Генетическая характеристика поволжских татар по данным об однородительских маркерах

Автор: Трофимова Наталья Вадимовна, Литвинов Сергей Сергеевич, Хусаинова Рита Игоревна, Ахметова Вита Леоновна, Ахатова Фарида Сериковна, Виллемс Рихард, Хуснутдинова Эльза Камилевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
Проведен анализ полиморфизма митохондриальной ДНК (мтДНК) и Y хромосомы в двух субпопуляциях татар, проживающих на территории Волго-Уральского региона России. Анализ полиморфизма мтДНК показал, что большинство обнаруженных гаплогрупп мтДНК характерны для народов Западной Евразии. Доля восточно-евразийского компонента у казанских и туймазинских татар составила 11,4% и 23% соответственно. В субпопуляциях татар были выявлены ранее не обнаруженные гаплогруппы B4, F. Показатель генного разнообразия по изученному набору гаплогрупп оказался выше у казанских татар по сравнению с туймазинскими татарами. Проведен анализ распределения частот гаплогрупп Y хромосомы, который показал, что мажорными гаплогруппами Y-хромосомы в обеих субпопуляциях являются R1a-Page07, N-M231 и I-M170. При этом у туймазинских татар, в отличие от казанских, была с относительно высокой частотой обнаружена гаплогруппа R1b-M269, что указывает, по всей видимости, на эффект основателя в этой субпопуляции.
Митохондриальная днк, y-хромосома, филогеография, поволжские татары
Короткий адрес: https://sciup.org/148202021
IDR: 148202021 | УДК: 575.174:599.9
Genetic characteristic of Volga Tatars based on uniparental markers data
The analysis of mitochondrial DNA in the two subpopulations of Tatars living in the Volga-Ural region of Russia was carried out. Analysis of mtDNA polymorphism showed that the majority of detected mtDNA are typical for the peoples of Western Eurasia. The share of East Eurasian component in the Kazan Tatars and Tuymazinsky was 10.7% and 22%, respectively. In Tatar subpopulations previously unidentified lines B4, F have been found. Genetic diversity index was higher in Kazan Tatars, compared with Tuymazinsky Tatars. Analysis of the Y-chromosome haplogroup frequency distribution showed that major haplogroups in two studied subpopulations are R1a-Page07, N-M231 and I-M170. At the same time Tuymazinsky Tatars, unlike Kazan Tatars, showed a relatively high frequency of R1b-M269 haplogroup, which indicates possible founder effect in this subpopulation.
Текст научной статьи Генетическая характеристика поволжских татар по данным об однородительских маркерах
Татары – одна из самых крупных народностей Российской Федерации. По численности в стране они уступают только русским. По данным переписи населения 2010 г., в общей сложности в России проживает около 5 млн 364 тыс татар. Наиболее многочисленная группа татар проживает в Республике Татарстан – 2 млн 43 тыс чел., в Башкортостане их насчитывается более 1 млн 132 тыс чел. Помимо Республики Татарстан, крупные по численности группы татарского населения присутствуют в Челябинской, Свердловской, Ульяновской, Самарской, Кировской областях, Пермском крае, Башкортостане, Удмуртии, Мордовии, Чувашии, Марий Эл [1]. Следует отметить, что татары расселены очень дисперсно, и более или менее многочисленные группы татарского населения имеются практически во всех субъектах Российской Федерации. Язык поволжских татар относится к кипчакским (кыпчакским) языкам тюркской группы алтайской семьи, к которой также причисляются башкиры [2].
По мнению Р.К. Кузеева [3], татары произошли от тюркоязычных племен, которые расселились в раннем средневековье на территории Средней Волги и Нижнего Прикамья, включив, по всей видимо-
сти, в свой состав более раннее финно-угорское и скифо-сарматское население этих территорий Восточной Европы.
В литературе имеются данные о генетическом разнообразии в субпопуляциях татар из Альметьевского, Елабужского [4-5], а также Азнакаевского и Буинского районов [6] Республики Татарстан, поэтому целью этой работы является получение представления о генетическом разнообразии поволжских татар Туймазинского района Республики Башкортостан, а также дисперсной выборки из различных районов Татарстана. При этом мы использовали комплексный подход, который включал в себя анализ полиморфизма как митохондриальной ДНК (мтДНК), так и нерекомбинирующей области Y-хромосомы, что позволяет оценить генетическую структуру популяций как по женской, так и по мужской линии.
