Генетическая структура региональных популяций Mycosphaerella graminicola (Septoria tritici) - возбудителя септориоза пшеницы (Triticum aestivum L.)
 - возбудителя септориоза пшеницы (Triticum aestivum L.)")
Автор: Пахолкова Е.В., Сальникова Н.Н., Куркова Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Фитопатология. Иммунитет и защита растений
Статья в выпуске: 5 т.51, 2016 года.
Бесплатный доступ
Возбудитель септориозной листовой пятнистости (STB) Mycosphaerella graminicola (анаморфа Septoria tritici ) - доминирующий вид в септориозном комплексе на посевах пшеницы в основных зернопроизводящих районах России. Устойчивость к данному заболеванию может быть как количественной (горизонтальной), так и изолят-специфичной (вертикальной). В настоящее время у пшеницы идентифицировано 17 генов устойчивости к STB ( Stb1 - Stb17 ). Благодаря генетическому анализу в патосистеме «пшеница- M. graminicola » было доказано взаимодействие «ген-на-ген», следовательно, наличие генов устойчивости у хозяина предполагает существование специфических генов вирулентности у патогена. Относительная частота генов вирулентности внутри географического региона может быть вычислена как доля изолятов, выражающих эти гены, от общего числа изолятов, используемых в исследовании. Целью нашей работы было, допуская связь «ген-на-ген», определить гены вирулентности в популяциях M. graminicola из разных географических регионов России, используя сорта с известными генами устойчивости - Bulgaria 88 ( Stb1 ), Oasis ( Stb1 ), Veranopolis ( Stb2 ), Israel 493 ( Stb3 ), Tadinia ( Stb4 ), CS/Synthetic 7D ( Stb5 ), Flame ( Stb6 ), Estanzuela Federal ( Stb7 ), W7984 ( Stb8 ). В условиях теплицы и камеры искусственного климата были протестированы 47 изолятов из Северо-Кавказского района, 66 изолятов из Центрально-Черноземного района, 29 изолятов из Поволжского района, 64 изолята из Центрального района и 34 изолята из Северо-Западного района. Оценку вирулентности изолятов проводили на всходах в фазу 2 листьев по двум параметрам - степени поражения растений и споруляции гриба in vivo. На основании частоты встречаемости генов вирулентности была выявлена степень эффективности Stb -генов по отношению к каждой региональной популяции гриба. Региональные популяции M. graminicola различались по генотипу вирулентности, спектру и частоте встречаемости генов вирулентности. Популяции из южной зоны (северо-кавказская, центрально-черноземная и поволжская) обладали более высокой вирулентностью по сравнению с центральной и северо-западной популяциями. Так, в северо-кавказской популяции не имели генов вирулентности 19,2 % изолятов, в центрально-черноземной - 6,0 %, тогда как в центральной и северо-западной их доля составляла соответственно 42,2 и 44,1 %. Изоляты из поволжской популяции M. graminicola имели наиболее разнообразные сочетания генов вирулентности. Во всех популяциях выявлена высокая частота вирулентности в отношении сортов с генами Stb1, Stb5 и Stb7. Значительной эффективностью против изолятов из Центрального, Центрально-Черноземного и Северо-Западного районов характеризовались гены Stb2, Stb3, Stb4, однако она заметно снижалась в отношении северо-кавказских изолятов и изолятов из Поволжского района. Гены Stb6 и Stb8 были в высокой степени эффективны ( Stb8 - абсолютно эффективен) против всех региональных популяций M. graminicola и могут быть рекомендованы для селекции на устойчивость к STB.
