Галектин-9/Tim-3-зависимый механизм регуляции функциональной активности натуральных киллеров при беременности

Автор: Орлова Е.Г., Логинова О.А., Горбунова О.Л., Ширшев С.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Иммунология
Статья в выпуске: 4, 2023 года.
Бесплатный доступ
Галектинами называют семейство β-галактозид-связывающих низкомолекулярных белков (лектинов), гомологичных по аминокислотной последовательности углевод-связывающего сайта. Галектины взаимодействуют с гликанами на поверхности клеток и участвуют в регуляции функциональной активности большинства клеток организма человека. Галектин-9 играет важнейшую роль в контроле функций клеток иммунной системы. Особое значение приобретают иммунорегуляторные эффекты галектина-9 при беременности, когда его продукция клетками трофобласта значительно увеличивается, что ассоциировано с формированием периферической иммунной толерантности. Основным лигандом для галектина-9 является молекула Tim-3 (Т-клеточный иммуноглобулиновый и муциновый домен, содержащий белок 3), высокая экспрессия которой выявляется на натуральных киллерах (NK-клетки). При беременности фенотип и функции NK-клеток существенно меняются, что необходимо для предотвращения цитотоксических реакций на клетки плода. В обзоре систематизированы данные о роли галектина-9 в Tim-3-опосредованной регуляции функций NK-клеток в аспекте беременности. Изучение регуляции функциональной активности NK-клеток необходимо для повышения эффективности NK-клеточной терапии.
Галектин-9, nk-клетки, tim-3, иммунная толерантность, беременность
Короткий адрес: https://sciup.org/147242774
IDR: 147242774 | УДК: 632.939:612.112.94:612.63 | DOI: 10.17072/1994-9952-2023-4-403-413
Galectin-9/Tim-3-dependent mechanism of regulation of functional activity of natural killers during pregnancy
Galectins are a family of β-galactoside-binding low molecular weight proteins (lectins) homologous in the amino acid sequence of the carbohydrate-binding site. Galectins interact with glycans on the cell surface and are involved in regulating the functional activity of most cells of the human body. Galectin-9 plays a crucial role in controlling the functions of cells of the immune system. Of particular importance are the immunoregulatory effects of galectin-9 during pregnancy, when its production by trophoblast cells increases significantly, which is associated with the formation of peripheral immune tolerance. The main ligand for galectin-9 is the Tim-3 (T-cell immunoglobulin and mucin domain 3) molecule, whose high expression is detected on natural killers (NK cells). During pregnancy, the phenotype and functions of NK cells change significantly, which is necessary to prevent cytotoxic reactions to fetal cells. The review systematizes data on the role of galectin-9 in Tim-3-mediated regulation of NK cell functions in the aspect of pregnancy. The study of the regulation of the functional activity of NK cells is necessary to increase the effectiveness of NK cell therapy.
Текст обзорной статьи Галектин-9/Tim-3-зависимый механизм регуляции функциональной активности натуральных киллеров при беременности
Галектинами называют семейство β-галактозид-связывающих низкомолекулярных белков (лектинов), гомологичных по аминокислотной последовательности углевод-связывающего сайта и распознающих остатки β-галактозы [Рапопорт, Курмышкина, Бовин, 2008; Рапопорт, Бовин, 2015]. В настоящее время у млекопитающих идентифицировано 16 представителей семейства галектинов, которые отличаются друг от друга строением и углеводной специфичностью [там же]. Галектины продуцируются разными типами клеток (эпителиальными, фибробластами, эндотелиальными, дендритными клетками, макрофагами, активированными Т- и В-лимфоцитами, костномозговыми стромальными клетками, натуральными киллерами (NK-клетки), регуляторными Т-лимфоцитами (Treg) и присутствуют во многих тканях и органах: плаценте, легких, печени, костном мозге, селезенке, сердце, лимфатических узлах, предстательной железе и др. [Рапопорт, Бовин, 2015; Iwasaki-Hozumi et al., 2021; Boron et al., 2022; Fuselier et al., 2023]. Галектины синтезируются и локализуются в цитоплазме клетки и в незначительных количествах присутствуют в ядре, а также при активации клетки секретируются в межклеточное пространство [Рапопорт, Курмышкина, Бовин, 2008; Рапопорт, Бовин, 2015]. Секреция галектинов осуществляется, главным образом, в ответ на провоспалительные стимулы при помощи микровезикул, которые образуются выпячиванием мембраны, минуя эндоплазматический ретикулум и аппарат Гольджи [там же]. Галектины оказывают как дистантные эффекты, так и способны связываться с компонентами гликокаликса клеток-продуцентов [там же]. Лигандами для галектинов являются углеводные цепи гликолипидов и гликопротеинов в составе различных клеток организма [там же]. Причем галектины могут вступать во взаимодействие как с гликанами разных клеток, обеспечивая межклеточные коммуникации, так и сшивать углеводные структуры на поверхности одной клетки [Рапопорт, Курмышкина, Бовин, 2008; Рапопорт, Бовин, 2015]. Внутри клетки галектины принимают участие в сигнальной трансдукции, взаимодействуя с другими белками цитоплазмы и ядра. Есть данные о том, что галектины участвуют в сплайсинге матричных РНК. Описаны специфические мембранные и внутриклеточные рецепторы для галектинов [там же].
