Гаплоидный партеногенез in vitro у сахарной свеклы (Beta vulgaris L.): факторы и диагностические признаки
: факторы и диагностические признаки")
Автор: Жужжалова Т.П., Подвигина О.А., Знаменская В.В., Васильченко Е.Н., Карпеченко Н.А., Землянухина О.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Селекция: культура тканей, молекулярный анализ
Статья в выпуске: 5 т.51, 2016 года.
Бесплатный доступ
Традиционное получение инбредных линий и гибридов в селекции сахарной свеклы -трудоемкий процесс, требующий длительного времени из-за 2-летнего цикла развития растений, само- и перекрестной несовместимости, инбредной депрессии. Для индукции генотипического многообразия в исходной популяции перспективны биотехнологии, в том числе гаплоидный партеногенез. Мы показали, что при индуцировании гаплоидов in vitro у сахарной свеклы ( Beta vulgaris L.) эффективна экспресс-диагностика по фенотипическим, эмбриологическим признакам, отражающим периоды развития цветоносных побегов, органов, бутонов, стадии формирования зародышевого мешка семязачатка и пыльцевых зерен. Регенерационная активность наблюдается в семязачатках с 1 по 25 бутоны, расположенных по колосовидному цветоносу плейохазия вверх от раскрывшегося цветка. Ядра и клетки женского гаметофита изолированных семязачатков этих бутонов в условиях in vitro способны к новообразованиям на всех этапах развития, но 8-ядерные или 7-клеточные зародышевые мешки наиболее компетентны к морфогенезу и переключению программы развития с гаметофитного на спорофитный путь. Наступление критического периода развития зародышевых мешков предварительно определяли по сопутствующим эмбриологическим признакам - наличию одноядерных микроспор и двух-трехклеточных пыльцевых зерен пыльников, находящихся с семязачатками в одном бутоне. Полученные нами результаты свидетельствуют, что гормональный состав питательной среды по Гамборгу (В5) служит важным фактором, который эффективно регулирует направление морфогенетического развития у изолированных семязачатков - через прямую регенерацию (эмбриоидогенез) или через каллус (гемморизогенез), что свидетельствует о тотипотентности как половых, так и соматических клеток экспланта. Полученные данные о воспроизведении in vitro гаплоидных регенерантов углубляют имеющиеся научные представления о специфике морфогенетического потенциала у растений сахарной свеклы. Стабилизирующий отбор при создании линий удвоенных гаплоидов способствует выявлению ценных морфологических признаков регенерантов. Определение числа хромосом и хлоропластов в замыкающих клетках устьиц, а также электрофоретическая подвижность изоферментов (по локусам Adh-1, Mdh-1, Mdh-2, Me-1, Idh-1, Idh-2, Gdh-1 ) могут служить маркерами при индуцировании гаплоидии и создании гомозиготных реституционных линий сахарной свеклы. Показана эффективность метода RFLP-анализа с использованием рестриктазы Hind III, позволившего впервые идентифицировать гаплоидные микроклоны по типу цитоплазмы. Молекулярные маркеры свидетельствовали, что регенеранты с нормальной цитоплазмой (N) имели один ПЦР-продукт длиной 800 п.н., у форм (S) с цитоплазматической мужской стерильностью (ЦМС) обнаруживались два продукта рестрикции (320 п.н. и 480 п.н.). Выявление гаплоидных регенерантов со стерильной цитоплазмой из исходных популяций имеет важное теоретическое и прикладное значение для селекции, облегчая задачу создания гомозиготных линий с ЦМС и высокопродуктивных гибридов на стерильной основе.
