Генетические эффекты в соматических клетках мышей, подвергнутых хроническому γ-облучению в малых дозах, и у их потомков

Автор: Раскоша О.В., Башлыкова Л.А.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 4 (28), 2016 года.
Бесплатный доступ
В экспериментах, проведенных на половозрелых мышах линии Af, установлены особенности влияния хронического γ-облучения в дозах 10, 20 и 30 сГр на степень поврежденности генома клеток костного мозга и щитовидной железы. Через четыре месяца после воздействия ионизирующего излучения выявлено изменение доли двунитевых разрывов ДНК в соматических клетках животных при частоте встречаемости клеток с микроядрами в обоих органах в пределах нормы. Более выраженные цитогенетические эффекты обнаружены в костном мозге по сравнению со щитовидной железой, в котором отмечена нелинейная зависимость проявления нестабильности генома от дозы облучения. Результаты, полученные на потомках (F1), подтверждают данные о наследуемости эффектов радиационно-индуцированной нестабильности генома.
Ионизирующее излучение, малые дозы, щитовидная железа, костный мозг, нестабильность генома
Короткий адрес: https://sciup.org/14992865
IDR: 14992865 | УДК: 575.155:575.113:599.323.4:539.166
Genetic effects in somatic cells of mice exposed to chronic low dose γ-irradiation, and their progeny
In order to reveal specific features of biological impact of low dose ionizing radiation, we studied genetic effects in bone marrow and thyroid gland cells in animals after exposure to low dose radiation, and also traced the possibility of the inheritance of radiation-induced changes in the progeny of animals of the first generation (F1). To assess genetic disorders in both organs, we used micronuclear test and DNA-comet approach (neutral version), we also determined mitotic index and the proportion of cells eliminated by apoptosis in the bone marrow. The study was performed using 160 adult mice of the Af line from the experimental animal nursery of the Institute of Biology, Komi Science Centre, Ural Branch, RAS. The animals were exposed to γ-irradiation during 29, 53 and 84 days using the dose of 150 μGy/h. The total absorbed dose was 10, 20 and 30 Gy consequently. The material for analyses was taken from F1 progeny, obtained from irradiated and control animals of 2-3 month age. As a result of molecular and cytogenetic analyses, the features of the effect of chronic γ-irradiation at used doses on the damage level of the genome of cells with different proliferative activity were revealed. Four months after the exposure to ionizing radiation changes were found in proportion of double-strand DNA breaks in somatic cells of animals at frequency of occurrence of cells with micronuclei in both organs being within normal. More pronounced cytogenetic effects were found in bone marrow in comparison with the thyroid gland, where there is marked nonlinear dependence of manifestation of genomic instability from the exposure dose. The results obtained from the progeny (F1) confirm the data on the heritability of the effects of radiation-induced gеnomic instability.
Текст научной статьи Генетические эффекты в соматических клетках мышей, подвергнутых хроническому γ-облучению в малых дозах, и у их потомков
Актуальность проблемы
Изучение нестабильности генома соматических клеток организма после действия ионизирующего излучения (ИИ) в дозах, реально встречающихся в природных условиях, а также исследование возможности наследования радиационно-индуцированных изменений представляется актуаль- ной проблемой современной радиобиологии. Полагают, что радиационно-индуцированная нестабильность генома является следствием неточной репарации повреждений структуры генома, так как облучение может менять его доступность для системы регуляции экспрессии генов [1]. Особенность этого эффекта проявляется в эпигенетическом характере наследования, т.е. в наследовании изме- ненного состояния генной экспрессии [2]. К настоящему времени накоплены данные, свидетельствующие о повышении нестабильности генома, в том числе и в соматических клетках, у потомства родителей, облученных в разных дозах [2–4]. Однако обнаруженная в области малых доз радиации нелинейность на кривых доза–эффект не позволяет получить законченного представления о закономерностях реакции биологических систем на воздействие факторов низкой интенсивности.