Галектины играют важную роль в регуляции большинства клеточных процессов, таких как дифференцировка клеток, адгезия, межклеточные коммуникации, реализация эффекторных функций, индукция апоптоза, отторжение трансплантата [Iwasaki-Hozumi et al., 2021]. Концентрация галектинов повышается при воспалительных процессах [Iwasaki-Hozumi et al., 2021; Boron et al., 2022; Fuselier et al., 2023]. Избыточная экспрессия галектинов характерна для опухолевых клеток различного происхождения, что свидетельствует об их важной роли в прогрессии опухолевого роста в результате иммуносупрессии [Fuselier et al., 2023]. Некоторые галектины участвуют в адгезии вирусов к поверхности клеток-мишеней [Elahi et al., 2012].
Эффекты галектинов, главным образом, имеют противовоспалительный характер и направлены на ограничение избыточной активации клеток при воспалительных реакциях, защиту клеток от повреждений и индукции аутоиммунных процессов [Sanchez-Fueyo et al., 2003; Zhu et al., 2005]. Однако разные галектины в зависимости от типа клеток и используемых рецепторов оказывают разнонаправленные эффекты на их функциональную активность. Так, взаимодействие галектина-9 с дендритными клетками, макрофагами, Тreg стимулирует их активность, тогда как его связывание c рецепторами на T-лимфоцитах хелперах (Тh1) вызывает их апоптоз [Sanchez-Fueyo et al., 2003; Zhu et al., 2005; Nagahara et al., 2008; Ocana-Guzman, Torre-Bouscoulet, Sada-Ovalle, 2016]. Все вышесказанное свидетельствует о высокой актуальности изучения регуляторных эффектов разных типов галектинов.
Установлено, что галектин-9, действуя через Tim-3-рецепторы (Т-клеточный иммуноглобулиновый и муциновый домен, содержащий белок 3), препятствует отторжению аллогенного кожного лоскута и ускоряет его приживление [Wang et al., 2008]. Беременность также является феноменом полуаллогенной трансплантации, поскольку плод экспрессирует антигены отца. Ключевыми эффекторами формирования периферической иммунной толерантности при беременности являются натуральные киллеры (NK-клетки), которые в большом количестве экспрессируют Tim-3 [Sun et al., 2016; Li et al., 2017]. Логично полагать, что галектин-9-Tim-3-сигналинг при беременности играет значимую роль в модуляции функций NK-клеток и сохранении беременности. Поэтому целью данного обзора является систематизация знаний о роли галектина-9 в Tim-3-опосредованном контроле функций NK-клеток в аспекте беременности. Понимание механизмов регуляции функциональной активности NK-клеток при беременности необходимо для прогнозирования и предотвращения патологий беременности иммунного генеза (бесплодие, задержку внутриутробного роста, повторные выкидыши и др.), а также для повышения эффективности NK-клеточной терапии.
Галектин-9
Галектин-9 относят к тандемному типу галектинов, для которых характерно наличие двух углевод-связывающих доменов, распознающих разные углеводные лиганды [Рапопорт, Курмышкина, Бовин, 2008; Рапопорт, Бовин, 2015]. Галектин-9 впервые был идентифицирован как хемоаттрактант для эозинофилов [Asakura et al., 2002]. Он продуцируется разными типами клеток: эозинофилами, эндотелиальными клетками, CD4+ и CD8+Т-лимфоцитами, дендритными клетками, макрофагами, лимфоидными клетками, клетками Купфера, эпителиальными клетками кишечника, клетками трофобласта и плаценты [Enninga et al., 2018; Iwasaki-Hozumi et al., 2021]. Продукция галектина-9 усиливается при воспалении, онкологических и аутоиммунных заболеваниях [Chen et al., 2023]. Интерферон-гамма (INF-гамма) повышает экспрессию галектина-9 в эндотелиальных клетках и фибробластах, что объясняет механизм обратной связи в негативной регуляции Th1-воспалительного ответа [Imaizumi et al., 2002; Asakura et al., 2002].
Во время беременности клетки трофобласта активно продуцируют галектин-9, и его концентрация значительно нарастает в периферической крови уже с первых недель беременности и остается повышенной до родов [Meggyes et al. 2014; Enninga et al., 2018; Boron et al., 2022; Pełech et al., 2023]. Так в работе E.A.L. Enninga с соавторами показано, что уже на 8 неделе беременности уровень галектина-9 в периферической крови в три раза выше, чем у небеременных (8 неделя беременности – 1 976 пг/мл; у небеременных – 773 пг/мл), и такой уровень сохраняется до родов [Enninga et al., 2018]. При этом беременные женщины, вынашивающие мужской плод, имеют более высокие концентрации галектина-9 в периферической крови по сравнению с женщинами, которые вынашивают женский плод (2 263 пг/мл и 1 874 пг/мл, соответственно) [там же]. Данный факт дополняет наши представления о возможных причинах предрасположенности к развитию разных осложнений беременности у женщин, которые вынашивают женский и мужской плод. Другими исследователями было показано, что уровень галектина-9 нарастает в периферической крови пропорционально сроку беременности [Meggyes et al., 2014].