Материалом для исследования служили образцы ДНК, выделенные из цельной крови неродственных индивидов, представляющих две субпопуляции татар: казанские татары из различных районов Татарстана (N=53) и татары, проживающие на территории Туймазинского района Республики Башкортостан (N=48). Несмотря на то, что в тексте обсуждаются туймазинские и казанские татары, данное разделение условно, поскольку указывает только на географическое различие, причем казанские татары, как было отмечено, представляют собой дисперсную выборку. Этническая принадлежность, происхождение и родственные отношения уточнялись путем анкетирования, учитывая данные до третьего поколения. Все обследованные дали информационное согласие на участие в исследовании.
Выделение тотальной ДНК проводилось стандартным фенол-хлороформным методом [7]. Проанализированы нуклеотидная последовательность гипервариабельного сегмента 1 (ГВС1) мтДНК длиной 377 п.н.(16024-16400), а также мутации в кодирующем регионе. Определение нуклеотидных последовательностей мтДНК осуществлялось на автоматическом секвенаторе Applied Biosystems (ABI) 3730XL DNA Analyzer. Последовательности были выровнены и проанализированы с помощью программы Sequencer 5.1. Определение гаплогрупп и гаплотипов мтДНК проводилось путем совместного анализа статуса диагностических сайтов рестрикции в кодирующем регионе мтДНК и мотивов ГВСI [8-10].
Показатель генного разнообразия рассчитывали по формуле [11]:
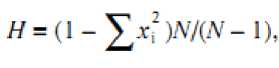
где x i - частота каждой гаплогруппы мтДНК в субпопуляции, N – объем выборки.
Анализ диаллельных маркеров нерекомбинирующей области Y-хромосомы проводился по следующим 14 локусам: M9, M89, YAP (M1), M20, M231, 92R7, M207, M306 (M173), Page07, M269, M170, 12f2, M175, M130. Гаплогруппы Y-хромосомы определялись согласно обновленной классификации консорциума по изучению Y-хромосомы (YCC) [12-14]. Исследование полиморфизма 14 диаллельных локусов Y-хромосомы производилось с использованием ПЦР и последующего ПДРФ анализа. Последний проводился с помощью эндонуклеаз рестрикции «Fermentas» (Литва) и «СибЭнзим» (Россия). Полиморфизм локусов YAP и 12f2 анализировали без проведения рестрикции.
В проведенном исследовании гаплогрупп мтДНК, выявленных у казанских татар и татар из Туймазинского района Республики Башкортостан, было показано, что в анализируемых субпопуляциях западно-евразийские линии преобладают над восточно-евразийскими. В таблице приведены частоты основных митохондриальных гаплогрупп, обнаруженных в ходе исследования.
У туймазинских татар наблюдалось более низкое значение индекса генетического разнообразия (Н) по проанализированному набору гаплогрупп мтДНК (0,92) по сравнению с казанскими татарами, у которых он составлял 0,94.