Для выявления особенностей биологического действия ИИ в малых дозах важным является изучение генетических эффектов в соматических клетках органов и тканей, в том числе обладающих разной пролиферативной активностью, таких как костный мозг и щитовидная железа. Костный мозг общепризнанно относится к наиболее радиочувствительным органам с быстрым темпом пролиферации, что позволяет даже при сравнительно небольшом облучении организма обнаруживать генетические нарушения в его клетках. Для определения эффектов после длительного воздействия ИИ перспективным представляется исследование ответной реакции щитовидной железы, состоящей преимущественно из медленно обновляющихся клеточных популяций, что, по данным [5], способствует накоплению структурных изменений ДНК после действия различных факторов.
Цель работы – изучить молекулярные и клеточные эффекты в костном мозге и щитовидной железе мышей линии Af, подвергнутых хроническому γ-облучению в диапазоне малых доз (10, 20 и 30 сГр), и проследить возможность наследования радиационно-индуцированных изменений в потомстве животных первого поколения (F 1 ).
Материал и методы
Эксперименты проводили на половозрелых мышах линии Af (n=96), полученных из питомника экспериментальных животных Института биологии Коми НЦ УрО РАН. Животные находились под воздействием ИИ в течение 29, 56 и 84 сут при мощности экспозиционной дозы 150 мкГр/ч (с кратковременными перерывами на кормление, смену поилок и уборку помещения), суммарные поглощенные дозы составили 10, 20 и 30 сГр, соответственно (опытные группы). Источником γ-излучения служили две ампулы, содержащие 0,474·106 и 0,451·106 кБк 226Ra, разнесенные на расстояние 2,5 м. Суммарную поглощённую дозу облучения определяли по показаниям термолюминесцентных дозиметров (ДТУ-01) с детекторами ДТГ-4 (LiF) на дозиметрической термолюминесцентной установке ДВГ–02ТМ (НПП "Доза", Россия). Для получения потомства (F1) во всех опытных группах (10, 20 и 30 сГр) сразу после облучения самцов спаривали с самками, семьи формировали из расчета один самец на одну/две самки, при этом учитывали их родословную, подбирая неродственных животных. Самцов через четыре месяца после облучения подвергали декапитации с последующим взятием материала для молекулярных и цитогенетических исследований клеток костного мозга и щитовидной железы. Одновременно с соответ- ствующими опытными группами в идентичные условия были посажены на размножение мыши контрольных групп. У потомков F1 (самцы, n=64), полученных от опытных и контрольных животных, материал для анализов брали у двух-трехмесячных особей (в каждой группе было пять–восемь зверьков).
Для цитогенетических исследований клеток костного мозга и щитовидной железы применяли микроядерный тест, который является информативным и быстрым способом индикации генотоксичности факторов внешней среды, в том числе и радиационной природы [6]. Для приготовления суспензии клеток костного мозга из бедренной кости с помощью фосфатно-солевого буфера (рН 7.4) вымывали костномозговую жидкость. Суспензию клеток щитовидной железы получали путем ферментативной диссоциации в 0,25%-ном растворе коллагеназы (Collagenase type IА, Sigma, USA). В зависимости от размеров ткани время инкубации составляло 40–60 мин. В суспензии клеток обоих органов добавляли гипотонический раствор (0,56% КCl, 20 мин, 37°С) и фиксировали охлажденной смесью метанола и ледяной уксусной кислоты (3:1), после чего клетки раскапывали на предметные стекла. Для окраски генетического материала использовали акридиновый оранжевый («Sigma»; 2 мкг/мл в фосфатно-солевом буфере). С помощью флуоресцентного микроскопа «Axioscop А1» ("Carl Zeiss", Jena) подсчитывали число клеток с микроядрами (МЯ) в обоих органах (1 тыс. клеток на препарат). В костном мозге на этих же препаратах анализировали митотический индекс (МИ) и долю клеток, элиминирующих по пути апоптоза [7]. Для подсчета разных типов клеток использовали лабораторный счетчик «СЛФ-ЭЦ-11-01М» (НПФ "Медтех", Россия).