В исследовании D.G. Boron с соавторами также выявлялись более высокие значения концентрации галектина-9 в периферической крови (14 760 пг/мл) перед родами (30 неделя беременности) по сравнению с ранее описанными исследованиями [Boron et al., 2022]. В работе A. Pełech с соавторами показано, что концентрация галектина-9 в периферической крови (7 800 пг/мл) перед родами (39 неделя беременности) позитивно коррелирует с индексом массы тела и количеством жировой ткани, однако взаимосвязи с полом плода данные авторы не выявили [Pełech et al., 2023]. По-видимому, различия в определении концентрации галектина-9 в работах разных авторов в определенные сроки беременности обусловлены индивидуальными особенностями обследуемых, определением срока беременности и методами анализа, что требует проведения дальнейших исследований. Следует отметить, что иммуномодулирующая активность галектина-9 напрямую зависит от концентрации этого лектина. Так, в концентрациях, значительно превышающих его уровень при беременности, галектин-9 стимулирует синтез провоспалительного цитокина INF-гамма NK-клетками, тогда как в концентрациях, характерных для беременности, оказывает обратный эффект [Gleason et al., 2012]. У женщин со спонтанными абортами и другими патологиями беременности продукция галектина-9 клетками трофобласта и его уровень в периферической крови значительно снижается [Wu et al, 2015; Boron et al., 2022].
В целом, можно заключить, что галектин-9 играет важную роль в регуляции иммунореактивности организма при различных состояниях.
Структура рецептора Tim-3, механизмы сигнальной трансдукции
Основным лигандом для галектина-9 на NK-клетках является молекула Tim-3, которая экспрессируется на большинстве клеток иммунной системы: NK-клетках, CD4+ и CD8+Т-лимфоцитах, В-лимфоцитах, Treg, натуральных Т-клетках с функциями NK(NKT), моноцитах, макрофагах, тучных и дендритных клетках [Ndhlovu et al., 2012; Sun et al., 2016; Li et al., 2016].
NK-клетки конститутивно экспрессируют молекулы Tim-3, которые рассматривают как маркер их активации и созревания [Ndhlovu et al., 2012; Gleason et al., 2012; So et al., 2019], причем среди лимфоцитов NK-клетки имеют наибольшую экспрессию Tim-3 [Gleason et al., 2012; Ndhlovu et al., 2012]. У здоровых людей цитотоксические CD56dimCD16+NK-клетки как более зрелые демонстрируют повышенную экспрессию Tim-3 по сравнению с менее зрелыми регуляторными CD56brightCD16-NK-клетками [Ndhlovu et al., 2012]. В работе Ndhlovu с соавторами убедительно показано, что повышение экспрессии Tim-3 под влиянием IL-15 коррелирует с созреванием и трансформацией CD56brightCD16-NK в CD56dimCD16+NK [там же].
В отличие от Т-лимфоцитов, Tim-3-позитивные NK-клетки не являются истощенными или анергичными, а проявляют высокую функциональную активность в ответ на активирующие стимулы [там же]. Так, стимуляция NK-клеток цитокинами и их комбинациями (IL-12, IL-18, IL-12+IL-18, IL-15, IL-15+IL-12, IL-2) за исключением INF-гамма, связывание CD16 c Fc-фрагментами иммуноглобулинов, повышает экспрессию Tim-3 на обеих субпопуляциях NK-клеток [там же], что коррелирует с увеличением экспрессии транскрипционного фактора T-bet, который отвечает за дифференцировку наивных Т-лимфоцитов в Т-хелперы 1-го типа (T-box expressed in T-cells) [Gleason et al., 2012; So et al., 2019].
Tim-3 относится к гликопротеиновым рецепторам и имеет мембранную и растворимую форму [Chen et al., 2023]. Молекула Tim-3 включает экстрацеллюлярный, трансмембранный и интрацеллюлярный участки [там же]. Экстрацеллюлярный N–терминальный участок состоит из вариабельного иммуноглобулинового и гликозилированного муцинового доменов [там же]. Муциновый участок обогащен серином и треонином [там же]. Цитоплазматический С–терминальный домен содержит остатки тирозина, фосфорилирование которых играет критическую роль во внутриклеточной трансдукции сигнала [там же].
Галектин-9 взаимодействует с вариабельным иммуноглобулиновым доменом Tim-3 [там же]. Следует отметить, что сигналинг с Tim-3 изучен, главным образом, на Т-лимфоцитах. В состоянии покоя интрацеллюлярный домен рецептора Tim-3 связан с протеином Bat3 (human leukocyte antigen B-associated transcript 3) [там же]. После взаимодействия Tim-3 с галектином-9 тирозиновая протеинкиназа ITK (interleukin-2-inducible T-cell kinase) запускает фосфорилирование остатков тирозина, что приводит к диссоциации Bat3 от интрацеллюлярного домена и запускает инфлюкс кальция в клетку, вызывая апоптоз Th1-лимфоцитов [Ocana-Guzman, Torre-Bouscoulet, Sada-Ovalle, 2016]. Лишь в единичных работах показано, что в NK-клетках трансдукция сигнала с Tim-3 активирует MAPK (mitogen-activated protein kinase) и ERK (extracellular signal-regulated kinase) -сигнальные пути и вызывает деградацию IkBa – ингибитора для транскрипционного фактора NF-kB [Gleason et al., 2012]. Другими авторами показано, что регуляторные эффекты галектин-9-Tim-3-сигналинга на NK-клетках зависят от JNK- и AKT-сигнальных путей и могут быть реализованы путем регуляции активности транскрипционных факторов Id2, Blimp-1, Gata-3 и Eomes [Li et al., 2017].