Западноевразийский компонент в изученных популяциях представлен гаплогруппами H, HV1, I1a, J1, K1, N, R2, T1a, T2, U1, U2, U4, U5, U5a, U5b, U8, V, V7, V13, W1, W3, W6, X2, Y1. Из них с максимальной частотой встречается гаплогруппа Н, относящаяся к числу самых распространенных гап-логрупп мтДНК в Евразии. Её частота составляет 40-50% всех гаплогрупп в большинстве стран
Таблица. Частоты гаплогрупп мтДНК у поволжских татар
|
Казанские татары |
Туймазинские татары |
|||
|
N |
Частота гаплогрупп, % |
N |
Частота гаплогрупп, % |
|
|
Западно-евразийский компонент |
||||
|
H |
11 |
20,8 |
12 |
25,2 |
|
HV1 |
0 |
0 |
1 |
2,1 |
|
I1a |
1 |
1,9 |
0 |
0 |
|
J1 |
7 |
13,2 |
4 |
8,3 |
|
K1 |
3 |
5,7 |
1 |
2,1 |
|
N |
3 |
5,7 |
0 |
0 |
|
R2 |
1 |
1,9 |
0 |
0 |
|
T1a |
2 |
3,8 |
4 |
8,3 |
|
T2 |
3 |
5,7 |
2 |
4,2 |
|
U1 |
1 |
1,9 |
0 |
0 |
|
U2 |
2 |
3,8 |
0 |
0 |
|
U4 |
1 |
1,9 |
4 |
8,3 |
|
U5 |
1 |
1,9 |
0 |
0 |
|
U5a |
2 |
3,8 |
2 |
4,2 |
|
U5b |
2 |
3,8 |
1 |
2,1 |
|
U8 |
1 |
1,9 |
2 |
4,2 |
|
V |
0 |
0 |
1 |
2,1 |
|
V7 |
1 |
1,9 |
1 |
2,1 |
|
V13 |
2 |
3,8 |
0 |
0 |
|
W1 |
0 |
0 |
1 |
2,1 |
|
W3 |
1 |
1,9 |
0 |
0 |
|
W6 |
0 |
0 |
1 |
2,1 |
|
X2 |
1 |
1,9 |
0 |
0 |
|
Y1 |
1 |
1,9 |
0 |
0 |
|
Всего |
47 |
89,1 |
37 |
77,4 |
|
Восточно-евразийский компонент |
||||
|
A |
1 |
1,9 |
1 |
2,1 |
|
B4 |
1 |
1,9 |
1 |
2,1 |
|
B5 |
0 |
0 |
1 |
2,1 |
|
C4 |
0 |
0 |
1 |
2,1 |
|
D4 |
3 |
5,7 |
4 |
8,3 |
|
F1a |
0 |
0 |
1 |
2,1 |
|
G2a |
0 |
0 |
2 |
4,2 |
|
M3a |
1 |
1,9 |
0 |
0 |
|
Всего |
6 |
11,4 |
11 |
23 |
Европы, и около 20-30% гаплогрупп Ближнего Востока и Кавказа [15-18].
У казанских татар ее частота составляет 20,8 %, а у туймазинских 25,2%. Данная картина в принципе соответствует распределению гаплогруппы H на территории Евразии.
В изученных нами выборках татар была обнаружена гаплогруппа V с частотой 5,7% у казанских татар и 4,2% у туймазинских. Как было показано ранее, данная гаплогруппа распространена в Западной и Восточной Европе с пиками в стране Басков [15] и в популяции саамов [19]. Возраст ее коалесценции варьирует от 16,3±4,8 тыс лет назад в Западной Европе до 8,5 ± 2,3 тыс лет назад на Востоке Европы [15, 20]. Кроме того, у казанских татар была обнаружена редко встречающаяся субгапло-группа V13, характеризующаяся заменой G12795A относительно rCRS [21]. Данная субгаплогруппа ранее была обнаружена в популяции татар [22]. Кроме того, она встречается в финно-угорских и тюркских популяциях Волго-Уральского региона [наши неопубликованные данные].
Восточно-евразийский компонент в изученных популяциях татар представлен гаплогруппами A, B4, B5, C4, D4, F1a, G2a, M3a и составляет 11,4% митохондриального пула у казанских татар и 23% у туймазинских. Отдельный интерес представляют гаплогруппы В4 и В5. Гаплогруппа B4 присутствует в генофондах практически всех этнических групп Северной и Восточной Азии, за исключением восточных эвенков, чукчей, коряков и эвенов. Максимальная частота гаплогруппы B4 зарегистрирована у корейцев (20,4%) и монголов (14,9%), а в Северной Азии ее частота изменяется в диапазоне от 3,1% у якутов до 11,3% у теленгитов [23]. Также было показано, что эти гаплогруппы встречаются у тувинцев и тофалар с частотой 5,3% и 4,3% соответственно [24]. Интересно, что гаплогруппы В4 и С4 были обнаружены при анализе древней ДНК из двух археологических памятников в Западной Сибири: Сайгатинский 6 (с VIII по XI вв.) и Зеленый Яр (XVIII в.) [25]. Ранее гаплогруппа B в ВолгоУральском регионе была обнаружена только у башкир [4]. Результаты нашего исследования показывают наличие этой гаплогруппы и в субпопуляции поволжских татар, причем гаплотип ГВС1 у проанализированных нами образцов для субгапло-группы B4b1 идентичен и представлен мотивом 16136, 16183C, 16189, 16217, 16256, 16519, что может свидетельствовать в пользу недавней дивергенции субпопуляций поволжских татар. С другой стороны, малая частота данной гаплогруппы в изученных популяциях не позволяет однозначно трактовать полученные результаты.