Популяция, изолят, гены вирулентности, частота встречаемости, эффективность stb-генов
Короткий адрес: https://sciup.org/142213976
IDR: 142213976 | УДК: 633.1:632.4.01/06:575.174 | DOI: 10.15389/agrobiology.2016.5.722rus
Genetic structure of regional populations of Mycosphaerella graminicola (Septoria tritici), the septoria leaf blotch agent of wheat
Mycosphaerella graminicola (anamorph Septoria tritici ), the causal agent of septoria tritici blotch (STB) of wheat, is dominating species in Septoria / Stagonospora complex on crops in the main grain-producing areas of Russia. Resistance to STB may be either quantitative (horizomtal) or isolate-specific (vertical). At present 17 genes for resistance have been identified ( Stb1 - Stb17 ). The gen-for-gen interaction in the «wheat- M. graminicola » pathosystem has been demonstrated by genetic analysis; therefore, the aviability of resistance genes in the host proposes the existence of specific virulence genes in the pathogen. The relative frequency of virulence genes within a geographic region may be calculated as a fraction of the isolates expressing this virulence genes from the overall number of isolates used in the study. The purpose of the present study was to estimate the virulence genes in populations of M. graminicola from different geographic regions of Russia on the basis of a gen-for-gen relationship, using the cultivars with known resistance genes, i.e. Bulgaria 88 ( Stb1 ), Oasis ( Stb1 ), Veranopolis ( Stb 2), Israel 493 ( Stb 3), Tadinia ( Stb4 ), CS/Synthetic 7D ( Stb5 ), Flame ( Stb6 ), Estanzuela Federal ( Stb7 ), W7984 ( Stb8 ). A total of 47 isolates from the North-Caucasian region, 66 isolates from the Central-Chernozem region, 29 isolates from the Volga region, 64 isolates from the Central region, and 34 isolates from the North-West region were tested under greenhouse and grows chamber conditions. The virulence was estimated on seedlings at two-leaf stage, using two parameters, the infection degree of plants and sporulation of fungus in vivo. The effectiveness of Stb -genes to each regional population of M. graminicola was revealed on the basis of the frequency of virulence genes. The regional populations of M. graminicola differed in virulence genotype, spectrum and frequency of virulence genes. The populations from south zone (the North-Caucasian, the Central-Chernozem and the Volga regions) are more virulent in comparison with the central and the north-west populations. For example, 19.2 % of isolates from the north-caucasian population and 6.0 % of isolates from the central-chernozem population have no virulence genes, while in the central and north-west populations - 42.2 % and 44.1 %, respectively. Isolates from the Volga population of M. graminicola had most various combinations of virulence genes. High frequency of virulence to genes Stb1, Stb5 and Stb7 was revealed in all populations. The genes Stb2, Stb3, Stb4 have considerable effectiveness to the central, the central-chernozem and the north-west populations of M. graminicola, however it distinctly reduced concerning isolates from the North-Caucasian and the Volga regions. The genes Stb6 and Stb8 were highly effective ( Stb8 - absolutely effective) to all investigated Russian populations of M. graminicola and may be recommended for using in selection as sources of resistance to STB.
Текст научной статьи Генетическая структура региональных популяций Mycosphaerella graminicola (Septoria tritici) - возбудителя септориоза пшеницы (Triticum aestivum L.)
Возбудитель септориозной листовой пятнистости (STB) Mycosphaerel-la graminicola (анаморфа Septoria tritici ) — доминирующий вид в септориоз-ном комплексе на посевах пшеницы в основных зернопроизводящих районах России (Северо-Кавказский, Центрально-Черноземный). Кроме того, он широко распространен в центральной части страны и на юге Поволжья, где его доля в видовой структуре популяций достигает 40-50 %. Отмечается присутствие этого вида в Северо-Западном районе, в Калининградской области, а также на территории Западной и Восточной Сибири.
Знание генетической структуры популяций патогена, частоты встречаемости генов вирулентности, их динамики во времени и пространстве, а также степени эффективности генов устойчивости относят к обязательным
∗ Работа выполнена при финансовой поддержке гранта Международного научно-технического центра (МНТЦ).
условиям успешной селекции на устойчивость к заболеванию. Если сравнивать с другими болезням (ржавчина, мучнистая роса и др.), то в случае септориоза генетика устойчивости растений менее изучена. К тому же в патосистеме «пшеница― M. graminicola » присутствуют два типа взаимодействия, то есть устойчивость может быть как количественной (горизонтальной), так и изолят-специфичной (вертикальной), причем оба эти взаимодействия важны для патосистемы.
Специфичность взаимодействия между M. graminicola и пшеницей впервые доказали Z. Eyal с соавт. (1, 2), которые, проанализировав многие индивидуальные комбинации «сорт ½ изолят», предположили наличие 28 комплементарных генов. Существование взаимодействующих пар генов было подтверждено результатами других исследователей с использованием 80 изолятов и 47 сортов (3). Заключительное свидетельство связи «ген-на-ген» было получено с помощью молекулярно-генетических исследований хозяина и патогена, что устранило любые сомнения в ее существовании в отношении по крайней мере некоторых известных генов устойчивости (4-7).