В большинстве работ убедительно показано, что взаимодействие галектина-9 с Tim-3 на NK-клетках угнетает их цитотоксичность, ограничивает продукцию провоспалительных цитокинов INF-гамма, фактора некроза опухоли-альфа (TNF-альфа), синтез и секрецию перфорина и гранзима [Sun et al., 2016; Li et al., 2016, 2017]. В экспериментах по изучению фибросаркомы у мышей показано, что в присутствии галектин-9–экспрессирующих дендритных клеток NK-клетки проявляют повышенную цитотоксическую активность к опухолевым клеткам, оцениваемую по продукции INF-гамма, перфорина и гранзима [Nagahara et al., 2008]. Ранее были описаны примеры зависимости направленности иммунорегуляторных эффектов от концентрации галектина-9 [Gleason et al., 2012]. Таким образом, плейотропность регуляторных эффектов при активации галектин-9/Tim-3-сигнального пути объясняется многообразием вовлекаемых каскадов вторичных мессенджеров в разных типах клеток, а также условиями активации и факторами микроокружения.
В циркуляции присутствует также растворимая форма (s)Tim-3 [Grossman et al., 2021]. Предполагается, что это продукт деградации мембранной формы рецептора под действие металлопротеиназ [там же]. sTim-3 конкурирует с мембранным рецептором за связывание с галектином-9 [там же]. sTim-3 участвует в регуляции цитотоксичности CD8+Т-лимфоцитов и NK-клеток [Lu et al., 2021]. Уровень sTim-3 повышается при беременности по сравнению с небеременными [Wu et al., 2015; Grossman et al., 2021], что ассоциируется с повышенной экспрессией провоспалительных цитокинов (IFN-гамма и TNF-альфа) и снижением продукции IL-4 и IL-10 [Tang , Rangachari , Kuchroo, 2019]. Таким образом, предполагают, что sTim-3 участвует в поддержании баланса между про- и противовоспалительными иммунными реакциями у беременных женщин.
Tim-3 также является рецептором для фосфатидилсерина, который также связывается с его иммуноглобулиновым доменом [Chen et al., 2023]. Известно, что этот сигнальный путь участвует в регуляции фагоцитарной активности моноцитов, макрофагов, дендритных клеток при клиренсе апоптотирующих клеток [Ocana-Guzman, Torre-Bouscoulet, Sada-Ovalle, 2016; Chen et al., 2023]. Известны и другие лигадны для Tim-3 [Zhu et al., 2005; Wu et al., 2015].
Дизрегуляция экспрессии Tim-3 ассоциируется с развитием аутоиммунных, инфекционных, аллергических, онкологических заболеваний, отторжением трансплантатов [Wang et al., 2008; Han et al., 2013]. При различных типах солидных опухолей экспрессия Tim-3 в опухоль-инфильтрирующих лимфоцитах сопряжена с плохим прогнозом и низкой выживаемостью. При аутоиммунных патологиях экспрессия Tim-3 является маркером тяжести заболевания и эффективности проводимой терапии [Chen et al., 2023].
Галектин-9 также может взаимодействовать с другими мембранными рецепторами (CD137, CD44, PD-1 (programmed cell death protein 1)), некоторыми молекулами адгезии и внутриклеточными молекулами [Chen et al., 2023]. Показано, что взаимодействие галектина-9 с PD-1-рецептором, также, как и с β 3-интегрином (CD61) на CD4-лимфоцитах, способствует проникновению ВИЧ в клетки [Elahi et al., 2012], тогда как галектин-9/Tim-3-сигналинг уменьшает чувствительность CD4-лимфоцитов к проникновению ВИЧ, поскольку угнетает экспрессию корецепторов CXCR4, CCR-5, необходимых для проникновения вируса в клетку и репликацию его в клетке [там же].
В целом можно заключить, что галектин-9-Tim-3-сигнальный путь является важнейшим регуляторным механизмом для большинства клеток иммунной системы. На сегодняшний день молекула Tim-3 рассматривается как одна из перспективных мишеней в иммунотерапии, о чем свидетельствует увеличение количества клинических исследований с применением ингибиторов Tim-3 при различных патологиях.
Галектин-9/Tim-3-опосредованная регуляция функций NK-клеток в аспекте беременности
NK-клетки относятся к лимфоидным клеткам врожденного иммунитета. Популяция NK-клеток периферической крови гетерогенна. Большая часть NK-клеток периферической крови обладает выраженной цитотоксической активностью и имеет фенотип CD16+CD56dimNK (до 90%). Регуляторные CD16-CD56brightNK составляют менее 10% от общего числа NK-клетки. В исследованиях последних лет описывают также минорную субпопуляцию цитотоксических NK-клеток с фенотипом CD16+CD56-NK, которые характеризуются как истощенные [Cocker et al., 2022, Orlova et al., 2023a]. NK-клетки периферической крови распознают вирусинфицированные, опухолевые, поврежденные клетки собственного организма, чужеродные клетки и лизируют их путем контактного взаимодействия (Fas-Fas-L, PD-PD-L) и индукции апоптоза, секретируя гранулы, содержащие литические продукты (перфорин, гранзимы) [Erlebacher et al., 2013].
