Генетическая структура популяции соболя ( Martes zibellina ) Буреинского нагорья по сборам одного охотничьего сезона
 Буреинского нагорья по сборам одного охотничьего сезона")
Автор: Фрисман Л.В., Шлюфман К.В., Брыкова А.Л.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Биология. Математическое моделирование
Статья в выпуске: 4 т.22, 2019 года.
Бесплатный доступ
Основываясь на анализе трех микросателлитных локусов, проведено исследование аллельного разнообразия и уровня генетической дифференциации для 4 географических субпопуляций соболя Буреинского нагорья и одной субпопуляции хребта Сихотэ-Алинь. Обнаружено 23 аллеля. Наибольшее аллельное разнообразие было обнаружено в субпопуляции хребра Сихотэ-Алинь (22 аллеля). 18 из 23 обнаруженных аллелей являются общими для соболей Буреинского нагорья и Сихотэ-Алиня. 5 из 23 обнаруженных аллелей оказались специфическими, т.е. представлены в субпопуляциях какой-либо одной из рассматриваемых горных систем. Однако частота встречаемости специфических аллелей в субпопуляциях достаточно низкая. Полученные при парном сравнении рассматриваемых географических субпопуляций показатели F(ST) отражают уровень слабой генетической дифференциации по шкале Райта (F(ST)
Соболь (martes zibellina), микросателлиты, аллельное разнообразие, генетическая дифференциация, буреинское нагорье, хребет сихотэ-алинь
Короткий адрес: https://sciup.org/143169082
IDR: 143169082 | УДК: 575.174:599.742.4(571.6) | DOI: 10.31433/2618-9593-2019-22-4-46-55
Genetic structure of the Bureinsky highlands sable (Martes zibellina) population on base of one hunting season collections
Based on microsatellite analysis of 3 loci, allelic diversity and the level of genetic differentiation were investigated for four geographic sable subpopulations of the Bureinsky highlands and one sable subpopulation of the Sikhote-Alin ridge. Totally, it was found 23 alleles. The researchers found the greatest allelic diversity in one subpopulation of the Sikhote-Alin ridge (22 alleles). Eighteen alleles of 23 detected ones were common to the Bureinsky highlands and Sikhote-Alin sable communities. Five of 23 detected alleles were specific ones. It was found four specific alleles in the Sikhote-Alin subpopulation, and only one specific allele - in the Bureinsky highlands. However, the occurrence of specific alleles in subpopulations is rather low. The pairwise F (ST) of geographical subpopulations reflected the minimum level of their intraspecific genetic differentiation according to the Wright scale (F(ST)
Текст научной статьи Генетическая структура популяции соболя ( Martes zibellina ) Буреинского нагорья по сборам одного охотничьего сезона
Соболь (Martes zibellina Linnaeus, 1758) – промысловый пушной вид млекопитающих, заселяющий хвойные и смешанные леса северной Евразии. Его ареал протяжен по территории России от Урала до Тихого океана и лишь частично заходит в сопредельные страны: Монголию, Китай, Корею и Японию. Высокая ценность пушнины и многовековой стабильный спрос привели в начале ХХ века к практически полному исчезновению многих популяций соболя на российской части ареала. Централизованной мерой сохранения вида явились запрещение промысла и, спустя 5 лет, интродукция животных на угнетенные территории. Из Прибайкалья происходило расселение по европейской части ареала, Уралу, Западной Сибири и Якутии [9]. Из Верхнебуреинского племенного соболиного рассадника расселение соболя в основном осуществлялось по дальневосточной части ареала. Зверьки отсюда были перемещены в Амурскую и Магаданскую области, Якутию, Хабаровский и Приморский края, а также в Читин- скую область [3]. Проведенная работа благотворно повлияла на численность соболей на территории России. Возникшая в результате рукотворного толчка, вновь сформировавшаяся структура популяций отдельных регионов и вида в целом, несомненно, должна была отличаться от исходной, привлекая внимание биологов к ее описанию и исследованию. Появился обширный спектр морфологических работ и возникла потребность получения генетических данных о внутривидовом разнообразии. Морфологические исследования не привели к единому представлению о современном подвидовом подразделении соболя России. На территориях интродукции были описаны популяции аборигенных форм, популяции интродуцен-тов и гибридные популяции. У акклиматизантов был обнаружен интенсивный процесс смещения морфологических характеристик в сторону аборигенных форм. Последнее могло являться результатом адаптации к новой окружающей среде и (или) отражать процесс гибридизации между интродуцированными и автохтонными формами [12].
Исследование генетической изменчивости и структуры популяций было начато на основе анализа маркеров митохондриальной ДНК на материале северо-восточной части ареала соболя [2, 11] и затем расширено на пределы всего ареала [13, 20].