За недавние годы на основании взаимодействия между сортами пшеницы и изолятами гриба идентифицировано 17 основных генов устойчивости к M. graminicola ( Stb1 - Stb17 ). Эксперименты, проведенные в разных лабораториях мира, позволили определить хромосомную локализацию и молекулярные маркеры этих генов, которые предлагается использовать в маркерной селекции (8-14).
Согласно теории «ген-на-ген», наличие генов устойчивости у хозяина предполагает существование специфических генов вирулентности у патогена, частота встречаемости которых в регионе может быть вычислена как доля изолятов, выражающих эти гены (вирулентности), от числа изо-лятов, используемых в исследовании (2). Это позволяет, в свою очередь, определить степень эффективности Stb -генов, что имеет большое значение для селекции, так как внедрение определенного Stb -гена не обусловит эффективную устойчивость, если часть популяции патогена вирулентна для несущего такой ген сорта. В частности, исследования, проведенные во Франции, показали, что при тестировании 11 сортов с известными генами устойчивости с использованием монопикнидиальных изолятов из пяти областей страны многие из Stb -генов были неэффективными в отношении большинства французских штаммов M. graminicola (15). Тем не менее, эффективность некоторых генов подтверждена мировой практикой. Так, ген Stb1 , введенный в озимые сорта пшеницы Oasis и Sullivan, более чем 25 лет сохранял свою эффективность в Индиане и соседних штатах (8). Сорт яровой пшеницы Tadinia обладает одним доминантным геном устойчивости Stb4 , который в течение 30 лет использовался для контроля S. tritici в Калифорнии (9). Ген Stb6 ― широко распространенный источник устойчивости к STB во всем мире (16). Об эффективности Stb -генов по отношению к российским популяциям патогена известно мало. Есть только данные Ю.В. Зеленевой (17), проводившей исследования на изолированных листьях пшеницы, согласно которым наибольшей эффективностью против изо-лятов M. graminicola из Центрально-Черноземного района обладали гены Stb1 , Stb4 , Stb5 , тогда как гены Stb2 , Stb3 были менее эффективными.
Пока что нет согласованной методологии оценки и определения границы разделения устойчивости и восприимчивости. Известно, что иммунитета (полной устойчивости) к M. graminicola не существует, так как некрозы и (или) пикниды присутствуют всегда (18, 19). Генетическая устойчивость к STB может выражаться в уменьшении размера зоны поражений и в снижении плодовитости гриба. Эти два параметра находятся под разным генетическим контролем и оба важны при оценке заболевания (3). Плодовитость гриба чаще всего принято оценивать по числу пикнид. В этом случае реакция растений группируется в классы от близкой к иммунной с небольшими некрозами без пикнид до очень восприимчивой с большими сливающимися пятнами и многочисленными пикнидами (5, 2022). Однако визуальная оценка числа пикнид, особенно по балльной шкале, довольно субъективна. Кроме того, было установлено, что на восприимчивых сортах число спор в одной пикниде в 2,0-2,5 раза больше, чем на устойчивых (23). В связи с этим некоторые исследователи рекомендуют определять спорулирующую способность гриба in vivo методом подсчета спор в камере Горяева (24).
Целью наших исследований было, допуская связь «ген-на-ген», определить гены вирулентности и частоту их встречаемости в популяциях Mycosphaerella graminicola из разных регионов России, используя набор сортов с известными генами устойчивости. Эта информация позволит выявить эффективность Stb -генов, оценить их функциональность на территории России и целесообразность использования в селекционных программах в качестве потенциальных источников устойчивости к STB.
Методика. Исследования проводили в течение 2009-2015 годов. Образцы пораженных растений собирали во время обследования посевов пшеницы в течение вегетационного периода по стандартной методике (25). При выделении в чистую культуру моноспоровых изолятов Myco-sphaerella graminicola применяли метод штрихов (26). Для этого кусочки пораженной ткани с пикнидами гриба сначала промывали под проточной водой, затем в нескольких порциях стерильной дистиллированной воды и помещали в стерильную чашку Петри на предметное стекло в каплю стерильной воды. Через несколько минут полученную споровую суспензию с помощью проволочной петли высевали на поверхность питательной среды в чашки Петри. В качестве питательной среды использовали картофельноглюкозный агар (КГА). Через 7-8 сут инкубации при температуре 20-25 ° С единичные колонии, произошедшие из одной конидии, отсевали в другие чашки. Стабильность изолятов отслеживали при трех последовательных пересевах 10-суточных колоний на свежую питательную среду. Культурально-морфологические (КМ) признаки изолятов оценивали на 30-е сут после посева, отмечая характер строения, диаметр и окраску колоний. Для исследований отбирали стабильные по КМ признакам изоляты из разных морфологических групп, стараясь как можно полнее отразить внутривидовое разнообразие популяций.