Гаплогруппы B5, C4, F1a, G2a были обнаружены у туймазинских татар и отсутствовали у казанских. Гаплогруппа G2 c высокой частотой встречается в популяциях бурят (11%), монголов (8,5%) [24], а в целом гаплогруппа G присутствует также в Волго-Уральском регионе – у коми (2,4%), татар (1,6%), чувашей (0,7%) и башкир (4,8%) [4].
В результате анализа Y-хромосомы нами было обнаружено различие в генетической структуре изученных субпопуляций. Несмотря на то, что пик частот в обеих субпопуляциях приходится на мажорные гаплогруппы R1a-Page07, N-M231 и I-M170, у казанских татар не было обнаружено гап-логруппы R1b-M269, в то время как у туймазин-ских ее доля составляет 15,7%. Даже учитывая, что в настоящий момент микросателлитное разнообразие внутри данной гаплогруппы у туймазинских татар не было проанализировано, обнаруженное различие весьма интересно. Однако можно предположить, что в данной субпопуляции имел место эффект основателя.
Таким образом, изученные субпопуляции татар в целом демонстрируют распределение гаплогрупп мтДНК, характерное для популяций Евразии с преобладающей в митохондриальном пуле гаплогруп-пой H. При этом восточно-евразийский компонент имеет большую встречаемость у туймазинских татар, хотя в целом его доля в изученных субпопуляциях существенно ниже по сравнению с западноевразийским. Мажорными гаплогруппами Y-хромосомы в обеих субпопуляциях являются R1a-Page07, N-M231 и I-M170. При этом у туймазин-ских татар, в отличие от казанских, была с относительно высокой частотой была обнаружена гапло-группа R1b-M269, что указывает, по всей видимости, на эффект основателя в этой субпопуляции.
-
1. Всероссийская перепись населения 2010 года. URL: http://www.perepis-2010.ru
-
2. Белых С.К. История народов Волго-Уральского региона: Учебное пособие. Ижевск, 2006. 54 с.
-
3. Кузеев Р.Г. Народы Среднего Поволжья и Южного Урала: этногенетический взгляд на историю. М.: Наука, 1992. 345 с.
-
4. Бермишева М.А., Тамбетс К., Виллемс Р., Хуснутдинова Э.К. Разнообразие гаплогрупп митохондриальной ДНК в этнических группах Волго-Уральского региона // Мол. биология. 2002. Т. 36. С. 990-1001.
-
5. Орехов В.А . Характеристика митотипов представителей трех этнических групп европейской части России: Авто-реф. дис. … канд. биол. наук. М.: ИОГен РАН, 2002. 22 с.
-
6. Денисова Г.А., Малярчук Б.А., Деренко М.В., Кравцова О.А. Популяционная структура поволжских татар по данным о разнообразии митохондриальной ДНК // Генетика. 2011. Т. 47. № 3. С. 387-393.
-
7. Mathew C.C . The isolation of high molecular weight eucari-otic DNA // Methods in molecular biology/ Ed. Walker J.M. New York, 1984. P. 31-34.
-
8. Macaulay V., Richards M., Hickey E. et al . The emerging tree of West Eurasian mtDNAs: a sinthesis of control region sequenses and RFLPs // Am. J. Genet. 1999. V. 64. P. 232249.
-
9. Richards M., Macaulay V., Hickey E. et al. Tracing European founder lineages in the Near eastern mtDNA pool // Am. J. Hum. Genet. 2000. V. 67. P. 1251-1276.
-
10. Oven M. van, Kayser M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation // Hum. Mutat. 2009. V. 30. N. 2. P. 386-394.
-
11. Nei M. Molecular Evolutionary Genetics. New York: Columbia Univ. Press, 1988.
-
12. Y Chromosome Consortium. A nomenclature system for the tree of human Y-chromosomal binary haplogroups // Genome Res. 2002. V. 12. P. 339-348.