Двунитевые разрывы ДНК (ДР ДНК) определяли по методу «ДНК-комет» в нейтральной версии рН, который основан на электрофорезе иммобилизованных в агарозу отдельных клеток [8]. Электрофорез осуществляли в трис-боратном буфере (рН 8.2, 4°C) при напряжении 1 В/см в течение 20 мин, с последующей фиксацией слайдов в 70° этаноле в течение 10 мин. «ДНК-кометы» визуализировали под микроскопом «Axioscop А1» с помощью видеосистемы на основе цифровой камеры «AxioCam» с программой «AxioVision SE64, Release 4.8.2» (“Carl Zeiss”, Германия) при флуоресцентном режиме, длина волны возбуждающего света – 450-490 нм, запирающий фильтр 510 нм. Для анализа фотоизображений «ДНК-комет» применяли программу «Со-met Score Pro» (“TriTek Corp.”, США), на слайде обрабатывали не менее 50–100 «комет» на каждое животное. Рассчитывали среднее значение %ТDNA, отражающее процент ДНК в «хвосте кометы»[8], вычисляли средние значения частоты клеток с неповрежденной и/или низко фрагментированной ДНК (практически не имеющие «хвоста кометы» или с небольшим «хвостом», диапазон 0-10%TDNA) и с высоко фрагментированной ДНК (имеющие более 40% ДНК в «хвосте кометы»). Достоверность различий между сравниваемыми группами оценивали по t-критерию Стьюдента и U-критерию Манна-Уитни.
Результаты и обсуждение
Результаты цитогенетического исследования костного мозга показали, что через четыре месяца после хронического γ-облучения мышей в диапазоне доз 10-30 сГр частота клеток с МЯ была в пределах значений, характерных для необлученных животных (рис. 1). В ответ на радиационное воздействие в этом органе происходило статистически значимое понижение относительно нормы элиминации клеток по пути апоптоза (во всех опытных группах) и уровня МИ (после доз 10 и 20 сГр). Известно, что замедление клеточного цикла (снижение МИ) может быть обусловлено остановкой клеточного цикла в точках контроля целостности ДНК, необходимой для увеличения продолжительности и эффективности репарации повреждений ДНК [9]. Сравнение ранжированных данных (см. таблицу) по процентному содержанию ДНК в «хвосте кометы» показало, что у мышей, подвергнутых радиационному воздействию в дозе 30 сГр, среднее значение %ТDNA было статистически значимо ниже нормы. По нашему мнению, на это повлияло увеличение в два раза (р<0,001) числа неповрежденных и/или малоповрежденных клеток, что может свидетельствовать об активации систем репарации ДНК в ответ на действие ионизирующей радиации в этой дозе. Средние значения %ТDNA у мышей, облученных в дозах 10 и 20 сГр, были в пределах нормы, однако ранжирование клеток по классам в зависимости от степени фрагментации ДНК в «хвосте кометы» позволило выявить различия с контролем и в этих опытных группах, причем направленность изменений зависела от дозы воздействия. Так, в костном мозге животных, облученных в дозе 10 сГр, по сравнению с контролем было снижено в 1,7 раза количество клеток с неповрежденной и/или низко фрагментированной ДНК (<10% ДНК в «хвосте кометы»; р<0,001), а после воздействия ИИ в дозе 20 сГр изменялась частота встречаемости клеток с высоко фрагментированной ДНК, когда более 40% ДНК находилось в «хвосте кометы» (понижение в 1,8 раза; р<0,01).