В опытах использовали моногенные сорта пшеницы с известными генами устойчивости — Bulgaria 88 и Oasis ( Stb1 ), Veranopolis ( Stb2 ), Israel 493 ( Stb3 ), Tadinia ( Stb4 ), CS/Synthetic 7D ( Stb5 ), Flame ( Stb6 ), Estanzuela Federal ( Stb7 ) и W7984 ( Stb8 ). Исследования проводили в камерах искусственного климата и в теплице. Растения выращивали в вазонах объемом 400 см3 (по 10 шт. на вазон) до фазы полностью развернутого 2-го листа. Инокулюм гриба культивировали на КГА в чашках Петри в течение 4-5 сут в лабораторных условиях без дополнительного освещения. Растения заражали индивидуальными изолятами, опрыскивая споровой суспензией гриба с помощью пульверизатора. Плотность суспензии ― 1½107 спор/мл, расход суспензии ― 100 мл/м2. Перед нанесением в суспензию добавляли каплю поверхностно-активного вещества Твин 20. После инокуляции растения на 48 ч помещали во влажную камеру при температуре 20-25 ° С, затем переносили в ростовую камеру или бокс, где поддерживались следующие условия: температура 18-20 ° С (ночь) и 22-24 ° С 724
(день), относительная влажность воздуха 70-80 %, фотопериод 16 ч, освещенность ― около 15 тыс. лк.
Оценку пораженности растений проводили через 20 сут после инокуляции. За основу был взят разработанный во Всероссийском НИИ фитопатологии метод, согласно которому в качестве основных параметров использовали степень поражения растений и интенсивность споруляции гриба in vivo (24, 27). Степень поражения определяли визуально по проценту пораженной поверхности 1-го и 2-го листа. Для определения споруляции срезали учетные листья растений и помещали их на 3-4 ч в лабораторные стаканы с точно измеренным количеством воды. Затем с помощью камеры Горяева определяли число спор в суспензии в пересчете на один учетный лист: N = 2500 MV/n, где N — число спор на одном учетном листе; M — число спор в 100 больших квадратах камеры Горяева; V — объем воды в сосуде, мл; n — число листьев в пробе; 2500 — коэффициент, полученный экспериментально.
Степень поражения дифференцировали как низкую (поражено в среднем до 20 % площади листа), среднюю (21-50 %) или высокую (более 50 %). По интенсивности споруляции изоляты относили к слабоспорулиру-ющим (до 100 тыс. спор/лист), среднеспорулирующим (от 100 до 200 тыс. спор/лист) или высокоспорулирующим (более 200 тыс. спор/лист). На основании сочетания двух этих параметров изоляты делили на три группы: I — слабовирулентные, II — средневирулентные, III — высоковирулентные.
По проценту вирулентных изолятов от общего числа исследованных определяли частоту и спектр генов вирулентности в популяции, а также оценивали эффективность Stb -генов. Stb -гены условно разделяли на эффективные (сорт восприимчив к 0-20 % изолятов), среднеэффективные (сорт восприимчив к 21-50 % изолятов) и неэффективные (сорт восприимчив более чем к 50 % изолятов).
Результаты. Группам вирулентности у изученных изолятов соответствовали следующие сочетания интенсивности споруляции гриба и степени поражения растений (табл. 1). Изоляты I группы рассматривались как авирулентные, тогда как II или III группа вирулентности свидетельствовали о наличии у изолята гена вирулентности.