Сахарная свекла, гаплоидный партеногенез, женский гаметофит, удвоенные гаплоиды, эмбриоидогенез, органогенез, изоферменты, rflp-анализ днк
Короткий адрес: https://sciup.org/142213966
IDR: 142213966 | УДК: 633.63:631.52:581.3 | DOI: 10.15389/agrobiology.2016.5.636rus
Sugar beet (Beta vulgaris L.) haploid parthenogenesis in vitro: factors and diagnostic characters
Traditional obtaining of inbred lines and hybrids in sugar beet breeding requires a long time and is labour-consuming because of 2-year cycle of plant development, self- and cross-incompat-ibility, and inbreeding depression. To induce genotypic diversity in initial population, biotechnology methods including haploid parthenogenesis are promising. We have shown that, when inducing sugar beet ( Beta vulgaris L.) haploids in vitro, express-diagnostics using phenotypic and embryological characters that are representative of periods of flowering shoots, organs and buds development, and stages of embryo sac, ovule and pollen grains formation is effective. The regenerative activity is observed in ovules of bud 1 to bud 25 located on ear part of pleochasium upward from the open flower. The nuclei and cells of female gametophyte of isolated ovules under in vitro conditions are capable of neoplasm at all stages of development, but the 8-nuclear or 7-celled embryo sacs are the most appropriate to morphogenesis and switching of development program from gametophyte to sporophyte type. Critical period of embryo sac development has been beforehand determined from the accompanying embryological characters - the presence of single-nuclear and two-three-celled pollen grains of anthers located in the same bud as ovules. The results we obtained indicate that hormonal composition of the Gamborg's B-5 (B5) medium is an important factor that effectively regulates direction of morphogenetic development in isolated ovules through direct regeneration (embryoidogenesis) or via callus (hemorhizogenesis) that is the evidence of totipotency of both sexual and somatic cells in the explant. The obtained data on in vitro reproduction of haploid regenerants add to available scientific notion of morphogenetic potential specificity in sugar beet plants. Stabilizing selection used to produce double haploids promotes detection of valuable morphological features of the regenerants. Determination of chromosome and chloroplast numbers in stomata guard cells as well as isozyme electrophoretic mobility (for Adh-1, Mdh-1, Mdh-2, Me-1, Idh-1, Idh-2, Gdh-1 loci) can serve as markers when inducing haploidy and producing homozygous restitution lines of sugar beet. Efficiency of RFLP-analysis method using Hind III restrictase that has allowed for the first time to identify haploid microclones according cytoplasm type is shown. Molecular markers have indicated that regenerants with normal cytoplasm (N) have one PCR-product of 800 bp in length not digested by Hind III. Two fragments (320 bp and 480 bp) of 800 bp product digestion are found in cytoplasmic male sterile (CMS) forms (S) that reflects combination of recessive and dominant genes. Obtaining haploid regenerants with sterile cytoplasm from initial population is of great theoretical and practical importance for sugar beet breeding thus facilitating the problem of producing homozygous lines with CMS and high-productive hybrids on the sterile basis.
Текст научной статьи Гаплоидный партеногенез in vitro у сахарной свеклы (Beta vulgaris L.): факторы и диагностические признаки
Современный отечественный генофонд сахарной свеклы достаточен для создания сортов с требуемыми свойствами, однако традиционное получение инбредных линий и гибридизация длительны и трудоемки, что обусловлено 2-летним циклом развития растений, само- и перекрестной несовместимостью, инбредной депрессией (1). Биотехнологический метод гаплоидии, предполагающий культивирование растений в условиях in vitro (2), эффективен для индукции генотипического многообразия в исходной популяции, но его применение ограничено узкой специфичностью этих условий не только для разных генотипов одного вида, но и в зависимости от этапа культивирования. Варьирование, наблюдаемое при культивирова-
∗ Работа выполнена при поддержке гранта Президента РФ «Научная школа РФ» (НШ=5282.2014.4 — проект «Разработка теории репродукции растений с позиции проблемы целостности и надежности биосистем. Поливариантность морфогенетических программ развития, естественные и искусственные модели их реализации».
нии репродуктивных органов растений, может значительно расширить пределы изменчивости и облегчить получение форм для селекционных целей (в том числе на основе удвоенных гаплоидов) (3).
Для усовершенствования метода гаплоидного партеногенеза у сахарной свеклы оптимизируются условия индукции неоплодотворенных семязачатков и культивирования гаплоидных регенерантов с ценными генетическими рекомбинациями, отрабатываются приемы включения реституционных линий в селекционный процесс (4-8). Наиболее сложная проблема — гормональный состав питательных сред при культивировании in vitro (отметим, что принцип эмпирического подбора условий для индукции гаплоидии остается доминирующим) (9-10). Представляет интерес использование жидких сред, значительно повышающее выход гаплоидных регенерантов (11), спонтанная диплоидизация гаплоидов при длительном (более 1 года) культивировании (12), предобработка семязачатков агентами (колхицин, трифлуралин и др.), вызывающими антимитотическую активность клеток женского гаметофита и повышающими частоту гаплоидной регенерации (13). Сравнение морфологии гаплоидов и удвоенных гаплоидных регенерантов выявило преимущества первых по скорости развития побегов и размножения in vitro. На этом основании предлагается использовать гаплоиды сахарной свеклы не только для получения удвоенных гаплоидов, но и в специализированных проектах, касающихся геномного анализа, или при генетической трансформации, когда гаплоидные ткани служат исходным материалом (14).
У сахарной свеклы культивирование in vitro неоплодотворенных семязачатков и получение из них гаплоидов позволило направленно создавать гомозиготные линии для селекции (9, 15). Однако коммерческое применение этих технологий ограничивается низким выходом гаплоидных регенерантов и недостаточной методической разработанностью в том, что касается критических периодов в развитии зародышевого мешка, установления морфогенетической компетентности его элементов для переключения с гаметофитной на спорофитную программу развития, биохимической и молекулярно-генетической оценки создаваемых гомозиготных линий.
