М HAДH и коммерческий препарат лактатдегидрогеназы аналитической чистоты. О содержании исследуемых интермедиатов в пробах судили по изменению оптической плотности при длине волны 340 нм, сопровождающей восстановление или окисление HAДH, которое фиксировали на СФ-46 (Детлаф Т.А., 1974).
В работе представлены средние данные наиболее типичных опытов. Было проведено два независимых эксперимента, каждая проба бралась в трёх биологических и двух химических повторностях. Статистическую обработку данных проводили по Лакин, 1990.
РЕЗУЛЬТАТЫ
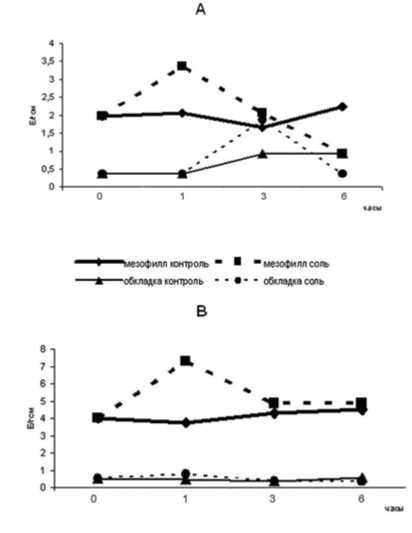
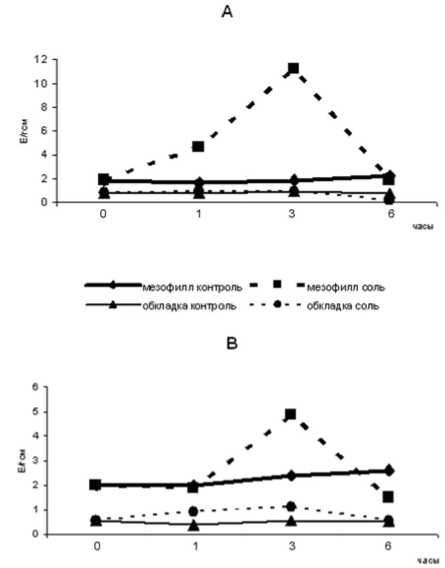
Данные о динамике активности ферментов МДГ- комплекса в клетках мезофилла и обкладки в условиях 150мМ засоления представлены на рисунках1 и 2.
Показано, что под действием хлорида натрия в мезофилле кукурузы происходит активация НАД+зависимой МДГ в первые часы действия стрессора (её актиность становится в 1,6 раз выше контрольной). Затем активность фермента в опытных образцах заметно падает и через 6
часов экспозиции становится в 2,4 раза ниже, чем в контроле. В обкладке соль также вызывает увеличение ферментативной активности НАД+зависимой МДГ, однако пик (в 2 раза больше контрольного значения) приходится на более позднее время (через три часа). Здесь, как и в мезофилле, дальнейшее засоление приводит к снижению активности в 2,5 раза (рис. 1).
Засоление приводило к активации НАД+-зависимого малик-энзима мезофилла на протяжении всего времени экспозиции. Наибольшая разница между опытными и контрольными значениями была зафиксирована через 3 часа засоления и составляла 95,2 %. В обкладке активность данного фермента в условиях опыта существенно не отличалась от контроля, хотя в первый час засоления была на 63% выше (рис. 1).
Активность НАДФ+-зависимого малик-энзима под действием хлорида натрия в мезофилле значительно увеличивается, достигая максимума ( в 12,9 раз выше, чем в контроле) к 3 часам экспозиции, однако, через 6 часов значение ферментативной активности в опытных образцах сравнивается с контрольной величиной. Различия контрольных и опытных показателей в обкладке для данного фермента не выходили за пределы погрешности (рис. 2).
Что касается функционирования НАДФ+-зависимой МДГ, то индуцированный солью всплеск активности мезофильной формы
^^^■мкс-^ипп юитроль ■ ■ ■ мезофилл соль
Рис.1. Влияние солевого стресса на НАД-зависимые ферменты МДГ-комплекса мезофилла и обкладки кукурузы: А- оксидоредуктазная МДГ (К.Ф.1.1.1.37); В- декарбоксилирующая МДГ (К.Ф.1.1.1.39).
фермента обнаруживается через три часа экспозиции. К 6 часам её величина падает и становится ниже контрольного уровня. В обкладке показана активация данной МДГ на протяжении шести часов засоления по сравнению с контролем (рис. 2).