В процессе беременности плацента продуцирует большое количество гормонов и других биологически активных соединений, что приводит к изменению гормонального фона, метаболизма, иммунореактивности организма матери, необходимых для успешного протекания беременности и родов. При этом количество NK-клеток периферической крови существенно не меняется [Li et al., 2017; Orlova et al., 2023a], но уровень цитотоксических NK-клеток и их функциональная активность снижаются, а число регуляторных NK-клеток, напротив, увеличивается [там же]. По результатам мета-анализа имеющихся в литературе данных женщины с высоким риском спонтанных абортов и бесплодием характеризуются увеличением количества NK-клеток в периферической крови и избыточной продукцией провоспалительных цитокинов (IFN-гамма, TNF-альфа) [Miko et al., 2013; Li et al., 2016], тогда как уровень регуляторных NK-клеток является маркером благополучного исхода беременности [Li et al., 2016; Dixit, Karande, 2020].
Регуляторные NK-клетки периферической крови на ранних сроках беременности мигрируют в зону фетоплацентарного контакта, где трансформируются в децидуальные, которые являются основными эффекторами формирования неспецифической иммунной толерантности на границе раздела мать-плод, а также выполняют фетотрофическую функцию [Koopman et al., 2003; Cerdeira et al., 2013; Fu et al., 2017; Song Yan, 2022]. Они обнаруживаются в непосредственной близости к спиральным артериям плаценты, и, продуцируя факторы роста сосудов (vascular endothelial growth factor (VEGF), инсулиноподобного фактора роста (insulin-like growth factor-binding protein 1, IGFBP-1), плацентарного фактора роста (placental growth factor, PLGF), плейотропина, моделируют рост спиральных артерий и инвазию [там же]. Децидуальные NK-клетки путем контактного взаимодействия и секреции толерогенных цитокинов (TGF-бета1, IL-10) индуцируют формирование адаптивных (индуцибельных) iTreg из наивных CD4+T-лимфоцитов, препятствуя образованию регуляторных Т-лимфоцитов, продуцирующих IL-17 (Th17), которые активируют провоспалительные клеточно-опосредованные реакции [Fu et al., 2017]. Treg также подавляют цитотоксические реакции против трофобласта в зоне фетоплацентарного контакта [Koopman et al., 2003; Cerdeira et al., 2013; Fu et al., 2017; Song Yan et al., 2022]. Хотя в случае инфекции децидуальные CD56brightNK сохраняют способность развивать цитотоксический ответ против патогенов и выполнять защитную функцию [Crespo et al., 2020]. Провоспалительные цитокины IFN-gamma и TNF-alpha, которые продуцируются децидуальными NK-клетками при физиологически протекающей беременности, также участвуют в ангиогенезе [Erlebacher et al., 2013]. Однако при спонтанных абортах доля децидуальных NK-клеток, продуцирующих IFN-гамма и TNF-альфа, увеличивается, что свидетельствует о важной роли этих клеток в регуляции иммунореактивности. Поэтому изучение принципов регуляции функциональной активности NK-клеток значимо для понимания механизмов формирования иммунной толерантности, реализуемых организмом как для сохранения беременности, так и при развитии онкологических процессов, трансплантации тканей.
При беременности Tim-3 экспрессируется на всех субпопуляциях NK-клеток периферической крови: регуляторных CD16-CD56brightNK, цитотоксических CD16+CD56dimNK, цитотоксических истощенных CD16+CD56-NK и децидуальных NK [Li et al., 2016; Orlova et al., 2023b]. При этом в I триместре беременности экспрессия Tim-3 увеличивается в периферической крови только на регуляторных NK (CD16-CD56bright), а в III триместре – на всех субпопуляциях NK-клеток [Orlova et al., 2023a]. В работах других авторов показано, что цитотоксические CD56dimNK-клетки периферической крови демонстрируют повышенную экспрессию Tim-3 по сравнению с CD56brightNK в ранние сроки беременности [Li et al., 2016]. На децидуальных NK-клетках экспрессия Tim-3 также увеличивается в ранние сроки беременности [там же].
Галектин-9/Tim-3-сигналинг играет критическую роль в регуляции функций NK-клеток, включая дегрануляцию, цитотоксичность, выработку про- и противовоспалительных цитокинов, ангиогенных факторов при беременности [Gleason et al., 2012; Ndhlovu et al., 2012; Sun et al., 2016; Li et al., 2016]. Tim-3+NK и Tim-3-NK беременных имеют разный цитокиновый профиль. Tim-3+NK-клетки продуцируют преимущественно противовоспалительные цитокины (TGF-бета 1, IL-10 и IL-4), и в незначительных количествах INF-гамма и TNF-альфа [Meggyes et al. 2014; Li et al., 2016]. Tim-3+NK периферической крови по сравнению с Tim-3-NK имеют сниженную цитотоксическую активность по отношению к клеткам трофобласта и способны эффективно индуцировать пролиферацию Treg [ там же ]. Tim-3-позитивные децидуальные NK также проявляют толерогенные свойства, а именно продуцируют противовоспалительные цитокины (TGF-бета1, IL-10 и IL-4) и факторы роста сосудов, но в незначительных количествах TNF-альфа и перфорин [Li et al., 2016].