Мы впервые проанализировали признаки, связанные с морфогенезом и воспроизведением растений-регенерантов сахарной свеклы in vitro при гаплоидном партеногенезе, и показали возможность их использования в качестве диагностических для оценки критических периодов в развитии женского гаметофита при переходе на спорофитный путь морфогенеза. Кроме того, применение RFLP-анализа с рестриктазой Hind III впервые позволило выделять гаплоидные микроклоны по типу цитоплазмы (N или S) из сортовых популяций сахарной свеклы, облегчая задачу создания линий с ЦМС.
Цель работы — выявить признаки, позволяющие повысить эффективность гаплоидного партеногенеза у сахарной свеклы in vitro, в том числе в сочетании с биохимическими и молекулярными маркерами.
Методика . Для культивирования органов и тканей использовали гибриды Beta vulgaris L. (коллекция Всероссийского НИИ сахарной свеклы и сахара им. А.Л. Мазлумова). Применяли общепринятую технику стерилизации и приготовления питательных сред по Р.Г. Бутенко (2).
Активность фермента NADP-изоцитратдегидрогеназы определяли в гомогенатах растительных тканей (16). Электрофорез изоферментов в крахмальном геле и ПААГ проводили согласно описанию (17).
RFLP-анализ (restriction fragment length polymorphism) с Hind III (18) применяли для выявления стерильных и фертильных гаплоидных регенерантов. Продукты разделяли электрофорезом в 1 % агарозном геле с добавлением бромистого этидия.
Плоидность определяли на проточном цитофотометре Ploidy Analys- er PA-2 («Partec GmbH», Германия) (11).
Достоверность цитоэмбриологических данных оценивали с помощью коэффициента вариации Сv (19).
Результаты . При культивировании in vitro неоплодотворенных семязачатков сахарной свеклы была обнаружена уникальная способность эмбриогенных клеток к реализации потенциала развития под влиянием экзогенных факторов (20), что позволило разработать технологию получения гомозиготных линий. Благодаря этому приему генетически и морфологически разнообразный материал от обогащенных в генетическом отношении растений-доноров получают без многократного самоопыления. Предложенная технология обеспечивает выход до 38 гаплоидных регенерантов от одного растения-донора и создание гомозиготного материала сахарной свеклы за 2-3 года (вместо 8-10 лет при традиционных методах). В настоящее время свыше 100 таких гомозиготных линий используются в селекционных программах (20). Эксперименты выявили ряд морфологических, цитоэмбриологических, биохимических и молекулярно-генетических признаков, повышающих эффективность обнаружения гаплоидов при индукции гаплоидного партеногенеза in vitro, что ускоряет создание гомозиготных линий.
На этапе введения семязачатков в культуру к критическим факторам, влияющим на эффективность гаплоидной регенерации, относится период развития цветоносных побегов у донорских растений. У сахарной свеклы оптимальна фаза бутонизации в конце периода раннего генеративного состояния соцветий (VII и VIII этапы органогенеза) (21). Бутоны с семязачатками, более способные к индуцированию гаплоидии и максимально пригодные для эксплантации, находятся в центральном колосе кистевидной части плейохазия. Регенерационная активность наблюдается в семязачатках с 1-го по 25-й бутон (расположены по колосовидному цветоносу вверх от раскрывшегося цветка).
Для успешного культивирования семязачатков также необходимо выбрать оптимальную стадию развития зародышевого мешка. Цитоэмбрио-логические наблюдения показали, что ядра и клетки женского гаметофита у сахарной свеклы способны к новообразованиям in vitro на всех этапах развития. По-видимому, это связано с особенностями женского гаметофита, в первую очередь с хорошо выраженной полярностью, дифференцирующими делениями и тотипотентностью его элементов, что в условиях in vitro стимулирует инициацию программы формирования гаплоидных регенерантов. По нашим данным, поляризация начиналась в одноядерном зародышевом мешке, который по мере развития увеличивался в размере и вытягивался в направлении к микропиле. В результате последующих трех митотических делений образуется 8-ядерный зародышевый мешок. После последнего митоза от каждого полюса к центру отходит по одному полярному ядру. Вокруг ядер локализуется цитоплазма и происходит цитокинез, который завершается образованием семи клеток зародышевого мешка (22).
Культивирование изолированных семязачатков, содержащих 8-ядерные (или 7-клеточные, но 8-ядерные) зародышевые мешки, наиболее благоприятно для индуцирования гаплоидии и регенерации (8). Можно предположить, что строгая полярность зародышевого мешка в этот период, а также тотипотентность его ядер и неспециализированных клеток женского гаметофита обеспечивают наибольшую способность к морфогенезу. Это критический фактор для переключения на спорофитный тип развития.