1. Распределение изученных изолятов Mycosphaerella graminicola по группам вирулентности с учетом споруляции и развития поражения у растений пшеницы ( Triticum aestivum L.) в лабораторных тестах
|
Средняя степень поражения растений, % |
Интенсивность споруляции, тыс. спор/лист |
||
|
низкая (до 100) |
средняя (100-200) |
высокая (более 200) |
|
|
Низкая (до 20) |
I |
I |
II |
|
Средняя (21-50) |
I |
II |
III |
|
Высокая (51-100) |
II |
III |
III |
При исследовании генетической структуры популяций M. gramini-cola протестировали 47 изолятов из Северо-Кавказского района (14 — из Северной Осетии, 19 — из Ставропольского края, 13 — из Краснодарского края, 1 — из Чечни); 66 изолятов из Центрально-Черноземного района (26 — из Воронежской, 10 — из Тамбовской, 10 — из Липецкой, 12 — из Курской, 8 — из Белгородской области); 29 изолятов из Поволжья (9 — из Саратовской и 20 — из Волгоградской области); 64 изолята из Центрального района (59 — из Московской, 3 — из Тульской, 1 — из Брянской и 1 — из Рязанской областей); 34 изолята из Северо-Западного района (6 — из Ленинградской, 9 — из Псковской, 19 — из Новгородской областей). Все изоляты тестировали на восьми сортах с моногенной устойчивостью, за исключением представителей северо-кавказской популяции, которые мы испытывали на шести сортах, поскольку на тот период не располагали сортами с генами Stb6 и Stb8.
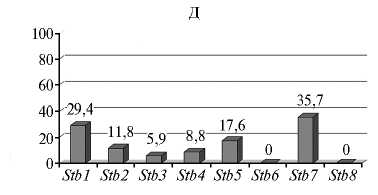
Во всех популяциях M. graminicola была выявлена вирулентность в отношении генов устойчивости Stb1 , Stb2 , Stb3 , Stb4 , Stb5 и Stb7 , значительно варьирующая по частоте (рис.).
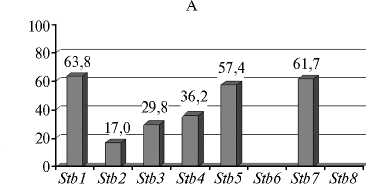
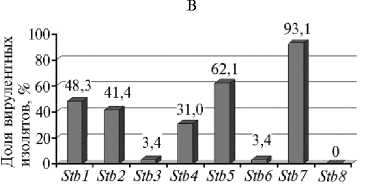
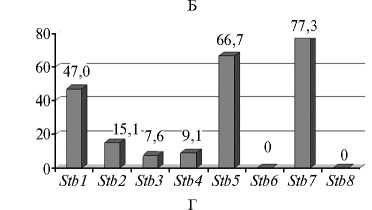
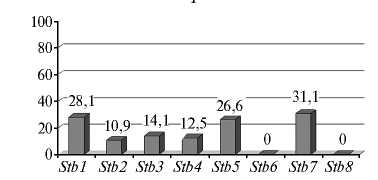
Частота проявления вирулентности в отношении Stb -генов устойчивости пшеницы ( Triticum aestivum L.) в популяциях Mycosphaerella grami-nicola из разных районов России : А — СевероКавказский, Б — Центрально-Черноземный, В — Поволжский, Г — Центральный, Д — Северо-Западный (лабораторные тесты, образцы сборов 2009-2015 годов).
Северо-кавказская популяция была представлена различными генотипами вирулентности, то есть изоляты несли разные комбинации соответствующих генов. При этом 55,3 % изолятов проявляли широкий спектр вирулентности (поражали 3-6 моногенных сортов), 25,5 % изолятов имели по 1-2 гена вирулентности. Изоляты, не обладающие генами вирулентности, составляли 19,2 %. Чаще других поражались сорта c генами устойчивости Stb1 , Stb5 и Stb7 : доля вирулентных для них изолятов составляла 57,4-63,8 %. Сорта с генами Stb2 , Stb3 и Stb4 поражались реже и были восприимчивы к 17,0-36,2 % изолятов.
В центрально-черноземной популяции у 50 % изолятов выявили только по 1-2 гена вирулентности. Доля изолятов с более широким спектром вирулентности составляла 43,9 %. Не имели генов вирулентности 6,0 % изолятов. Чаще других поражались сорта c генами устойчивости Stb1 , Stb5 и Stb7 (47,0-77,3 % вирулентных изолятов). Сорта с генами Stb2 , Stb3 и Stb4 были устойчивы против большинства изолятов (частота вирулентности 7,6-15,1 %). Вирулентность в отношении генов устойчивости Stb6 и Stb8 отсутствовала.