В исследованиях in vitro были изучены механизмы иммуномодулирующей активности галектина-9 в отношении NK-клеток. Показано, что инкубация NK-клеток, выделенных из периферической крови небеременных, с галектином-9 в концентрации, характерной для беременности, снижает процент цитотоксических CD16+CD56dimNK в культурах, но не влияет на число регуляторных CD16-CD56brightNK и цитотоксических истощенных CD16+CD56-NK клеток [Orlova et al., 2023b]. При этом галектин-9 увеличивает процент Tim-3-позитивных регуляторных клеток CD16-CD56brightNK и не влияет на количество цитотоксических Tim-3-позитивных CD16+CD56dimNK и CD16+CD56-NK-клеток [Orlova et al., 2023b]. Внесение Tim-3-блокирующих антител отменяло эффекты галектина-9 на трансформацию фенотипа и экспрессию Tim-3 в разных субпопуляциях NK [Orlova et al., 2023b]. Другими авторами также было показано, что галектин-9 влияет на формирование регуляторного фенотипа децидуальных NK, повышая экспрессию Tim-3 на них [Sun et al., 2016; Li et al., 2016]. Суммируя все вышесказанное можно заключить, что при беременности модулирующие эффекты галектина-9 реализуются, главным образом, через Tim-3-зависимые механизмы.
Помимо этого, показано, что Th2-цитокины – IL-4, а также низкие концентрации прогестерона, характерные для первого триместра беременности, эффективно усиливают экспрессию Tim-3 на NK-клетках периферической крови, вовлекая сигналинг с участием STAT6 транскрипционного фактора [Li et al., 2016]. Эстрадиол и хорионический гонадотропин в концентрациях, характерных для разных триместров беременности, подобных эффектов не оказывали [там же]. Хотя в наших исследованиях показано, что инкубация NK периферической крови небеременных женщин in vitro c гормонами, продуцируемыми плацентой (эстрадиол, эстриол, прогестерон, хорионический гонадотропин, лептин, грелин, кисспептин), в концентрациях, характерных для разных триместров беременности, эффективно влияет на трансформацию фенотипа, экспрессию молекул клеточной адгезии, продукцию перфорина и гранзима, цитокинов, а также регуляторных микро-РНК [Shirshev et al., 2017a,b; Orlova et al., 2023a,b,c]. По-видимому, гормонозависимые механизмы в Tim-3-опосредованной регуляции функций NK-клеток при беременности будут изучены в ближайшем будущем.
В исследованиях многих авторов убедительно показано, что концентрация галектина-9 и количество Tim-3-позитивных NK-периферической крови являются важным прогностическим маркером развития патологий беременности. Так, у женщин со спонтанными абортами количество Tim-3+NK снижается, а цитотоксичность и продукция провоспалительных цитокинов NK-клетками, напротив, увеличивается [Miko et al., 2013]. При проведении экспериментах на мышах убедительно показано, что блокада Tim-3 способствует резорбции плода, снижению количества Tim-3+NK-клеток периферической крови и нарушению их регуляторной функции, а адоптивный перенос Tim-3+NK-клеток, напротив, снижает частоту выкидышей [Li et al., 2016]. Суммируя вышеописанные данные литературы, можно заключить, что галектин-9 регулирует экспрессию молекулы Tim-3 и формирование регуляторного фенотипа NK-клеток как in vivo, так и in vitro, а галектин-9-Tim-3-сигналинг необходим для созревания NK-клеток и играет важную роль в сохранении и успешном протекании беременности. Молекулярные механизмы галектин-9/Tim-3-регуляции функций NK-клеток в аспекте беременности представлены на рисунке
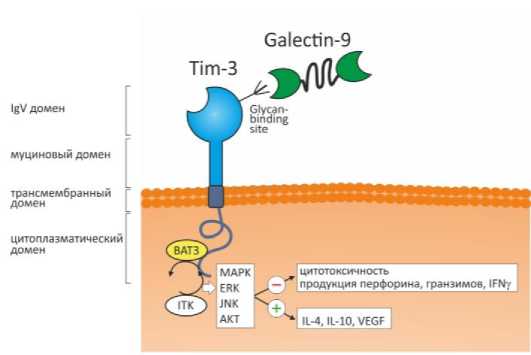
Молекулярные механизмы галектин-9/Tim-3-регуляции функций NK-клеток при беременности
[Molecular mechanisms of Galectin-9/Tim-3-regulation of functional activity of natural killers during pregnancy]
Из схемы видно, что галектин-9 (Galectin-9) взаимодействует с гликан-связывающим сайтом (Glycan-binding site) вариабельного иммуноглобулинового домена (IgV) рецептора Tim-3 на NK-клетках. Молекула Tim-3 включает экстрацеллюлярный (IgV и муциновый домены), трансмембранный и интрацеллюлярный участки. В состоянии покоя интрацеллюлярный домен рецептора Tim-3 связан с протеином Bat3 (human leukocyte antigen B-associated transcript 3). После взаимодействия Tim-3 с галектином-9 тирозиновая протеинкиназа ITK (interleukin-2-inducible T-cell kinase) запускает фосфорилирование остатков тирозина, что приводит к диссоциации Bat3 от интрацеллюлярного домена. Трансдукция сигнала с Tim-3 активирует MAPK (mitogen-activated protein kinase) и ERK (extracellular signal-regulated kinase) -сигнальные пути. Другими авторами показано, что регуляторные эффекты галектин-9-Tim-3-сигналинга на NK-клетках зависят от JNK- и AKT-сигнальных путей и могут быть реализованы путем регуляции активности транскрипционных факторов Id2, Blimp-1, Gata-3 и Eomes. При физиологически протекающей беременности галектин-9/Tim-3-сигналинг способствует формированию толерогенного фенотипа NK-клеток, снижая их цитотоксичность, ограничивая дегрануляцию и выброс литических факторов (перфоринов и гранзимов) и провоспалительного цитокина INF-гамма (INFy), способствуя продукции противовоспалительных цитокинов (IL-4, IL-10) и ангиогенных факторов VEGF (vascular endothelial growth factor).