У потомков (F 1 ) облученных мышей по сравнению с потомками животных контрольной группы в клетках костного мозга выявлено статистически значимое повышение индукции клеток с МЯ, и наиболее ярко это проявлялось после доз 10 и 20 сГр (рис. 1). Данные литературы [4] и полученные нами результаты свидетельствуют о том, что увеличение частоты встречаемости МЯ в костном мозге является следствием повышенного уровня нестабильности генома соматических клеток потомков, облученных родителей. Кроме того, в костном мозге потомков, рожденных от облученных родителей, уменьшалась в 1,5–2,8 раза относительно нормы доля клеток с мало поврежденной ДНК (во всех
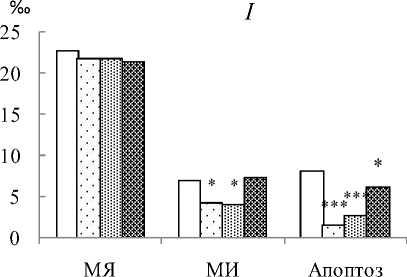
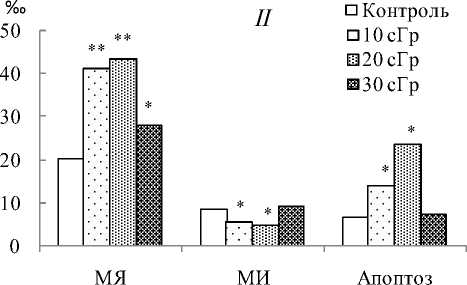
Рис. 1. Цитогенетические эффекты в клетках костного мозга мышей линии Af, подвергнутых хроническому γ -облучению в малых дозах ( I ), и у их необлученных потомков F1 ( II ).
Примечание. МЯ – микроядра, МИ – митотический индекс. Здесь и в таблице различия с контролем статистически значимы при *р<0.05, **р<0.01 и ***р<0.001.
Результаты метода ДНК-комет, проведенного на соматических клетках мышей линии Af, подвергнутых хроническому γ -облучению в малых дозах, и у их необлученных потомков (F 1 )
|
Группы животных |
Варианты эксперимента |
Костный мозг |
Щитовидная железа |
||||
|
%TDNA |
0-10% TDNA |
41-100% TDNA |
%TDNA |
0-10% TDNA |
41-100% TDNA |
||
|
Облученные животные |
Контроль |
23,3±0,3 |
13,1±0,8 |
5,7±0,6 |
31,7±0,96 |
18,6±1,1 |
33,3±1,3 |
|
10 сГр |
24,6±0,4 |
7,6±1,1*** |
4,0±0,8 |
31,9±0,97 |
19,3±1,9 |
34,7±2,3 |
|
|
20 сГр |
23,2±0,4 |
13,7±1,4 |
3,1±0,7** |
27,7±0,8* |
19,7±1,8 |
17,4±1,7*** |
|
|
30 сГр |
19,6±0,5*** |
27,9±1,8*** |
6,3±1,0 |
30,4±1,0 |
16,79±1,96 |
21,3±2,15*** |
|
|
Потомки облученных животных (F 1 ) |
Контроль |
15,6±0,6 |
40,2±2,6 |
5,3±1,2 |
15,8±0,8 |
42,6±2,9 |
5,2±1,3 |
|
10 сГр |
19,3±0,4*** |
23,8±1,3*** |
3,4±0,6 |
15,8±0,7 |
50,7±1,98* |
5,35±0,9 |
|
|
20 сГр |
24,7±0,8*** |
26,6±1,3*** |
12,2±1,0*** |
18,2±0,8* |
42,6±2,2 |
7,6±1,2 |
|
|
30 сГр |
25,6±0,5*** |
14,2±1,2*** |
12,9±1,2*** |
15,78±0,7 |
46,6±2,1 |
4,8±0,9 |
|
Примечание: %TDNA – среднее значение ДНК в «хвосте кометы»; 0-10%TDNA – диапазон встречаемости клеток с низко фрагментированной ДНК, 41-100%TDNA – диапазон встречаемости клеток с высоко фрагментированной ДНК.