Изоляты из поволжской популяции M. graminicola имели наиболее разнообразные сочетания генов вирулентности. При этом только один изолят (3,4 %) был авирулентным для всех сортов, а 13,8 % изолятов имели по одному гену вирулентности. Большинство изолятов несли по 3-6 генов вирулентности (65,5 %), подавляющее большинство (93,1 %) были вирулентны для сорта с геном Stb7 . Частота вирулентности в отношении сортов с геном Stb5 составляла 62,1 %, с генами Stb1 , Stb2 и Stb4 ― 31,0726
48,3 %. Незначительная вирулентность (3,4 %) наблюдалась в случае Stb3 и Stb6 . Сорт с геном Stb8 был устойчив ко всем изолятам из этой популяции.
В популяции из Центрального района 42,2 % изолятов не имели генов вирулентности. У остальных главным образом выявлялось по 1-2 гена (40,6 % изолятов), реже по 3-5 генов (17,2 % изолятов). В целом, частота вирулентности в отношении Stb -генов была невысокой. Так, для Stb1 , Stb5 и Stb7 она составляла 26,6-31,1 %, для Stb2 , Stb3 и Stb4 — была вдвое ниже (10,9-14,1 %), для Stb6 и Stb8 — равнялась 0 %.
Северо-западная популяция M. graminicola была наименее разнообразной по генотипам вирулентности. Основное число изолятов не имели генов вирулентности (44,1 %) либо имели по одному гену вирулентности (32,3 %). Однако были и изоляты, которые несли по 3-6 генов, но их доля оказалась значительно меньше (14,7 %). Вирулентность наблюдалась чаще всего к генам устойчивости Stb1 и Stb7 (29,4-35,7 %). В отношении сортов с генами Stb2 , Stb3 , Stb4 и Stb5 частота вирулентности была заметно ниже и составляла от 5,9 до 17,6%. Ни у одного из изолятов не зафиксировали вирулентность к генам Stb6 и Stb8 .
Таким образом, региональные популяции M. graminicola различались по генотипам вирулентности, спектру и частоте встречаемости генов вирулентности. Вирулентность популяций из южной зоны (северо-кавказская, центрально-черноземная и поволжская) в целом была выше по сравнению с центральной и северо-западной популяциями, где наблюдалось высокое число авирулентных изолятов. Кроме того, популяции из южной зоны выделялись разнообразием генотипов вирулентности и более высоким числом изолятов с широким спектром вирулентности (3-6 генов). Такое географическое распределение вирулентности M. graminicola на территории России указывает на то, что южные популяции гриба более агрессивны, и это может быть одной из причин доминирования M. graminicola в септориозном комплексе на посевах пшеницы на юге страны.
2. Степень эффективности Stb -генов пшеницы ( Triticum aestivum L.) против региональных российских популяций Mycosphaerella graminicola (лабораторные тесты, образцы сборов 2009-2015 годов)
|
Регион |
Эффективность Stb -генов (по проценту вирулентных изолятов) |
||
|
эффективные (< 20 %) |
среднеэффективные (20-50 %) |
неэффективные (> 50 %) |
|
|
Северо-Кавказский |
Stb2 |
Stb3, Stb4 |
Stb1, Stb5, Stb7 |
|
Центрально-Черноземный Stb2, Stb3, Stb4, Stb6, Stb8 |
Stb1 |
Stb5, Stb7 |
|
|
Поволжский |
Stb3, Stb6, Stb8 |
Stb1, Stb2, Stb4 |
Stb5, Stb7 |
|
Центральный |
Stb2, Stb3, Stb4, Stb6, Stb8 |
Stb1, Stb5, Stb7 |
Отсутствуют |
|
Северо-Западный |
Stb2, Stb3, Stb4, Stb5, Stb6, Stb8 |
Stb1, Stb7 |
Отсутствуют |
Во всех популяциях M. graminicola наблюдалась более высокая частота вирулентности в отношении сортов с генами устойчивости Stb1, Stb5 и Stb7, свидетельствующая о том, что эти Stb-гены наименее эффективны (табл. 2). Реакция растений обычно выражалась в наличии некротических пятен с большой площадью поражения листа и формированием пикнид при средней и высокой интенсивности споруляции. В то же время значительной эффективностью против центральной, центрально-черноземной и северо-западной популяций M. graminicola обладали гены Stb2, Stb3, Stb4, однако отмечалось снижение их эффективности при заражении растений изолятами из северо-кавказской и поволжской популяций. Ген Stb6 был в высокой степени эффективным против всех пяти популяций M. graminicola. Только в поволжской популяции выявили вирулентность для сорта с этим геном, но она встречалась с низкой частотой (3,4 %). Ген Stb8 был абсолютно эффективным против всех протестированных изолятов. Растения редко имели видимые симптомы поражения, споруляция была очень скудной либо отсутствовала.