Определение стадий развития зародышевого мешка — сложный и длительный процесс. Здесь критический период диагностируется по эмбриологическим признакам — наличию одноядерных микроспор и 2-3638
клеточных пыльцевых зерен пыльников, находящихся в одном бутоне с семязачатками. В зависимости от погодных условий в период цветения эти признаки можно наблюдать за 1-5 сут до раскрытия бутонов.
Диагностическими признаками также могут служить аномалии мужского и женского гаметофитов, которые проявлялись у некоторых генотипов, полученных при длительном самоопылении и полиплоидии, а также у ЦМС-гибридов, где с частотой до 40 % встречался апомиксис. Возможно, рецессивные гены, вовлеченные в контроль апомиксиса (23), в том числе в гаплоидный партеногенез, по-разному накапливаются при селекции. Генотипы с наибольшими аномалиями более склонны к индукции гаплоидии in vitro.
Существенный недостаток гаплоидной технологии in vitro — низкая частота регенерации (1,7-10,5 %). Ее повышают стрессовые воздействия, например предобработка семязачатков холодом (4-6 ° С, 2-4 сут) или рентгеновским облучением (1000-5000 Р, 50 мин) (24). Возможно, in vitro у семязачатков сахарной свеклы (по аналогии с микроспорами пшеницы) (25) низкие положительные температуры, изменяя метаболизм, задерживают развитие зародышевого мешка и индуцируют спорофитный морфогенез.
Гормональный состав сред при культивировании семязачатков тоже был ключевым фактором, определяя тип морфогенеза — через регенерацию (эмбриоидогенез) или через каллус (гемморизогенез) (26).
В наших опытах индукцию и морфогенез гаплоидных регенерантов по типу спорофита лимитировала среда на основе минеральных солей по Гамборгу (В5) с витаминами по Уайту и различным гормональным составом (26). Так, добавление гиббереллина (2 мг/л) индуцировало эмбриоидо-генез гаплоидных регенерантов. В зародышевом мешке семязачатка с первых дней культивирования наблюдали деление клеток яйцевого аппарата с образованием уже на 3-и сут многоклеточного (5-8 ярусов клеток) про-эмбрио. К 5-м сут зародыш принимал форму шара и увеличивался в размере за счет делений в поперечном и продольном направлениях, заполняя на 8-12-е сут все пространство зародышевого мешка (при этом гаусториаль-ный вырост в нуцеллус не увеличивался). Центральная клетка, не приступая к делению, сохранялась до 5-8-х сут, затем дегенерировала. Иногда формировалось несколько ядер эндосперма, которые также дегенерировали. Крахмал в клетках нуцеллуса не откладывался, перисперм и оболочки семени не формировались. На 28-е сут зародыш разрывал интегументы и проросток появлялся на поверхности семязачатка с образованием небольших семядольных листочков, гипокотиля и первичного корешка. Это свидетельствует о сходстве гаплоидного эмбриоидогенеза и эмбриогенеза зиготического зародыша, однако эмбриоид формируется только с использованием минеральных солей среды культивирования (без участия питательных веществ эндосперма, который в условиях in vitro вообще не образуется).
Ауксины стимулировали рост каллусных структур наряду с гиноге-нетическими зародышами. Гиббереллин (2 мг/л), 6-бензиламинопурин (0,1 мг/л) и индолил-3-масляная кислота (0,1 мг/л) индуцировали развитие гаплоидных эмбриоидов, а затем вторичных регенерантов из каллусных тканей гипокотиля, что 6-10-кратно увеличивало выход гаплоидов (20). Поскольку тотипотентность клеток гетерогенного каллуса неодинакова, возможно формирование двух основных типов каллуса — морфогенного и неморфогенного. Неморфогенный состоит из паренхимных клеток и не регенерирует. В морфогенном каллусе (имел более плотную мелкозернистую структуру) происходили формообразовательные процессы — либо ризогенез, либо гем-могенез (появление почек), либо гемморизогенез. Гемморизогенез начинался в субэпидермальном слое каллуса. В условиях in vitro появлялись инициаль- ные клетки с утолщенной клеточной оболочкой и более крупным ядром, из которых de novo развивались зоны меристемы (в виде бугорков). В результате периклинальных делений на внешней стороне бугорков формировался апекс (серповидный валик) и примордий 1-го листа, то есть ростовая почка. Апексы корней закладывались в боковой или базальной части каллуса. Между корнем и почкой формировался проводящий пучок. Воспроизведение растений через каллус происходило при гемморизогенезе или геммоге-незе с последующей индукцией корневой системы по истечении 3-4 нед.