-
13. Karafet T.M., Mendez F.L., Meilerman M.B., Underhill P.A., Zegura S.L., Hammer M.F . New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree // Genome Res. 2008. V. 5. P. 830-838.
-
14. Myres N.M., Rootsi S., Lin A.A. et al. A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe // Eur. J. Hum. Genet. 2011. V. 19. P. 95-101.
-
15. Álvarez-Iglesias V., Mosquera-Miguel A., Cerezo M. et al . A. New Population and Phylogenetic Features of the Internal Variation within Mitochondrial DNA Macro-Haplogroup R0 // PLoS ONE. 2009. 4(4): e5112.
-
16. Achilli A., Rengo C., Magri C. et al. The molecular dissection of mtDNA haplogroup H confirms that the Franco-Cantabrian glacial refuge was a major source for the European gene pool // Am. J. Hum. Genet. 2004. V. 75. P. 910918.
-
17. Loogvali E.L., Roostalu U., Malyarchuk B.A. et al . Disuniting uniformity: a pied cladistic canvas of mtDNA haplo-group H in Eurasia // Mol. Biol. Evol. 2004. V. 21. P. 20122021.
-
18. Roostalu U., Kutuev I., Loogvali E.L. et al. Origin and expansion of haplogroup H, the dominant human mitochondrial DNA lineage in West Eurasia: the Near Eastern and Caucasian perspective // Mol. Biol. Evol. 2007. V. 24. P. 436448
-
19. Tambets K., Rootsi S., Kivisild T. et al. The Western and Eastern roots of the Saami – the story of genetic “outliers” told by mitochondrial DNA and Y-chromosomes // Am. J. Hum. Genet. 2004. V. 74. P. 661-682.
-
20. Torroni A., Bandelt H.-J., Macaulay V., Richards M., Cru-ciani F. et al . A signal, from human mtDNA, of postglacial recolonization in Europe // Am. J. Hum. Genet. 2001. V. 69.
-
21. Andrews R.M., Kubacka I., Chinnery P.F. et al . Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA // Nature Genet. 1999. V. 23. P. 147.
-
22. Malyarchuk B., Derenko M., Denisova G., Kravtsova O . Mi-togenomic diversity in Tatars from the Volga-Ural region of Russia // Mol. Biol. and Evolution. 2010. V. 27. P. 22202226.
-
23. Деренко М.В., Малярчук Б.А . Молекулярная филогеография населения Северной Евразии по данным об изменчивости митохондриальной ДНК / Oтв. ред. И.А. За-харов-Гезехус. Магадан: СВНЦ ДВО РАН, 2010. 120 с.
-
24. Starikovskaya E.B., Sukernik R.I., Derbeneva O.A. et al . Mitochondrial DNA diversity in indigenous populations of the southern extent of Siberia, and the origins of Native American haplogroups // Ann. Hum. Genet. 2005. V. 69. P. 67-89.
-
25. Sato A., Razhev D., Amano T., Masuda R . Genetic features of ancient West Siberian people of the Middle Ages, revealed by mitochondrial DNA haplogroup analysis // J. Human Genetics. 2011. V. 56. P. 602-608.
P. 844-852.
GENETIC CHARACTERISTIC
ON UNIPARENTAL
OF VOLGA TATARS BASED MARKERS DATA
Список литературы Генетическая характеристика поволжских татар по данным об однородительских маркерах
- Всероссийская перепись населения 2010 года. URL: http://www.perepis-2010.ru
- Белых С.К. История народов Волго-Уральского региона: Учебное пособие. Ижевск, 2006. 54 с.
- Кузеев Р.Г. Народы Среднего Поволжья и Южного Урала: этногенетический взгляд на историю. М.: Наука, 1992. 345 с.
- Бермишева М.А., Тамбетс К., Виллемс Р., Хуснутдинова Э.К. Разнообразие гаплогрупп митохондриальной ДНК в этнических группах Волго-Уральского региона//Мол. биология. 2002. Т. 36. С. 990-1001.
- Орехов В.А. Характеристика митотипов представителей трех этнических групп европейской части России: Автореф. дис. … канд. биол. наук. М.: ИОГен РАН, 2002. 22 с.