Заключение
В целом, можно заключить, что галектин-9/Tim-3-сигналинг играет ключевую роль в регуляции функций NK-клеток. Направленность иммунорегуляторных эффектов галектина-9 на NK-клетки, реализуемых через Tim-3-рецептор, зависит от концентрации лектина, активационных стимулов, факторов микроокружения. При физиологически протекающей беременности галектин-9/Tim-3-сигналинг способствует формированию толерогенного фенотипа NK-клеток, снижая их цитотоксичность, ограничивая дегрануляцию и выброс литических факторов (перфоринов и гранзимов), способствуя продукции противовоспалительных цитокинов и пролиферации Treg, что необходимо для успешного протекания и сохранения беременности. Модулирование Tim-3-сигналинга на NK-клетках представляет новый терапевтический подход, перспективный не только для преодоления патологий беременности, но также и для лечения хронических вирусных, онкологических, аутоиммунных заболеваний.
Список литературы Галектин-9/Tim-3-зависимый механизм регуляции функциональной активности натуральных киллеров при беременности
- Рапопорт Е.М., Бовин Н.В. Специфичность галектинов человека в составе клеток // Биохимия. 2015. Т. 80, № 7. С. 1010–1022.
- Рапопорт Е.М., Курмышкина О.В., Бовин Н.В. Галектины млекопитающих: структура, углеводная специфичность и функции // Биохимия. 2008. Т. 73, № 4. С. 483–497.
- Asakura H. et al. Selective eosinophil adhesion to fibroblast via IFN-gamma-induced galectin-9 // J. Immunol. 2002. Vol. 169. P. 5912–5918.
- Boron D.G. et al. Galectin-1 and galectin-9 concentration in maternal serum: implications in pregnancies complicated with preterm prelabor rupture of membranes // Journal of Clinical Medicine. 2022. Vol. 11, № 21. Р. 6330.
- Cerdeira A.S. et al. Conversion of peripheral blood NK cells to a decidual NK-like phenotype by a cocktail of defined factors // J. of immunol. 2013. Vol. 190, № 8. P. 3939–3948.
- Chen H. et al. T-cell immunoglobulin and mucin-domain containing-3 (TIM-3): Solving a key puzzle in autoimmune diseases // Intern. immunopharmacol. 2023. Vol. 121. P. 110418.
- Cocker A.T.H. et al. CD56-negative NK cells: frequency in peripheral blood, expansion during HIV-1 infection, functional capacity and KIR expression. // Frontiers in Immunology. 2022. Vol. 13. P. 992723.
- Crespo A.C. et al. Decidual NK cells transfer granulysin to selectively kill bacteria in trophoblasts // Cell. 2020. Vol. 182, № 5. P. 1125–1139.
- Dixit A., Karande A.A. Glycodelin regulates the numbers and function of peripheral natural killer cells // J. Reprod. Immunol. 2020. Vol. 137. P. 102625.
- Elahi S. et al. Galectin-9 binding to tim-3 renders activated human CD4+ T cells less susceptible to HIV-1 infection // Blood. 2012. Vol. 119, № 18. P. 4192–204.
- Enninga E.A.L. et al. Immune checkpoint molecules soluble program death ligand 1 and galectin-9 are increased in pregnancy // Am. J. of Reprod. Immunol. 2018. Vol. 79, № 2. e12795.
- Erlebacher A. Immunology of the maternal-fetal interface // Annu. Rev. Immunol. 2013. Vol. 31. P. 387–411.
- Fu B. et al. Natural killer cells promote fetal development through the secretion of growth-promoting factors // Immunity. 2017. Vol. 47, № 6. P. 1100–1113.
- Fuselier C. et al. Placental galectins in cancer: why we should pay more attention // Cells. 2023. Vol. 12, № 3. P. 437.
- Gleason M.K. et al. Tim-3 is an inducible human natural killer cell receptor that enhances interferon gamma production in response to galectin-9 // Blood. 2012. Vol. 119, № 13. Р. 3064–3072.
- Grossman T.B. et al. Soluble T cell immunoglobulin mucin domain 3 (sTim-3) in maternal sera: a potential contributor to immune regulation during pregnancy // The J. of maternal-fetal & neonatal medicine: the official J of the Europ. Associat. of Perinat. Med. 2021. Vol. 34, № 24. P. 4119–4122.
- Han G. et al. Tim-3: an activation marker and activation limiter of innate immune cells // Front. Immunol. 2013. Vol. 4. P. 449–455.
- Imaizumi T. et al. Interferon-gamma stimulates the expression of galectin-9 in cultured human endothelial cells // J. Leukoc. Biol. 2002. Vol. 72. P. 486–491.