В последнее десятилетие для исследования внутривидовой генетической изменчивости и структуры природных популяций соболя активно и успешно применяется анализ полиморфизма микросателлитов – кодоминантно наследуемых и адаптивно нейтральных ядерных генетических маркеров. Так, анализ аллельного разнообразия пяти микросателлитных локусов в современной популяции бассейна реки Вах, берущей свое начало от интродуцентов из Прибайкалья, показал ее происхождение в результате перемешивания генофондов интродуцентов и представителей приобских автохтонов [10]. Исследование аллельного разнообразия восьми микросателлитных локусов в семи географических популяциях соболя из пространственно удаленных регионов (Урал, Сибирь, хребет Сихотэ-Алинь и полуостров Камчатка) обнаружило большие генетические различия между популяциями краевых регионов в сравнении с генетически близкими сибирскими. Наибольшее аллельное разнообразие было выявлено в популяции восточных отрогов хребта Сихотэ-Алинь. Последний результат позволил С.Н. Каштанову с соавторами [6] присоединиться к гипотезе В.Г. Абрамова о существовании на Сихотэ-Алине в последний ледниковый период соболиного рефугиума [1], откуда шло последующее продвижение этого вида по территории северной Евразии. О справедливости этой гипотезы говорили и данные молекулярно-генетического исследования субъединицы 2 митохондриальной NADH-дегидрогеназы [21]. При этом J.J. Sаtо с соавторами обнаружили самое высокое разнообразие в группе соболей южного Сихотэ-Алиня. Последующее исследование [20] было проведено с включением большого объема материала из левобережных горных систем Среднего Приамурья, в том числе Буреинского нагорья. По рассматриваемому признаку обитатели данного региона, совместно с приморскими, вошли в число наиболее полиморфных по ареалу. В силу отсутствия в последний ледниковый период крупных покровных оледенений на территории юга Дальнего Востока, возможно, Буреинское нагорье – это территория наиболее раннего расселения в голоцене либо постоянного обитания соболя в последний ледниковый период.
В начале ХХ века, во времена антропогенно организованной депрессии численности соболя, на Сихотэ-Алине этот зверек остался лишь в высокогорных недоступных охотникам районах и полностью отсутствовал в южной части Буреинского нагорья. В период искусственного расселения соболя на хребет Сихотэ-Алинь были интродуцированы животные из Иркутской области [4] и из светлохвойных лесов верхнего течения реки Бурея (Верхнебуреинского племенного соболиного рассадника). Из последнего источника были расселены зверьки и по Буреинскому нагорью. Везде, где был произведен выпуск соболей, они хорошо приживались и начали активно размножаться [3].
В настоящее время обитатели Среднего Приамурья могут быть потомками как аборигенной, так и интродуцированных форм. Несомненный интерес представляет сравнение современной популяционной структуры у обитателей Буреинско-го нагорья и хребта Сихотэ-Алинь, разделенных в пределах Среднего Приамурья географически и экологически важным географическим барьером – рекой Амур с болотистой Среднеамурской низменностью. Морфологические исследования показывают некоторые отличия лево- и правобережных амурских соболиных групп. Приамурский и Приморский регионы заселяют соболя, характеризующиеся мелкими размерами. При этом самыми мелкими среди них являются обитатели Сихотэ-Алиня [9].
В предварительном исследовании аллельного состава микросателлитного локуса Ма-3 в сборах соболей Среднего Приамурья [15] наряду с широко распространенными аллелями 131 и 133 были обнаружены аллели, встречающиеся с небольшой частотой, но при этом маркирующие либо географические выборки Буреинско-го хребта (аллель 135), либо выборку с Сихотэ-Алиня (аллель 129). В настоящей работе продолжено исследование популяционной структуры соболя Среднего Приамурья с использованием накопленного биологического материала сборов одного зимнего сезона 2012–2013 гг. Кроме ранее рассматриваемых локусов Ма-2 и Ма-3, в список исследования включен Мег-41, который, судя по данным Каштанова с соавторами [6], перспективен для исследования внутривидовой географической изменчивости соболя. Задача настоящего исследования – оценка аллельного разнообразия и уровня дифференциации по трем микросател-литным локусам для соболя Среднего Приамурья, обитающего на территории Буреинского нагорья в сравнении с материалом с хребта Сихотэ-Алинь.
Микросателлитные локусы, как правило, характеризуются большим числом аллелей, что предполагает использование локальных выборок достаточно большого объема, а при недостатке материала объединение сборов отдельных географических районов. Последний подход был использован в настоящей работе для некоторых географических районов.
Материал и методы
Использован биологический материал от 117 соболей охотничьих сборов зимнего сезона 2012–2013 гг. из 5 географических районов. Четыре района («Маган», «Сутырь», «Тырма» и «Ка-менушка») локализованы на Буреинском нагорье, расположенном по левой стороне от реки Амур. Самый северный район «Маган» находится на территории бывшего Верхнебуреинского соболиного рассадника. Для районов «Маган» и «Каменушка» проведено суммирование географически близких точек сбора. Выборка «Манома» локализована по правую сторону от реки Амур – на западном склоне хребта Сихотэ-Алинь. В табл. 1 приведены точ- ки сбора материала и их группировка в сборные выборки, характеризующие соболей соответствующих географических районов, которые мы далее называем субпопуляциями.