Культивирование неоплодотворенных семязачатков в жидкой питательной среде усиливало пролиферацию клеток женского гаметофита, длительно (до 4-6 мес) сохраняя их жизнеспособность. Перенос этих эксплантов на агаризованную среду того же состава стимулировал регенерацию (на 13,7 и 18,9 % — через каллусо- и эмбриоидогенез). Отметим, что каллус — система с непрямым органогенезом, что увеличивает время получения регенерантов, поэтому прямая регенерация in vitro более перспективна для селекции.
Этап стабилизации включает отбор нормально развитых гаплоидных растений-регенерантов (27) с высокой способностью к формированию адвентивных побегов и микроклональному размножению. Для повышения эффективности на этом этапе использовали морфологические и цитологические признаки. Так, у гаплоидных регенерантов высота и размер всех органов обычно меньше, чем у диплоидов. У нормально развитых гаплоидных форм в зависимости от генотипа более узкие листовые пластинки с длинными черешками или, наоборот, широкие листья с волнистым краем и короткими черешками. Характерный цитологический признак гаплоидов — 9 хромосом, которые выявляются при микроксопировании или цитофотометрически по содержанию ядерной ДНК (28). Последний метод быстрее, надежнее и может широко применяться для выявления растений с измененной плоидностью. К цитоэмбриологическим признакам следует отнести наличие 8-10 хлоропластов в замыкающих клетках устьиц, среди которых встречаются аномальные или не формирующие хлоропласты либо имеющие одну бобовидную клетку. Так как цитоэмбриологические признаки у гаплоидов варьировали незначительно, их можно использовать в качестве морфологических маркеров для идентификации и отбора гаплоидных регенерантов. Это позволяет проводить оценку на самых ранних этапах развития регенерантов, когда анализ хромосомного набора крайне затруднен и приводит к гибели эксплантов.
При колхицинировании гаплоидов (27) происходила полиплоидиза-ция с образованием диплоидных, триплоидных и тетраплоидных форм. При повышении плоидности увеличивалось число хлоропластов в паре замыкающих клеток устьиц: с 9-11 у гаплоидов до 12-14, 15-17 и 18-21 соответственно у ди-, три- и тетраплоидов. Изменялось и соотношение площади замыкающих клеток у гаплоидных и ди-, три и тетраплоидных форм — соответственно 1:1,56; 1:1,87; 1:2,68 (табл. 1).
1. Цитоморфологические характеристики пар замыкающих клеток устьиц у растений сахарной свеклы ( Beta vulgaris L.) при разной плоидности
|
Плоидность |
Просмотрено пар клеток |
Число хлоропластов, шт. |
Cv , % |
Площадь, мкм2 |
Cv , % |
|
|
среднее |
min-max |
|||||
|
Гаплоидный (½) |
250 |
10,2 |
9,4-10,9 |
9,3 |
0,95 |
0,9 |
|
Дигаплоидный (2½) |
250 |
13,2 |
11,8-14,5 |
9,7 |
1,48 |
1,6 |
|
Триплоидный (3½) |
250 |
15,5 |
14,4-16,6 |
5,8 |
1,78 |
1,8 |
|
Тетраплоидный (4½) |
250 |
19,7 |
18,5-20,9 |
6,4 |
2,55 |
1,3 |
Отметим, что у некоторых удвоенных гаплоидов при размножении (in vitro и в грунте) восстанавливалось гаплоидное состояние, что, возможно, объясняется более коротким митотическим циклом у гаплоидных регенеран-640
тов. Поэтому при колхицинировании и внутриклеточном отборе возможно увеличение доли гаплоидных клеток в миксоплоидной меристеме регенерантов, что отрицательно сказывается на диплоидизации. Для усиления эффекта диплоидизации гаплоидов in vitro мы изменяли гормональный состав питательной среды, добавляя цитокинины и гиббереллин, тормозящие рост гаплоидной ткани. Высокой способностью стимулировать клеточное деление у диплоидов и тормозить — у гаплоидов обладал кинетин при добавлении до 0,25 мг/л, что позволяло получать до 90 % регенерантов с постоянным диплоидным набором хромосом.
Удобным инструментом выявления гаплоидных регенерантов сахарной свеклы служит сравнение изоферментных спектров. При исследовании электрофоретической подвижности изоформ NADP-изоцитратдегидрогеназы (NADP-ИДГ, КФ 1.1.1.42) нами обнаружены отличия гаплоидных регенерантов от исходного диплоидного растения. Ранее мы показали присутствие двух изоформ NADP-ИДГ — цитоплазматической ( ∼ 95 % активности) и митохондриальной ( ∼ 5 % активности) в листьях контрольных растений, что согласуется с результатами электрофореза: у контрольных вариантов имелись две изоформы NADP-ИДГ (Rf 0,31 и 0,39) с полосами одинаковой интенсивности, а у гаплоидов фермент был представлен двумя зонами активности — мономорфной (Rf 0,37) и полиморфной (Rf 0,26 и 0,31).