- Iwasaki-Hozumi H. et al. Blood levels of galectin-9, an immuno-regulating molecule, reflect the severity for the acute and chronic infectious diseases // Biomolecules. 2021. Vol. 11, № 3. P. 430.
- Koopman L.A. et al. Human decidual natural killer cells are a unique NK cell subset with immunomodulatory potential // The J. of exp. medicine. 2003. Vol. 198, № 8. P. 1201–1212.
- Li Y. et al. The Galectin-9/Tim-3 pathway is involved in the regulation of NK cell function at the maternal-fetal interface in early pregnancy // Cellular & molecul. Immunol. 2016. Vol. 13, № 1. Р. 73–81.
- Li Y. et al. Tim-3 signaling in peripheral NK cells promotes maternal-fetal immune tolerance and alleviates pregnancy loss // Science signaling. 2017. Vol. 10, № 498. P. 4323.
- Lu C. et al. An emerging role of TIM3 expression on T cells in chronic kidney inflammation // Front. Immunol. 2021. Vol. 12. P. 798683.
- Meggyes M. et al. Peripheral blood TIM-3 positive NK and CD8+T cells throughout pregnancy: TIM-3/Galectin-9 interaction and its possible role during pregnancy // PLoS One. 2014. Vol. 9, № 3. P. 92371.
- Miko E. et al. Involvement of Galectin-9/TIM-3 Pathway in the Systemic Inflammatory Response in Early-Onset Preeclampsia // PLoS One. 2013. Vol. 8, № 8. P. 71811.
- Nagahara K. et al. Galectin-9 increases Tim-3dendritic cells and CD8 T cells and enhances anti-tumor immunity via galection-9-Tim-3 interactions // J. Immunol. 2008. Vol. 181, № 11. P. 7660–7669.
- Ndhlovu L.C. et al. Tim-3 marks human natural killer cell maturation and suppresses cell-mediated cytotoxicity // Blood. 2012. Vol. 119, № 16. Р. 3734–3743.
- Ocana-Guzman R., Torre-Bouscoulet L., Sada-Ovalle I. TIM-3 regulates distinct functions in macrophages // Front. Immunol. 2016. Vol. 13, № 7. Р. 229.
- Orlova E.G. et al. Expression of TIM-3 and CD9 molecules on natural killer cells (NK) and T-lymphocytes with NK functions (NKT) in the peripheral blood at different trimesters of physiological pregnancy // J. Evol. Biochem. Phys. 2023a. Vol. 59. P. 809–821.
- Orlova E.G. et al. Galectin-9 influences the Tim-3 molecule expression in natural killer different subpopulations // Med. Immunol. (Russia). 2023b. Vol. 25, № 3. P. 469–476.
- Orlova E.G. et al. Hormone-Dependent Reprogramming of NK Cell Functions in the Aspect of Pregnancy // Science and Global Challenges of the 21st Century – Innovations and Technologies in Interdisciplinary Applications. Perm Forum 2022. Lecture Notes in Networks and Systems. 2023c. Vol. 622. Springer, Cham.
- Pełech A. et al. Do Serum galectin-9 levels in women with gestational diabetes and healthy ones differ before or after delivery? A pilot study // Biomolecules. 2023. Vol. 13, № 4. P. 697.
- Sanchez-Fueyo A. et al. Tim-3 inhibits T helper type 1-mediated auto- and allo- immune responses and promotes immunological tolerance // Nat. Immunol. 2003. Vol. 4, № 11. P. 1093–1101.
- Shirshev S.V. et al. Hormonal regulation of NK cell cytotoxic activity // Dokl. Biol. Sci. 2017b. Vol. 472, № 1. P. 28–30.
- Shirshev S.V. et al. MicroRNA in hormonal mechanisms of regulation of NK cell function // Dokl. Biochem. Biophys. 2017a. Vol. 474, № 1. P. 168–172.
- So E.C. et al. NK cell expression of Tim-3: first impressions matter // Immunobiology. 2019. Vol. 224, № 3. Р. 362–370.
- Song Yan et al. The mTORC1 signaling support cellular metabolism to dictate decidual NK cells function in early pregnancy // Front Immunol. 2022. Vol. 13. P. 771732.
- Sun J. et al. Tim-3 is up regulated in NK cells during early pregnancy and inhibits NK cytotoxicity toward trophoblast in galectin-9 dependent pathway // PloS one. 2016. Vol. 11, № 1. P. 0147186.
- Tang R., Rangachari M., Kuchroo V.K. Tim-3: A co-receptor with diverse roles in T cell exhaustion and tolerance // Semin. Immunol. 2019. Vol. 42. P. 101302.
- Wang F. et al. Activation of Tim-3–Galectin-9 pathways improves survival of fully allergenic skin grafts // Transpl. Immunol. 2008. Vol. 19, № 1. P. 12–19.
- Wu M. et al. Soluble co stimulatory molecule sTim3 regulates the differentiation of Th1and Th2 in patients with unexplained recurrent spontaneous abortion // Int. J. Clin. Exp. Med. 2015. Vol. 8, № 6. Р. 8812–8819.
- Zhu C. et al. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity // Nat. Immunol. 2005. Vol. 6, № 12. P. 1245–1252.
- Zhu W. et al. Tim-3: An inhibitory immune checkpoint is associated with maternal-fetal tolerance and recurrent spontaneous abortion // Clin. Immunol. 2022. Vol. 245. P. 109185.