Итак, наши исследования показали, что 5 из 8 известных генов устойчивости ( Stb1 - Stb5 ) обладают только частичной функциональностью в природных популяциях Mycosphaerella graminicola в России, а эффективность гена Stb7 не подтвердилась в отношении всех изученных популяций. В то же время идентифицированы потенциальные источники устойчивости к септориозной листовой пятнистости — это сорта с генами Stb6 и Stb8 , которые могут быть рекомендованы для использования в селекционно-генетических программах по созданию форм, устойчивых к M. graminicola на территории Российской Федерации.
Авторы благодарят Dr. S.B. Goodwin (USDA-ARS, Crop Production and Pest Control Research Unit, Small Grain Fungal Disease Laboratory, США) за любезно предоставленные семена сортов с известными генами устойчивости Stb1-Stb8.
Список литературы Генетическая структура региональных популяций Mycosphaerella graminicola (Septoria tritici) - возбудителя септориоза пшеницы (Triticum aestivum L.)
- Eyal Z., Amari Z., Whal I. Physiological Specialization of Septoria tritici. Phytopathology, 1973, 63: 1087-1091.
- Eyal Z., Scharen A.L., Huffman M.D., Prescott J.M. Global insights into virulence frequencies of Mycosphaerella graminicola. Phytopathology, 1985, 75: 1456-1462.
- Kema G.H.J., Annone J.G., Sayoud R., Van Silfhout C.H., Van Gincel M., de Bree J. Genetic variation for virulence and resistance in the wheat-Mycosphaerella graminicola pathosystem. I. Interaction between pathogen isolates and host cultivars. Phytopathology, 1996, 86: 200-212.
- Kema G.H.J., Verstappen E.C.P., Waalwijk C. Avirulence in the wheat Septoria tritici leaf blotch fungus Mycosphaerella graminicola is controlled by a single locus. Mol. Plant-Microbe Interact., 2000, 13: 1375-1379 ( ) DOI: 10.1094/MPMI.2000.13.12.1375
- McCartney C.A., Brule-Babel A.L., Lamari L. Inheritance of race-specific resistance to Mycosphaerella graminicola in wheat. Phytopathology, 2002, 92: 138-144 ( ) DOI: 10.1094/PHYTO.2002.92.2.138
- Kema G.H.J., Goodwin S.B., Hamza S., Verstappen E.C.P., Cavaletto J.R., van der Lee T.A.J., Hagenaar-de Weerdt M., Bonants P.J.M., Waalwijk C. A combined AFLP and RAPD genetic linkage map of Mycosphaerella graminicola, the septoria tritici leaf blotch pathogen of wheat. Genetics, 2002, 161: 1497-1505.
- Brading P.A., Verstappen E.C.P., Kema G.H.J., Brown K.M. A gene-for-gene relationship between wheat and Mycosphaerella graminicola, the Septoria tritici blotch pathogen. Phytopathology, 2002, 92: 439-445 ( ) DOI: 10.1094/PHYTO.2002.92.4.439
- Adhikari T.B., Anderson J.M., Goodwin S.B. Identification and molecular mapping of a gene in wheat conferring resistance to Mycosphaerella graminicola. Phytopathology, 2003, 93: 1158-1164 ( ) DOI: 10.1094/PHYTO.2003.93.9.1158
- Adhikari T.B., Cavaletto R., Dubcovsky J., Giego J., Schlatter A., Goodwin S.B. Molecular mapping of the Stb4 gene for resistance to Septoria tritici blotch in wheat. Phytopathology, 2004, 94: 1198-1206 ( ) DOI: 10.1094/PHYTO.2004.94.11.1198
- Adhikari T.B., Wallwork H., Googwin S.B. Microsatellite markers linked to the Stb2 and Stb3 genes for resistance to Septoria tritici blotch in wheat. Crop Sci., 2004, 44: 1403-1411 ( ) DOI: 10.2135/cropsci2004.1403
- Arraiano L.S., Chartrain L., Bossolini E., Slatter H.N., Keller B., Brown J.K.M. A gene in European wheat cultivars for resistance to an African isolate of Mycosphaerella graminicola. Plant Pathol., 2007, 56: 73-78 ( ) DOI: 10.1111/j.1365-3059.2006.01499.x