Изоферментный анализ можно также использовать для определения степени гомозиготности формируемых в результате стабилизирующего отбора реституционных линий. Теоретически все линии удвоенных гаплоидов должны быть гомозиготными. Однако у реституционных линий, полученных после обработки колхицином, выявляется в среднем 4 % гетерозиготных локусов (29). Не исключено, что такой полиморфизм можно объяснить эпигенетической изменчивостью у удвоенных гаплоидов сахарной свеклы (30, 31). Индекс изоферментной гомозиготности (Iiz) в проанализированных линиях, определенный как средний процент гомозиготности по семи изофер-ментным локусам, варьировал от 0,81 до 1 (29) и в среднем составил 0,96 (табл. 2). Вероятно, гомозиготность этих реституционных линий достоточна высока, несмотря на возможные эпигенетические изменения.
2. Показатели изоферментной гомозиготности у реституционных линий сахарной свеклы ( Beta vulgaris L.)
|
Линия |
Доля гомозигот по изоферментным локусам, % |
Индекс изоферментной |
||||||
|
Adh-1 1 |
Mdh |
1 1 Mdh-2 1 |
Me-1 |
Idh-1 1 |
Idh-2 |
1 Gdh-1 |
гомозиготности Iiz |
|
|
1 |
100 |
87 |
93 |
88 |
100 |
100 |
100 |
0,95 |
|
2 |
100 |
97 |
100 |
68 |
100 |
100 |
96 |
0,94 |
|
3 |
100 |
40 |
93 |
42 |
100 |
100 |
93 |
0,81 |
|
4 |
100 |
87 |
93 |
100 |
100 |
100 |
100 |
0,97 |
|
5 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
1,00 |
|
6 |
100 |
100 |
100 |
96 |
100 |
96 |
100 |
0,99 |
|
7 |
100 |
100 |
100 |
96 |
93 |
96 |
100 |
0,98 |
|
8 |
97 |
79 |
84 |
96 |
100 |
100 |
100 |
0,94 |
|
Среднее |
99 |
86 |
95 |
98 |
99 |
99 |
99 |
0,96 |
Молекулярные маркеры нейтральны по отношению к фенотипу, нетканеспецифичны, их можно обнаружить на любой стадии развития растений. Они позволяют контролировать передачу генетического материала от растений-доноров и вести отбор на искомый селекционный признак, например на цитоплазматическую мужскую стерильность (32, 33). Известно, что в популяциях сахарной свеклы присутствуют растения с нормальной (N) и стерильной (S) цитоплазмой. У N-растений пыльца фертильна и жизнеспособна, у S-растений она может быть как фертильной, так и стерильной в зависимости от взаимодействия стерильной (S) цитоплазмы с рецессивными генами ядра rf1 и rf2. Стерильность цитоплазмы у сахарной свеклы обусловлена изменением нуклеотидной последовательности в митохондриальном и хлоропластном геномах (34, 35).
Наши исследования показали, что PCR и RFLP-анализ с использованием рестриктазы Hind III позволяет идентифицировать тип цитоплазмы у создаваемых гаплоидов по числу рестриктов. У гаплоидных микроклонов с нормальной и стерильной цитоплазмой амплифицировался один фрагмент (800 п.н.), который рестрицировался только у стерильных (S) форм (рис.; видны два рестрикта и остатки фрагмента 800 п.н.).
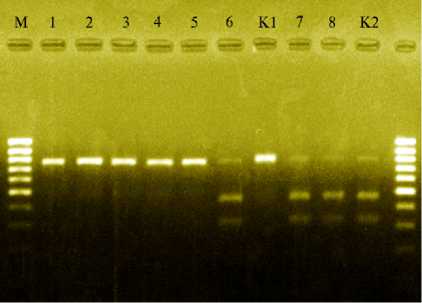
Электрофореграмма рестриктов ампли-фицированных фрагментов ДНК (RFLP-анализ, рестриктаза Hind III ) у гаплоидных растений-регенерантов сахарной свеклы ( Beta vulgaris L.): К1 — контрольные фертильные растения, К2 — контрольные стерильные растения; 1-5 — формы с нормальной (N) цитоплазмой, 6-8 — формы со стерильной (S) цитоплазмой; М — маркеры молекулярных масс (ДНК-маркер MassRuler™, 80-1031 п.н., SM0383, «Thermo Scientific», США).