опытных группах) и увеличивалось в 2,4 раза количество клеток с фрагментацией ДНК более 40% (родители были облучены в дозах 20 и 30 сГр) (см. таблицу). В результате средние значения содержания ДНК в «хвосте комет» в костном мозге потомков всех опытных групп были статистически значимо выше контроля (р<0,001). По данным литературы [4, 10], повреждения ДНК, с одной стороны, являются субстратом для репаративных ферментов в клетке, а с другой – запускают программу клеточной гибели – апоптоз. В условиях проведенного эксперимента воздействие ИИ на мышей в дозах 10 и 20 сГр вызывало в костном мозге у их потомков статистически значимое повышение частоты встречаемости апоп-тозных клеток и уменьшение доли МИ, чего не наблюдали у животных в случае, если их родителей подвергали радиационному воздействию в большей дозе (30 сГр). Изменение пролиферативной активности и элиминации клеток под воздействием малых доз ИИ может свидетельствовать о перестройке работы системы защиты клеток, которая к тому же передается следующему поколению животных. Полагаем, что изменения МИ и количества апоптозных клеток в костном мозге потомков животных, облученных в дозах 10 и 20 сГр, являются ответной реакцией на повышенный уровень повреждений ДНК и свидетельствуют об изменении программы поддержания стабильности генома не только через активацию процессов репарации ДНК, но и апоптоза.
Данные микроядерного теста, проведенного на щитовидной железе мышей, показали, что после хронического воздействия ИИ в разных дозах число клеток с МЯ в исследуемой ткани, как и в костном мозге, сохранялось в пределах значений, соответствующих контролю (рис. 2). Применение метода «ДНК-комет» позволило выявить изменения на молекулярном уровне в геноме тироцитов животных, облученных в дозах 20 и 30 сГр: после радиационного воздействия в этих дозах число клеток с высоким содержанием разрывов нитей ДНК (диапазон 41–100%TDNA) было ниже нормы в 2 и 1,6 раза, соответственно (см. таблицу). Это повлияло на средние значения содержания ДНК в «хвосте кометы» (20 сГр; р≤0,01). О понижении доли тироцитов с ДР ДНК после облучения животных в дозе 30 сГр свидетельствуют результаты, описанные нами ранее в работе [11], где использовали интегральный показатель «Olive tail moment», учитывающий и длину
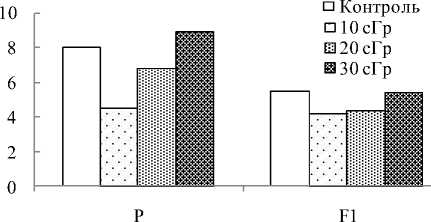
Рис. 2. Частота клеток с МЯ в щитовидной железе мышей линии Af, подвергнутых хроническому γ -об-лучению в малых дозах, и у их необлученных потомков F1.
«хвоста кометы», и плотность содержания ДНК в «хвосте». Наряду с этим выявлено, что после воздействия ИИ в максимальной дозе была значительно повышена частота встречаемости клеток на стадии позднего апоптоза, когда более 90% ДНК выходило за пределы «головы кометы» [7]. Малые дозы облучения (20 и 30 сГр) могли явиться адаптирующим фактором и вызвать усиление репарации разрывов нитей ДНК в клетках щитовидной железы, которое в конечном итоге привело по сравнению с нормой к понижению числа поврежденных клеток и усилению апоптоза (30 сГр). Следует отметить, что в условиях проведенного эксперимента в щитовидной железе мышей, подвергнутых воздействию ИИ в дозах 10-30 сГр, число клеток, в которых ДНК находилось вблизи «головы кометы», было в пределах значений, характерных для необлученных животных.