На рис. 1 представлена карта-схема локализации точек сбора материала.
ДНК выделяли из мышечной ткани с использованием стандартного метода солевой экстракции [16]. Количество выделенной ДНК определяли при помощи спектрофотометра Nano-drop 2000 либо при электрофорезе в 1,5% агарозном геле в присутствии ДНК фага лямбда.
Использовали амплификационную смесь следующего состава из расчета 10 µl. на 1 образец (30-50- ng ДНК): 10 ПЦР буфер KCl+, 1,5 мМ MgCl2, 0,2 мМ dNTP, 0,5 мМ прямой и обратный праймеры, 0,5U Taq ДНК полимераза. Амплификацию проводили в следующем режиме: 8 циклов, включающих 1 минуту денатурации ДНК при температуре 94 °С, отжиг праймеров 30 секунд –
Таблица 1
Географическое происхождение образцов Martes zibellina используемых в данном исследовании
Table 1
Geographic origin of the Martes zibellina samples used in this study
Mapper, версия 4.1 (Applied Biosystems, США). Оценка результатов изучения полиморфизма микросателлитов была проведена на основе анализа генотипических и аллельных частот. Оценки частот аллелей в суммарных географических выборках вычислялись по формуле [8]:
( r ^
Pi = nu + 0.5 'E nU / N
ˆ
I j = 1
7 ,
где p ˆ i – искомая частота i -го аллеля, r – количество аллелей в полиморфном локусе, ni,j – число особей с генотипом, соответствующим комбинации i , j – аллелей в локусе, N – объем выборки;
и их стандартная ошибка:
SE ( pi ) = VVar l p ) , где Var ( pi ) - выборочная биномиальная варианса ( 2 ) в состоянии равновесия популяции по Харди-Вайнбергу:
Var ( pi ) = pi " ( 1 - pi ) /2 N •
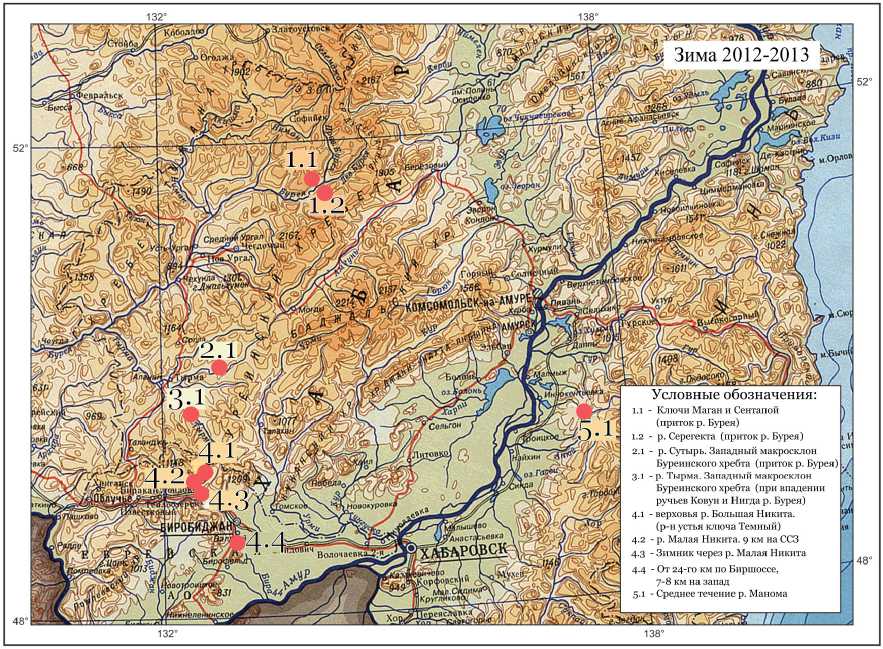
Рис. 1. Карта мест сбора материала, используемого в данном исследовании. Номера на карте соответствуют таковым в табл. 1
Fig. 1. Sampling locations map used in the study.
Numbers on the map correspond to those in Table 1
Если в выборке состояние равновесия нарушено, то:
Var ( pi ) =( pi + ni, i / N - 2 pI ) /2 N .
Определение частоты встречаемости «нуль»-аллеля и ее влияния на частоты остальных микросателлитных аллелей выполнено в программе Micro-checker 2.2.3 [22]. Оценки неравновесного сцепления генов, наблюдаемой и ожидаемой гетерозиготности, соответствия распределению Харди-Вайнберга и F-статистик проводили в программе Arlequin [17].