Гаплоиды, у которых этот фрагмент не рестрицировался Hind III, были представлены полностью фертильными формами с нормальной цитоплазмой (N) и ядерными генами в рецессивном состоянии ( rf ). В остальных образцах наблюдался полиморфизм фрагментов, что, по-видимому, предполагает наличие у соответствующих гаплоидных форм стерильной цитоплазмы (S) и разное сочетание рецессивных и доминантных аллелей ядерных генов Rf1 / rf1 и Rf2 / rf2 . Отметим, что PCR-профили у всех стерильных регенерантов (как гаплоидов, так и удвоенных гаплоидов) одинаковы. Поэтому выявление растений-регенерантов со стерильной цитоплазмой на разных этапах культивирования представляет значительный интерес для селекции сахарной свеклы, облегчая создание линий с ЦМС и высокопродуктивных гибридов на стерильной основе. Таким образом, показана эффективность использования комплекса диагностических признаков для совершенствования метода гаплоидного партеногенеза in vitro у сахарной свеклы. Выявленные маркерные фенотипические, морфологические, цитоэмбриологические показатели надежно характеризуют критические периоды для развития генеративных органов растений-доноров и индукции гаплоидии, а также стадии морфогенеза in vitro, наиболее благоприятные при стабилизирующих отборах гаплоидов и удвоенных гаплоидов. Определение электрофоретической подвижности изоферментов и молекулярное маркирование позволяют оценивать степень гомозиготности созданных линий. Использование RFLP-анализа дает возможность вести отбор гаплоидных регенерантов на искомый селекционный признак, например цитоплазматическую мужскую стерильность (ЦМС), и формировать линии, гомозиготные по ЦМС.
Список литературы Гаплоидный партеногенез in vitro у сахарной свеклы (Beta vulgaris L.): факторы и диагностические признаки
- Буренин В.И. Генетические ресурсы рода Beta L. (Свекла). СПб, 2007.
- Бутенко Р.Г. Биология клеток высших растений in vitro и биотехнологии на их основе. М., 1999.
- Baenziger P.S., Russell W.K., Groef G.L., Campbell B.T. Improving lives: 50 years of crop breeding genetics and cytology. Crop Sci., 2006, 46: 2230-2244 ( ) DOI: 10.2135/cropsci2005.11.0404gas
- Atanassov A.I. Method for continuous bud for mation in tissne culture of sugar beet (Beta vulgaris L.). Pflazenzuchtg, 1980, 84: 23-29.
- Slavova J. Effective method for sugar beet haploids obtained from unpollinated ovules. Biotechnology & Biotechnological Equipment, 2014, 7(2): 34-35 ( ) DOI: 10.1080/13102818.1993.10818689
- Chen J.F., Cui L., Malik A.A., Mbira K.G. In vitro haploid and diploid production via unfertilized ovule culture. Plant Cell Tiss. Organ Cult., 2011, 104(3): 311-319 ( ) DOI: 10.1007/s11240-010-9874-6
- Murovec J., Bohanec B. Haploids and doubled haploids in Plant breeding. In: Plant breeding/I. Abdurakhmonov (ed.). InTech d.o.o., Croatia, 2012: 87-106 (doi: 10.5772/1389).
- Знаменская В.В., Жужжалова Т.П., Подвигина О.А. Индукция гиногенеза у сахарной свеклы. Мат. Российского симпозиума «Новые методы по биотехнологии растений». Пущино, 1993: 111.
- Подвигина О.А. Морфогенетическое развитие эксплантов сахарной свеклы в условиях in vitro. Сахарная свекла, 2003, 10: 22-24.
- Nagl N., Mezei S., Kovačev L., Vasić D., Čačić N. Induction and micropropagation potential of sugar beet haploids. Genetica, 2004, 36(3): 187-194 ( ) DOI: 10.2298/GENSR0403187N
- Sliwinska E., Pedersen H. Flow cytometric analsis of nuclear replication stages during germination of sugar-beet seeds different in vigour. Electronic Journal of Polish Agricultural Universities, 1999, 2(1).
- Svirshchevskaya A.M., Doležel J. Production and performance of gynogenetic sugarbeet lines. Journal of Sugar Beet Research, 2000, 37(4): 117-133 ( ) DOI: 10.5274/jsbr.37.4.117
- Gürel S., Gürel E., Kaya Z., Erdal M., Güler E. Effects of antimitotic agents on haploid plant production from unpollinated ovules of sugar beet (Beta vulgaris L.). Biotechnology & Biotechnological Equipment, 2003, 17(2): 97-101 ( ) DOI: 10.1080/13102818.2003.10817065
- Klimek-Chodacka M., Baranski R. Comparison of haploid and doubled haploid sugar beet clones in their ability to micropropagate and regenerate. Electron. J. Biotechn., 2013, 16(2): 1-12 ( ) DOI: 10.2225/vol16-issue2-fulltext-3
- Свирщевская А.М. Триплоиды, диплоиды и гаплоиды в селекции сахарной свеклы (Beta vulgaris L.). Весцi нацыянальнай Акадэмii навук Беларусi, 2008, 2: 48-55.