- Денисова Г.А., Малярчук Б.А., Деренко М.В., Кравцова О.А. Популяционная структура поволжских татар по данным о разнообразии митохондриальной ДНК//Генетика. 2011. Т. 47. № 3. С. 387-393.
- Mathew C.C. The isolation of high molecular weight eucariotic DNA//Methods in molecular biology/Ed. Walker J.M. New York, 1984. P. 31-34.
- Macaulay V., Richards M., Hickey E. et al. The emerging tree of West Eurasian mtDNAs: a sinthesis of control region sequenses and RFLPs//Am. J. Genet. 1999. V. 64. P. 232-249.
- Richards M., Macaulay V., Hickey E. et al. Tracing Euro pean founder lineages in the Near eastern mtDNA pool//Am. J. Hum. Genet. 2000. V. 67. P. 1251-1276.
- Oven M. van, Kayser M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation//Hum. Mutat. 2009. V. 30. N. 2. P. 386-394.
- Nei M. Molecular Evolutionary Genetics. New York: Columbia Univ. Press, 1988.
- Y Chromosome Consortium. A nomenclature system for the tree of human Y-chromosomal binary haplogroups//Genome Res. 2002. V. 12. P. 339-348.
- Karafet T.M., Mendez F.L., Meilerman M.B., Underhill P.A., Zegura S.L., Hammer M.F. New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree//Genome Res. 2008. V. 5. P. 830-838.
- Myres N.M., Rootsi S., Lin A.A. et al. A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe//Eur. J. Hum. Genet. 2011. V. 19. P. 95-101.
- Álvarez-Iglesias V., Mosquera-Miguel A., Cerezo M. et al. A. New Population and Phylogenetic Features of the Internal Variation within Mitochondrial DNA Macro-Haplogroup R0//PLoS ONE. 2009. 4(4): e5112.
- Achilli A., Rengo C., Magri C. et al. The molecular dissection of mtDNA haplogroup H confirms that the Franco-Cantabrian glacial refuge was a major source for the European gene pool//Am. J. Hum. Genet. 2004. V. 75. P. 910-918.
- Loogvali E.L., Roostalu U., Malyarchuk B.A. et al. Disuniting uniformity: a pied cladistic canvas of mtDNA haplogroup H in Eurasia//Mol. Biol. Evol. 2004. V. 21. P. 2012-2021.
- Roostalu U., Kutuev I., Loogvali E.L. et al. Origin and expansion of haplogroup H, the dominant human mitochondrial DNA lineage in West Eurasia: the Near Eastern and Caucasian perspective//Mol. Biol. Evol. 2007. V. 24. P. 436-448
- Tambets K., Rootsi S., Kivisild T. et al. The Western and Eastern roots of the Saami -the story of genetic “outliers” told by mitochondrial DNA and Y-chromosomes//Am. J. Hum. Genet. 2004. V. 74. P. 661-682.
- Torroni A., Bandelt H.-J., Macaulay V., Richards M., Cruciani F. et al. A signal, from human mtDNA, of postglacial recolonization in Europe//Am. J. Hum. Genet. 2001. V. 69. P. 844-852.
- Andrews R.M., Kubacka I., Chinnery P.F. et al. Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA//Nature Genet. 1999. V. 23. P. 147.
- Malyarchuk B., Derenko M., Denisova G., Kravtsova O. Mitogenomic diversity in Tatars from the Volga-Ural region of Russia//Mol. Biol. and Evolution. 2010. V. 27. P. 2220-2226.
- Деренко М.В., Малярчук Б.А. Молекулярная филогеография населения Северной Евразии по данным об изменчивости митохондриальной ДНК/Oтв. ред. И.А. Захаров-Гезехус. Магадан: СВНЦ ДВО РАН, 2010. 120 с.
- Starikovskaya E.B., Sukernik R.I., Derbeneva O.A. et al. Mitochondrial DNA diversity in indigenous populations of the southern extent of Siberia, and the origins of Native American haplogroups//Ann. Hum. Genet. 2005. V. 69. P. 67-89.
- Sato A., Razhev D., Amano T., Masuda R. Genetic features of ancient West Siberian people of the Middle Ages, revealed by mitochondrial DNA haplogroup analysis//J. Human Genetics. 2011. V. 56. P. 602-608.