Расчет оценки потока генов между локальными субпопуляциями ( Nm ) проведен по формуле:
4 N m = —7 -1 , F ( ST )
Таблица 2
Генетические характеристики исследованных микросателлитных локусов
Table 2
Genetic characteristics of the investigated microsatellite loci
|
№ п/п |
Районы и объединенные территории |
Размер Выборки |
NA |
Ho |
HЕ |
HWE |
|
2.1 Локус Ma-2 |
||||||
|
1 |
Маган |
14 |
6 |
0.769 |
0.834 |
0.349 |
|
2 |
Сутырь |
24 |
6 |
0.625 |
0.791 |
0.223 |
|
3 |
Тырма |
28 |
7 |
0.778 |
0.774 |
0.985 |
|
4 |
Каменушка |
21 |
6 |
0.524 |
0.771 |
0.060 |
|
5 |
Манома |
30 |
7 |
0.867 |
0.787 |
0.074 |
|
6 |
Буреинское нагорье |
87 |
7 |
0.671 |
0.785 |
0.110 |
|
7 |
Среднее Приамурье |
117 |
7 |
0.722 |
0.786 |
0.227 |
|
2.2. Локус Ma-3 |
||||||
|
1 |
Маган |
14 |
3 |
0.461 |
0.396 |
1,00 |
|
2 |
Сутырь |
24 |
3 |
0.458 |
0.541 |
0,045 |
|
3 |
Тырма |
28 |
3 |
0.458 |
0.552 |
0.283 |
|
4 |
Каменушка |
21 |
3 |
0.476 |
0.479 |
0.080 |
|
5 |
Манома |
30 |
3 |
0.866 |
0.787 |
0.074 |
|
6 |
Буреинское Нагорье |
87 |
3 |
0.482 |
0.507 |
0.031 |
|
7 |
Среднее Приамурье |
117 |
4 |
0.469 |
0.478 |
0.027 |
|
2.3. Локус Mer-41 |
||||||
|
1 |
Маган |
14 |
9 |
0.615 |
0.812 |
0.100 |
|
2 |
Сутырь |
24 |
6 |
0.792 |
0.859 |
0.407 |
|
3 |
Тырма |
28 |
8 |
0.667 |
0.813 |
0.063 |
|
4 |
Каменушка |
21 |
7 |
0.843 |
0.809 |
0.737 |
|
5 |
Манома |
30 |
12 |
0,700 |
0,897 |
0.027 |
|
6 |
Буреинское Нагорье |
87 |
9 |
0.729 |
0.836 |
0.141 |
|
7 |
Среднее Приамурье |
117 |
12 |
0.721 |
0.861 |
0.012 |
Примечание: NA – число аллелей по локусу, HE – ожидаемая гетерозиготность, HO – наблюдаемая гетерозиготность. Гипотеза о соответствии распределению Харди-Вайнберга HWE принималась на уровне значимости 0,05
где N – эффективный размер популяции, а m – коэффициент миграции [23].
Результаты
В табл. 2 приведены сведения о количестве аллелей, наблюдаемой и ожидаемой гетерозиготности, а также результаты точного теста на соответствие распределению Харди-Вайнберга отдельно для локальных субпопуляций, суммарно для субпопуляций Буреинского хребта и суммарно для всех исследованных субпопуляций (Среднее Приамурье).
В табл. 3 показано общее число аллелей в каждой из региональных выборок, среднее по трем локусам число аллелей, ожидаемая и наблюдаемая гетерозиготности, а также значения индивидуального индекса фиксации Райта F(IS).