Также были исследовано влияние 150мМ хлорида натрия на содержание малата и пирувата в мезофилле и обкладке кукурузы (рис3). Выявлено, что соль индуцирует быстрое накопление малата как в мезофилле (на протяжении трех часов действия стрессора), так и в обкладке кукурузы (в течение часа). Дальнейшее засоление приводило к падению концентрации малата ниже контрольного уровня, как в мезофилле, так и в обкладке кукурузы.
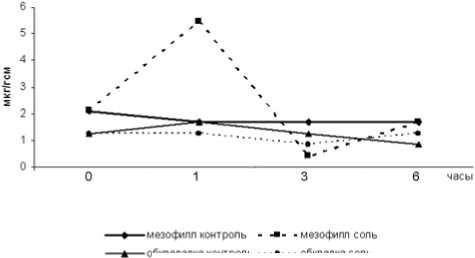
Солевой стресс в первый час экспозиции вызывал увеличение содержания пирувата – в 3,3 раза по сравнению с контролем - в мезофилле и снижение его концентрации в клетках обкладки в 1.3 раза (рис. 4). В мезофилле через три часа солевого воздействия значения концентрации пирувата были значительно ниже контрольных, а через 6 часов выходили на контрольный уровень. В клетках обкладки после 3 часов засоления содержание пирувата в опытных образцах было все ещё ниже, чем в контроле, а к концу экспозиции превышало контрольный уровень в 1,5 раза.
^■^^^^™ мезофи л л i он троль ■ ■ ■ мезофилл соль
---А ■ об мидиконтроль - - •- - обклмисоль
Рис.2. Влияние солевого стресса на НАДФ+-зависимые ферменты МДГ-комплекса мезофилла и обкладки кукурузы А-декарбоксилирующая МДГ (К.Ф.1.1.1.40); В- оксидоредуктазная МДГ (К.Ф.1.1.1.82).
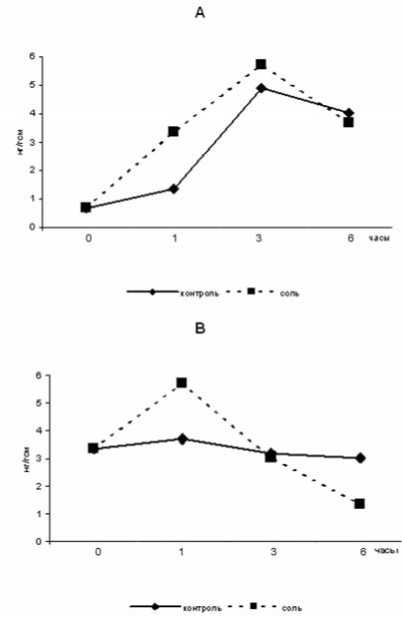
Рис.3. Изменение концентрации малата в клетках кукурузы под действием NaCl: А- мезофилл В- обкладка.
обклададка контроль обкладка соль
Рис.4. Изменение концентрации пирувата в клетках кукурузы под действием NaCl
ОБСУЖДЕНИЕ
Анализируя полученные данные мы пришли к заключению, что хлорид натрия вызывает интенсификацию основных метаболических процессов в клетках мезофилла и обкладки кукурузы в первые часы действия стрессора. Активация НАД-зависимой МДГ в обеих тканях свидетельствует об индукции ЦТК в стрессовых условиях, что соответствует известному эффекту «солевого дыхания» (Головко, 1999). Однако, специфическая особенность данной МДГ в клетках обкладки заключается в более позднем, по сравнению с мезофиллом, ответе на действие стрессора. Ранее подобная закономерность была обнаружена для маркерного фермента ЦТК фумаратгидратазы, а также для аконитатгидратазы из мезофилла и обкладки кукурузы (Артемьева, 2002).
Наблюдаемая в мезофилле значительная интенсификация хлоридом натрия НАДФ+-зависимого малик-энзима возможно связана с быстрым накоплением в стрессовых условиях пирувата (рис 4), выполняющего по крайней мере две функции: во-первых, обеспечивает клетку энергией; во-вторых, является предшественником аланина – аминокислоты, играющей существенную роль в осморегуляции клеток (Штеменко, Сорочан, 2001). В наших предыдущих исследованиях было показано, что содержание аланина в листьях кукурузы в стрессовых условиях гораздо выше, чем в контроле (Епринцев и др., 2002). Возможно, часть молекул пирувата, синтезированного в ответ на засоление в первый час экспозиции в дальнейшем идёт на синтез аланина, чем можно объяснить падение его концентрации, наблюдаемое через три часа действия соли (рис. 4).