- Землянухин А.А., Землянухин Л.А. Метаболизм органических кислот растений. Воронеж, 1995.
- Левитес Е.В., Свирщевская А.М., Кирикович А.А., Милько Л.В. Удвоенные гаплоиды в изучении эпигенетической изменчивости сахарной свеклы. Информационный вестник ВОГИС, 2005, 9(4): 512-517.
- Beckmann J.S., Soller M. Restriction fragment length polymorphisms in genetic improvement: methodologies, mapping and costs. Theor. Appl. Genet., 1983, 67: 35-43 ( ) DOI: 10.1007/BF00303919
- Зайцев Г.Н. Методика биометрических расчетов. М., 1973.
- Подвигина О.А. Индуцирование гаплоидии из неоплодотворенных семяпочек сахарной свеклы в условиях in vitro. В сб.: Энциклопедия рода Вeta. Биология, генетика и селекция свеклы (науч. тр. Института цитологии и генетики СО РАН). Новосибирск, 2010: 455-465.
- Жужжалова Т.П., Фоменко Н.Р. Возрастные изменения сахарной свеклы в онтогенезе. Сахарная свекла, 2003, 7: 26-27.
- Жужжалова Т.П., Знаменская В.В., Подвигина О.А., Ярмолюк Г.И. Репродуктивная биология сахарной свеклы. Воронеж, 2007.
- Петров Д.Ф. Генетические основы апомиксиса. Новосибирск, 1979.
- Таратонов Н.А. Влияние экзогенных факторов на рост и развитие регенерантов в процессе формирования дигаплоидных форм сахарной свеклы. Канд. дис. Рамонь, 1999.
- Батыгина Т.Б., Круглова Н.Н., Горбунова В.Ю., Титова Г.Е., Сельдимирова О.А. От микроспоры -к сорту. М., 2010.
- Жужжалова Т.П., Подвигина О.А., Знаменская В.В. Пути воспроизведения нового организма сахарной свеклы в культуре in vitro. В сб.: Энциклопедия рода Вeta. Биология, генетика и селекция свеклы (науч. тр. Института цитологии и генетики СО РАН). Новосибирск, 2010: 403-419.
- Знаменская В.В. Микроклонирование in vitro как метод поддержания и размножения линий сахарной свеклы. В сб.: Энциклопедия рода Вeta. Биология, генетика и селекция свеклы (науч. тр. Института цитологии и генетики СО РАН). Новосибирск, 2010: 420-437.
- Васильченко Е.Н., Жужжалова Т.П. Изменчивость морфологических и биохимических признаков межвидовых гибридов свеклы. Сахарная свекла, 2011, 1: 18-21.
- Федулова Т.П. Теоретические и практические аспекты молекулярно-генетического маркирования в селекции сахарной свеклы (Beta vulgaris L.). Докт. дис. Рамонь, 2005.
- Kirikovich S.S., Svirshchevskaya A.M., Levites E.V. Variation at isozyme loci in seed offspring of sugar beet gynogenetic lines. Sugar Tech., 2003, 5(4): 289-292 ( ) DOI: 10.1007/BF02942487
- Levites E.V., Svirshchevskaya A.M., Kirikovich S.S., Mil’ko L.V. Variation at isozyme loci in cultured in vitro sugar beet regenerants of gynogenetic origin. Sugar Tech., 2005, 7(1): 71-75 ( ) DOI: 10.1007/BF02942421
- Nishizawa S., Kubo T., Mikami T. Variable number of tandem repeat loci in the mitochondrial genomes of beets. Curr. Genet., 2000, 37: 34-38 ( ) DOI: 10.1007/s002940050005
- Nishizawa S., Mikami T., Kubo T. Mitochondrial DNA phylogeny of cultivated and wild beets: relationships among cytoplasmic male-sterility-inducing and nonsterilizing cytoplasms. Genetics, 2007, 177: 1703-1712 ( ) DOI: 10.1534/genetics.107.076380
- Cheng D., Kitazaki K., Xu D., Mikami T., Kubo T. The distribution of normal and male-sterile cytoplasms in Chinese sugar-beet germplasm. Euphytica, 2009, 165: 345-354 ( ) DOI: 10.1007/s10681-008-9796-0
- Moritani M., Taguchi K., Kitazaki K., Matsuhira H., Katsuyama T., Mikami T., Kubo T. Identification of the predominant nonrestoring allele for Owen-type cytoplasmic male sterility in sugar beet (Beta vulgaris L.): development of molecular markers for the maintainer genotype. Mol. Breeding, 2013, 32: 91-100 ( ) DOI: 10.1007/s11032-013-9854-8