Все рассматриваемые локусы полиморфные. Наименьшим аллельным разнообразием обладает локус Ма-3, для которого представлено по три аллеля в каждой субпопуляции, тогда как общее число аллелей этого локуса, обнаруженных в приамурских субпопуляциях в сборах зимнего сезона 2012–2013, равно четырем. Локус Ма-2 занимает промежуточное положение по уровню полиморф-ности, с шестью или семью аллелями в каждой из субпопуляций при общем числе аллелей равным семи. Для Mer-41, обладающего самым высоким уровнем полиморфности среди рассматриваемых нами локусов, в субпопуляциях представлено от шести до двенадцати аллелей при общем числе аллелей равным двенадцати. Рассматриваемые локусы не показали неравновесного сцепления (при уровне значимости 0,05), что указывает на их расположение в различных хромосомах. Как видно из табл. 2 (столбцы 5, 6), соотношение величин наблюдаемой и ожидаемой гетерозиготности в большинстве рассматриваемых локальных выборок указывают на некоторый недостаток гетерозигот. Некоторый недостаток гетерозигот обнаружен как при суммировании локальных географических выборок Буреинского нагорья, так и при суммировании всех рассматриваемых выборок Среднего Приамурья. На некоторый недостаток гетерозигот (3,6–14,7% особей в локальных выборках и 9,7–10% в суммарных выборках) указывают и полученные для средних по трем локусам значения индивидуального индекса фиксации Райта F(IS) (столбец 8 табл. 3). Результаты точного теста показали отклонения от распределения Харди-Вайнберга (на уровне значимости 0,05) для локуса Ма-3 в субпопуляции «Сутырь» и локуса Mer-41 в субпопуляции «Манома» (столбец 7 табл. 2). Для
Таблица 3
Суммарные генетические характеристики исследуемых локусов
Table 3
Total genetic characteristics of the investigated loci
|
№ п/п |
Районы и объединенные территории |
Количество животных |
NО |
Среднее по 3-м локусам |
|||
|
NA |
Ho |
He |
F(IS) |
||||
|
1 |
«Маган» |
14 |
17 |
6 (1.732) |
0.615 (0.078) |
0.681 (0.075) |
0.036 |
|
2 |
«Сутырь» |
24 |
16 |
5.67 (1.202) |
0.625 (0.057) |
0.730 (0.052) |
0.147 |
|
3 |
«Тырма» |
28 |
18 |
6 (1.453) |
0.654 (0.053) |
0.713 (0.050) |
0.084 |
|
4 |
«Каменушка» |
21 |
16 |
5.33 (1.202) |
0.603 (0.062) |
0.698 (0.058) |
0.139 |
|
5 |
«Манома» |
30 |
22 |
7.33 (2.646) |
0.667 (0.050) |
0.691 (0.049) |
0.100 |
|
6 |
Буреинское нагорье |
87 |
19 |
6.33 (1.764) |
0.627 (0.030) |
0.710 (0.028) |
0.097 |
|
7 |
Среднее Приамурье |
117 |
23 |
7.67 (2.333) |
0.638 (0.026) |
0.709 (0.024) |
0.101 |
Примечание: Nо – общее число аллелей в выборке, NA – число аллелей по локусу, HE – ожидаемая гетерозиготность, HO – наблюдаемая гетерозиготность. Стандартные ошибки величин NA, HE и HO представлены в скобках. F(IS) ─ значения индивидуального индекса фиксации Райта суммарной выборки субпопуляций соболей Бу-реинского нагорья по локусу Ма-3 статистически отклонена гипотеза о соответствии панмиктичной популяции. В суммарной выборке соболей Среднего Приамурья гипотеза о соответствии панмиктич-ной популяции отклонена по двум локусам: Ма-3 и Mer-41. Полученный результат может отражать некоторую внутреннюю гетерогенность как указанных локальных субпопуляций, так и соболей Буреинского нагорья и Среднего Приамурья в целом. Однако возможны и другие причины нарушения панмиксии, среди них – наличие нуль-аллелей, т.е. отсутствие амплификации при ПЦР из-за мутаций во фланкирующих микросателлит-последо-вательностях ДНК, с которыми происходит гибридизация праймеров. Микросателлитный аллель, сцепленный с такой мутацией, не амплифицирует-ся у гомозиготной по нуль-аллелю особи, а у гетерозиготных по нему экземпляров амплифициру-ется лишь один микросателлит; и в таком случае гомозиготность является ложной [7]. Она влечет за собой смещение популяционно-генетических оценок, в том числе определяет избыток гомозигот. Среди исследованных 117 экземпляров ни по одному из рассматриваемых локусов мы не обнаружили животных гомозиготных по нуль-аллелю, т.е. с отсутствием ПЦР-продукта. Оценка частоты встречаемости нуль-аллелей у гетерозиготных особей предполагает знание доли наблюдаемого недостатка гетерозигот, обусловленного присутствием нуль-аллеля. Однако, даже если рассматривать весь наблюдаемый недостаток гетерозигот в каждой из выборок как результат влияния только нуль-аллелей, то проведенная корректировка частот микросателлитных аллелей (данные не приведены из-за большого объема) показала статистически незначимо отличающиеся результаты от полученных без учета влияния нуль-аллеля. Этот результат позволяет провести оценку изменчивости и дифференциации совокупности субпопуляций соболя Среднего Приамурья на основании выявленного аллельного разнообразия.
В общем пуле 117 особей из Среднего При- амурья суммарно по трем локусам обнаружено 23 аллеля. В географических выборках (субпопуляциях) Буреинского нагорья обнаруживается от 16 до 18 аллелей, а в единственной выборке хребта Сихотэ-Алинь найдено 22 аллеля. Можно предположить, что преобладание числа аллелей в Сихотэ-Алинской выборке является результатом ее большего объема (30 экз.) по сравнению с отдельными буреинскими, которые представлены 14–28 экземплярами. Однако сравнение выборок Сихотэ-Алинской и суммарной Буреинского нагорья (87 экз.) указывает на реальное преобладание аллельного разнообразия у соболей Сихотэ-Алиня (22 против 19 аллелей).