- Chartrain L., Sourdille P., Bernard M., Brown J.K.M. Identification and location of Stb9, a gene for resistance to Septoria tritici blotch in wheat cultivars Courtot and Tonic. Plant Pathol., 2009, 58: 547-555 ( ) DOI: 10.1111/j.1365-3059.2008.02013.x
- Liu Y., Zhang L., Thompson I.A., Goodwin S.B., Ohm H. Molecular mapping re-locates the Stb2 gene for resistance to Septoria tritici blotch derived from cultivar Veranopolis on wheat chromosome 1BS. Euphytica, 2012, 190: 145-156 ( ) DOI: 10.1007/s10681-012-0796-8
- Goodwin S.B., Cavaletto J.R., Hale I.L., Thompson I., Xu S.S., Adhikari T.B., Dubcovsky J. A new map location of gene Stb3 for resistance to Septoria tritici blotch in wheat. Crop Sci., 2015, 55: 35-43 ( ) DOI: 10.2135/cropsci2013.11.0766
- Tabib Ghaffary M.S., Laurent V., Guerreiro L. et al. Unravelling and exploitation of diversity for resistance to Mycosphaerella graminicola and Fusarium graminearum in wheat and its progenitors. In: Actes de la rencontre scientifique «Synthèse des programmes de recherché FSOV (Fonds de Soutien à l'Obtention Végétale)» (8 janvier 2008, Paris, France). Groupement national interprofessionnel des semences et plants (GNIS), Section Céréales et Protéagineux, Paris, 2009: 31-38 (ISBN 9782952964401).
- Chartrain L., Brading P.A., Brown J.K.M. Presence of the Stb6 gene for resistance to Septoria tritici blotch (Mycosphaerella graminicola) in cultivars used in wheat-breeding programmes worldwide. Plant Pathol., 2005, 54: 134-143 ( ) DOI: 10.1111/j.1365-3059.2005.01164.x
- Зеленева Ю.В., Судникова В.П. Влияние сорта на патогенные свойства возбудителя Septoria tritici Rob. et Desm. в условиях Центрального Черноземья. Вопросы современной науки и практики, 2014, 2(51): 15-20.
- Nelson L.R., Marshall D. Breeding for resistance to Septoria nodorum and Septoria tritici. Adv. Agron., 1990, 44: 257-277.
- Kema G.H.J., Yu D.Z., Rijkenberg F.H.J., Shaw M.W., Baayen R.P. Histology of the pathogenesis of Mycosphaerella graminicola in wheat. Phytopathology, 1996, 86: 777-786.
- Somasco O.A., Quaiset C.O., Gilchrist D.G. Single-gene resistance to Septoria tritici blotch in the spring wheat cultivar ‘Tadinia’. Plant Breeding, 1996, 115: 261-267 ( ) DOI: 10.1111/j.1439-0523.1996.tb00914.x
- Rosielle A.A. Sources of resistance in wheat to speckled leaf blotch caused by Septoria tritici. Euphytica, 1972, 21: 152-161.
- Saadaoui E.M. Physiologic specialization of Septoria tritici in Morocco. Plant Dis., 1987, 71(2): 153-165.
- Gough F.I. Effect of wheat host cultivars on pycnidiospore production by Septoria tritici. Phytopathology, 1978, 68(9): 1343-1345.
- Санина А.А. Физиологическая специализация Septoria tritici Rob. et Desm. Микология и фитопатология, 1991, 25(4): 338-342.
- Пыжикова Г.В., Санина А.А., Курахтанова Т.И., Давыдова Е.П., Породенко В.В., Санин С.С., Васецкая М.Н., Чигирев С.М., Дубынина Т.С., Москвитин Э.В., Катукова Н.П. Септориозы зерновых культур: метод. указ. М., 1988.
- Санина А.А., Анциферова Л.В. Способы выделения и хранения возбудителей септориоза пшеницы. Микология и фитопатология, 1989, 23(2): 172-175.
- Санина А.А., Анциферова Л.В. Определение патогенных свойств изолятов Septoria nodorum (Berk.) Berk. и S. tritici Rob. et Desm. на пшенице. Микология и фитопатология, 1991, 25(2): 155-159.