Концентрационные колебания малата в мезофилле и обкладке кукурузы в условиях солевого стресса, главным образом, обусловлено функционированием цикла Хэтча-Слейка (Эдвадс, Уокер, 1986). Накопление малата в течение светового дня в мезофилле проходит гораздо интенсивнее при засолении в первые 3 часа действия стрессора за счет активации НАДФ+-зависимых МДГ и малик-энзима (пики их активности приходятся на это время).
Известно, что образовавшийся в мезофилле малат мигрирует в клетки обкладки, где подвергается окислительному декарбоксилированию с помощью НАДФ-зависимого малик-энзима. Образовавшийся СО 2 направляется на восстановительный пентозо-фосфатный путь, а пируват может принимать участие в процессах осморегуляции клеток (Эдвадс, Уокер, 1986). В значительной степени это подтверждается активацией НАДФ+-зависимой МДГ и высоким содержанием малата в клетках обкладки, подверженных действию NaCl в первый час экспозиции.
Таким образом, в ходе наших исследований были выявлены существенные отличия в функционировании МДГ-системы мезофилла и обкладки листьев кукурузы при засолении, которые заключаются в том, что активность ферментов изучаемого комплекса в обкладке подвержена минимальным изменениям по сравнению с мезофиллом. Дифференциация данных тканей на физиолого-биохимическом уровне дает им возможность по-разному реагировать на изменения окружающей среды, что в конечном итоге создает оптимальные условия для функционирования растительного организма в стрессовых условиях.
Список литературы Функционирование малатдегидрогеназного компекса в мезофилле и обкладке кукурузы в условиях солевого стресса
- Артемьева С.С., Солодилова О.С., Федорова Т.С.,Хомченкова Г.А., Епринцев А.Т. Действие солевого стресса на ферменты различных метаболических путей//Организация и регуляцияфизиолого-биохимических процессов. 2002. С.27 -34.
- Балнокин Ю.В. Растения в условиях стресса//Физиология растений под ред. Ермакова И.П./М: Издательский центр «Академия», 2005. -640 с.
- Головко Т.К. Дыхание растений. (физиологические аспекты). Спб.: Наука. -1999. -С.204.
- Епринцев А.Т., Ивентьев А.Н., Попов В.Н. Распределение и свойства изоформ гликолатоксидазы из клеток обкладки и мезофилла листьев амаранта//Физиология растений. -2005. -Т.52. -№.4. -С.622-627.
- Епринцев А.Т., Попов В.Н. Ферментативная регуляция метаболизма ди-и трикарбоновых кислот в растениях//Воронеж: Изд-во ВГУ. -1999. -С.134-135.
- Епринцев А.Т., Солодилова О.С., Хожаинова Г.Н. Роль свободных аминокислот в адаптивной реакции кукурузы в условиях солевого стресса//Вестник ВГУ, №2, серия химия, биология. 2002
- Землянухин А.А., Землянухин Л.А. Большой практикум по физиологии и биохимии растений. Воронеж: изд-во ВГУ 1996. С.121-128.
- Иванищев В.В., Курганов Б.И. Ферменты метаболизма малата: характеристика, регуляция активности и биологическая роль//Биохимия Т.57 вып.5, 1992, С.653-661.
- Косулина Л.Г. Физиология устойчивости растений к неблагоприятным факторам среды/Л.Г. Косулина, Э.К. Луценко, В.А. Аксенова. -Ростов -на -Дону, 1993, 126.
- Лакин Г.Ф. Биометрия//Москва. Высш. шк.-1990.-351 с.
- Пинейру де Карвалью М.А.А., Землянухин А.А., Епринцев А.Т. Малатдегидрогеназа высших растений//Воронеж: Изд-во ВГУ. -1991.-216с.
- Детлаф Т.А. Проблемы биологии развития. Методы биологии развития.//М: Наука. -1974. 280с.
- Штеменко Н.Т., Сорочан О.О. Вшьт амшокислоти на раншх фазах проростаиня зерна кукурудзи//Физиология и биохимия культурных растений. -2001. -Т.33. -№.5. -С.441-445.
- Эдвадс Дж., Уокер Д. Фотосинтез С3-и С4-растений: механизмы и регуляция. М.: Мир, 1986. 590с.