Отметим, что 18 из 23 обнаруженных аллелей являются общими для соболей Буреинско-го нагорья и Сихотэ-Алиня. Более половины обнаруженных аллелей (13 из 23) представлены во всех географических выборках. Еще пять аллелей мозаично распределены по субпопуляциям обследуемой территории. Все эти аллели обнаружены в Сихотэ-Алинской субпопуляции, а кроме того и в одной (аллель 158 локуса Mer-41) либо нескольких субпопуляциях Буреинского нагорья (аллели 162, 174 локуса Ма-2, а также аллели 144, 156 локуса Mer-41). Как видно на рис. 2, суммарная доля общих аллелей очень высока и охватывает от 0,875 до 1,00 аллельного состава по соответствующим локусам в локальных субпопуляциях. Аналогичную картину преобладания общих фенотипов по субпопуляциям данной территории мы наблюдали при исследовании морфотипов строения небных складок соболя Приамурья [14].
5 из 23 обнаруженных аллелей являются специфическими, т.е. представлены в выборках соболя какой-либо одной из рассматриваемых горных систем. Аллель 135 локуса Ма-3 маркирует все рассматриваемые локальные субпопуляции Буреинского нагорья. Аллель 129 локуса Ма-3 и аллели 138, 160, 162 локуса Mer-41 маркируют субпопуляцию «Манома» хребта Сихотэ-Алинь. Доля специфического аллеля 135 в субпопуляциях Буреинского хребта изменяется от 0,083 до 0,125.
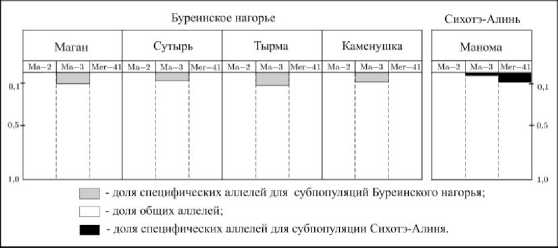
Рис. 2. Распределение общих и специфических аллелей в субпопуляциях Буреинского нагорья и Сихотэ Алиня
Fig.2. Distribution of common and specifi c alleles in sable subpopulations of the Bureinsky highlands and Sikhote-Аlin
Доля специфического аллеля субпоуляции «Ма-нома» (аллель 129) равна 0,017. Суммарная доля трех специфических аллелей локуса Mer-41, маркирующих субпопуляцию «Манома», равна 8,1%.
Количественная оценка уровня дифференциации проведена на основе показателей дифференциации субпопуляций - F(ST), полученных при парном сравнении рассматриваемых географических субпопуляций (табл. 4).
Полученные показатели соответствуют уровню слабой генетической дифференциации (F(ST) <0,05) по шкале Райта [23].
Расчет соответствующих F(ST) р -величин (p -value, проверка нулевой гипотезы об отсутствии генетической дифференциации между субпопуляциями, уровень значимости 0,05) показал, что из общего числа полученных коэффициентов данная гипотеза не отклоняется при парном сравнении субпопуляций Буреинского нагорья между собой. При парном сравнении субпопуляций Буреинского нагорья с субпопуляцией западного макросклона хребта Сихотэ-Алинь результат неоднозначен. Гипотеза об отсутствии генетический дифференциации не отклоняется при сравнении двух субпопуляций Буреинского нагорья («Сутырь» и «Каменушка») с субпопуляцией Сихотэ-Алиня («Манома»). Статистически значимая дифференциация обнаруживается при сравнении субпопуляций «Манома» – «Тырма» ( F(ST) = 0,022) и субпопуляций «Манома» - «Маган» ( F(ST) = 0,023) (табл. 4). Также статистически значимый уровень дифференциации показан для сравнений субпопуляции Сихотэ-Алиня «Мано-ма» с суммарной выборкой рассматриваемых четырех субпопуляций Буреинского нагорья («Маган» + «Сутырь» + «Тырма» + «Каменушка»). В
Таблица 4
Матрица значений F(ST) при парном сравнении локальных географических субпопуляций
Table 4
Matrix of FST) values in paired comparison of local geographic subpopulations
Обсуждение
Исследование изменчивости митохондриальных и ядерных генов представляет соболей Сихотэ-Алиня как наиболее высокополиморфную географическую часть вида. Сихотэ-Алинь – это предполагаемая территория соболиного рефугиума, откуда соболь расселялся по Северной Евразии. Сихотэ-Алинь, кроме того, – это территория, куда были интродуцированы животные двух подвидовых форм: из Прибайкалья и севера Буреинского нагорья. Вполне возможно, что эти два фактора совместно, как и каждый из них, могут объяснить увеличение генетического разнообразия в рамках совокупности соболей указанной территории.
Представленные в настоящей статье результаты исследования биологического материала одного охотничьего сезона указывают на то, что вывод о более высоком уровне разнообразия соболей Сихотэ-Алиня верен и для сравнения субпопуляции западного макросклона Сихотэ-Алиня с географически близкими субпопуляциями левобережья Амура (Буреинское нагорье). При рассмотрении трех микросателлитных локусов в Сихотэ-Алинской субпопуляции обнаружено аллельное разнообразие (22 аллеля), которое близко к аллельному разнообразию, представленному у исследованных соболей региона в целом (23 аллеля). Рассматриваемые географические субпопуляции левобережья Амура обладают меньшим аллельным разнообразием – по 16–18 аллелей в каждой. Меньшим аллельным разнообразием, чем в рассматриваемой Сихотэ-Алинской субпопуляции, характеризуется и суммарная выборка Буреинского нагорья (19 аллелей). Полученные значения парных F(ST) для субпопуляций левобережья (табл. 3) не противоречат нулевой гипотезе об отсутствии дифференциации между пространственно разобщенными группами обитателей Бу-реинского нагорья, что позволяет рассматривать географические субпопуляции Буреинского нагорья в рамках единой популяции.
При сравнении субпопуляций Буреинского нагорья и субпопуляции хребта Сихотэ-Алинь наблюдается несколько иная картина. Несмотря на значительное преобладание в рассматриваемых субпопуляциях животных с общими аллелями, обнаружены аллели, маркирующие совокупности соболей каждой из рассматриваемой территорий, разделенных рекой Амур. Однако частота встречаемости таких аллелей в субпопуляциях достаточно низкая. При парном сравнении субпопуляций Буреинского нагорья с субпопуляцией Сихотэ-
Алиня (Манома) гипотеза о нулевом уровне их дифференциации отклоняется для сравнений «Маган» – «Манома» и «Тырма» – «Манома» и не отклоняется для сравнений «Сутырь» – «Манома» и «Каменушка» – «Манома». Однако при сравнении суммарной выборки субпопуляций Буреин-ского нагорья и Сихотэ-Алинской субпопуляции гипотеза об отсутствии их дифференциации также отклоняется.
В островной модели Райт показал, что в отсутствии отбора и при низком уровне миграций на основании F(ST) можно оценить поток мигрантов между популяциями по соотношению
F ( ST ) =----1---,
-
v ’ 4 N e m + 1
где Ne – эффективный размер популяции, m – коэффициент миграции.
Согласно современным представлениям, микросателлиты адаптивно нейтральны. Если рассматривать водную преграду реку Амур как возможный изолирующий барьер, то можно (попытаться) представить рассматриваемые горные системы как аналог островов. В нашем случае, для показателя F(ST) =0,015 получаем Nm =16, т.е. поток мигрантов между рассматриваемыми горными системами оценивается в 16 экз. на поколение, что предполагает отсутствие тенденций к дифференциации, возможность которой оценивается величиной, равной не более чем одному мигранту на поколение [18]. Однако, к интерпретации величины Nm =16 приходится относиться с большой осторожностью, поскольку она объединяет суммарный вклад результатов естественного миграционного процесса, дрейфа генов и антропогенно созданных в не столь далеком прошлом волн жизни и потока мигрантов. Любой из этих факторов в отдельности или совместно способны обеспечивать изменение аллельных частот, т.е. влияют на величину F(ST) , а значит, и на Nm . «Парные оценки F(ST) дают некоторое представление о степени исторической связи популяций, но не позволяют определить, являются ли они результатом продолжающейся миграции или недавнего общего происхождения» [19].
На биологическом материале одного охотничьего сезона (зима 2012–2013) мы показали генетическое единство субпопуляций соболя Бу-реинского нагорья между собой и слабое, но статистически значимое отличие некоторых из них от субпопуляции хребта Сихотэ-Алинь.
Возникает вопрос – сохраняются ли во временной последовательности те статистически значимые различия, что мы наблюдаем при иссле- довании материала одного охотничьего сезона? Иными словами, являются ли эти наблюдаемые различия флуктуациями частот аллелей во времени либо мы обнаруживаем начальный этап дифференциации соболиных субпопуляций Буреинского нагорья и хребта Сихотэ-Алинь. Исследование материала нескольких последовательных охотничьих сезонов может помочь в поисках ответа на поставленный вопрос.
Авторы выражают искреннюю признательность Л.В. Капитоновой, Е.А. Ерофеевой и П.В. Будилову за помощь в сборе материала, а Е.А. Ерофеевой благодарность за помощь в графическом оформлении собранного материала.
Настоящее исследование выполнено при частичной финансовой поддержке проекта № 18-05-51 программы ДВО РАН «Дальний Восток».
Список литературы Генетическая структура популяции соболя ( Martes zibellina ) Буреинского нагорья по сборам одного охотничьего сезона
- Абрамов К.Г. Соболь в охотничьем хозяйстве Дальнего Востока. М.: Наука, 1967. 116 с
- Балмышева Н.П., Соловенчук Л.Л. Генетическая изменчивость гена итохрома b митохондриальной ДНК соболя (Martes zibellina L.) магаданской популяции // Генетика. 1999. Т. 35, № 9. С. 1252-1257
- Голобокова О.А. История создания Верхнебуреинского соболиного племенного рассадника. 2017. URL: https://komza.khabkrai.ru/Municipalnye-arhivy/Novosti-/890 (дата обращения: 22.11.2019)
- Каплин А.А. Пушнина СССР. М.: Внешторгиздат, 1960. 464 с
- Каштанов С.Н., Рубцова Г.А., Лазебный О.Е. Исследование генетической структуры промышленной популяции соболя (Martes zibellina Linnaeus,1758) по микросателитным маркерам // Вестник ВОГиС. 2010. Т. 14, № 3. С. 426-431
- Каштанов С.Н., Свищева Г.Р., Лазебный О.Е., Колобков Д.С., Пищулина С.Л., Мещерский И.Г., Рожнов В.В. Влияние антропогенных факторов на генетическое разнообразие вида соболь (Martes zibellina L.) // Молекулярная биология. 2015. Т. 49, № 3. С. 449-454
- Кордичева С.Ю., Рубцова Г.А., Шитова М.В. и др. Выявление нуль-аллелей в микросателлитном локусе кеты (Oncorhynchus keta Walbaum) // Генетика. 2010. Т. 46, № 8. С. 1143-1147
- Кузнецов В.М. F-статистика Райта: оценка и интерпретация // Проблемы биологии продуктивных животных. 2014. № 4. C. 80-104
- Монахов В.Г. Динамика размерной и фенетической структуры соболя в ареале. Екатеринбург, 2006. 201 с
- Монахов В.Г., Модоров М.В., Ранюк М.Н. Генетические последствия реинтродукции соболя (Martes zibellina L.) в Западной Сибири // Генетика. 2018. Т. 54, № 3. С. 361-365
- Петровская А.В. Генетическая структура популяций соболя в Магаданской области по данным об изменчивости митохондриальной ДНК // Генетика. 2007. Т. 43, № 4. С. 530-536
- Ранюк М.Н., Монахов В.Г. Изменчивость краниологических признаков в популяциях соболя (Martes zibellina), возникшая в результате акклиматизации // Зоологический журнал. 2011. Т. 90, № 1. С. 82-96
- Рожнов В.В., Пищулина С.Л., Мещерский И.Г. и др. Генетическая структура соболя (Martes zibellina L.) Евразии и анализ распределения митохондриальных линий // Генетика. 2013. Т. 49, № 2. С. 251-258
- Фрисман Л.В., Капитонова Л.В. Изменчивость небных складок у соболя ((Martes zibellina) Среднего Приамурья // Региональные проблемы. 2018. Т. 21, № 2. С. 39-51
- Шишацкая Д.И., Фрисман Л.В. Предварительные данные по генетической изменчивости соболя Среднего Приамурья (Martes zibellina L.): анализ двух микросателлитных локусов // Региональные проблемы. 2014. Т. 17, № 2. С. 60-64
- Aljanabi S.M., Martinez I. Universal and rapid salt-extraction of high quality genomic DNAb for PCR-based techniques // Nuclec Acids Research. 1997. Vol. 25, N 22. Р. 4692-4693
- Excoffier L., Laval G., Schneider C. Arlequin. Ver 3.0. An integrated software package for population genetics data analisys // Evolutionary Bioinformatic Online. 2005. 1. P. 47-50
- Holsinger K.E. Lecture notes in population genetics. Storrs: University of Connecticut, 2010. 275 p
- Holsinger K.E., Weir B.S. Genetics in geographically structured populations: defining, estimating and interpreting Fst // Nature Reviews Genetics. 2009. Vol. 10(9). P. 639-650
- Kinoshita Gohta, Sato J.J., Meschersky I.G., Pishchulina Sofiko L., Simakin L.V., Rozhnov V.V., Malyarchuk B.A., Derenko M.V., Denisova G.A., Frisman L.V., Kryukov A.P., Hosoda Tetsuji, Suzuki Hitoshi. Colonization history of the sable Martes zibellina (Mammalia, Carnivora) onthe marginal peninsula and islands of northeastern Eurasia // Journal of Mammalogy. 2015. N 96(1). P. 172-184
- Sato J.J., Hosoda T., Kryukov A.P., Kartavtseva I.V., Suzuki H. Genetic diversity of the sable (Martes zibellina, Mustelidae) in Russian Far East and Hokkaido inferred frommitochondrial NADH dehydrogenase subunit 2 gene sequences // Mammal Study 2011. N 36. P. 209-222
- Van-Oosterhout C., Hutchinson W.F., Wills D.P., Shipley P. Micro-checker: software for identifying and correcting genotyping errors in microsatellite data // Mol. Ecol. Notes. 2004.Vol. 4, N 3. P. 535-538
- Wright S. Evolution and Genetics of Population. V. 4. Variability Within and Among Natural Populations. Chicago: Univ. Chicago Press, 1978. 590 p