При сравнительном цитогенетическом анализе клеток щитовидной железы потомков мышей, полученных от контрольных и облученных (разными дозами) родителей, не обнаружено статистически значимых отличий по частоте встречаемости клеток с МЯ. Тем не менее, при изучении фрагментации ДНК в тироцитах потомков выявлены некоторые особенности, проявляющиеся в зависимости от используемой при экспонировании их родителей дозы. В связи с этим заслуживают внимания данные о повышении доли неповрежденных клеток (диапазон 0-10%TDNA; р≤0,05) в щитовидной железе потомков мышей, облученных в дозе 10 сГр, а с увеличением дозы облучения животных до 20 сГр, наоборот, у их потомков происходило удлинение «хвоста кометы». Это может свидетельствовать о повышении ДР ДНК в тироцитах мышей данной опытной группы.
Заключение
Таким образом, можно сделать вывод об эффективности хронического воздействия ИИ в малых дозах (10-30 сГр) на костный мозг и щитовидную железу – органы, обладающие разной пролиферативной активностью. Применение цитогенетического и молекулярного методов показало, что через четыре месяца после хронического воздействия ИИ в малых дозах отмечено изменение доли ДР ДНК в клетках костного мозга и щитовидной железы, при этом частота встречаемости клеток с МЯ после облучения мышей в обоих органах сохранялась в пределах нормы. В ответ на малые дозы ИИ более выраженные эффекты на клеточном и молекулярном уровнях установлены в костном мозге по сравнению со щитовидной железой. Важным является выявленная в костном мозге нелинейная зависимость проявления нестабильности генома от дозы облучения, когда большая эффективность наблюдалась после низких доз радиационного воздействия (10 и 20 сГр по сравнению с 30 сГр). Результаты, полученные на потомках F 1 , подтверждают данные о наследуемости эффектов радиационно-индуцированной нестабильности генома. Обнаруженные в потомстве нарушения проявлялись как и у их родителей, преимущественно в клетках костного мозга с наследованием нелинейного характера зависимости доза–эффект.
Работа поддержана из средств темы НИР «Сохранение коллекций экспериментальных животных для фундаментальных исследований».
Список литературы Генетические эффекты в соматических клетках мышей, подвергнутых хроническому γ-облучению в малых дозах, и у их потомков
- Заварыкина Т.М. Структурные изменения ДНК при действии низкоинтенсивной ионизирующей радиации в малых дозах: Автореф. дис. канд. мед. наук. М., 2008. 26 с
- Дуброва Ю.Е. Нестабильность генома среди потомков, облученных родителей. Факты и интерпретация//Генетика. 2006. Т. 42. №10. С. 1335-1347
- Воробцова И.Е. Трансгенерационная передача радиационно-индуцированной нестабильности генома//Радиационная биология. Радиоэкология. 2006. Т.46. №4. С. 441-446
- Кузнецова E.А., Заичкина С.И., Сирота Н.П. и др. Индукция редко-и плотноионизирующими излучениями повреждений ДНКвлейкоцитах крови и цитогенетических повреждений в полихроматофильных эритроцитах костного мозга мышей и их потомков//Радиационная биология. Радиоэкология. 2014. Т. 54. № 4. С. 341-349
- Гансбургский М.А. Анализ клеток с микроядрами в оценке пролиферации эпителия щитовидной железы: Автореф. дис. канд. мед. наук. М., 2005. 21 с
- Полиорганный микроядерный тест в эколого-гигиенических исследованиях/Под ред. Ю.А. Рахманина, Л.П. Сычевой. М.: Гениус, 2007. 312 с
- Тронов В.А. Репарация ДНКиапоптоз//Цитология. 1999. Т. 41. № 5. С. 405-410
- Olive P.L., Banath J.P. Detection of DNA double-strand breaks through the cell cycle after exposure to X-rays, bleomycin, etoposide and 125IdUrd//Int. J. Radiat. Biol. 1993. Vol. 64. № 4. Р. 349-358
- Жижина Г.П. Влияние малых доз низкоинтенсивной ионизирующей радиации на структуру и функции ДНК//Радиац. биология. Радиоэкология. 2011. Т. 51. № 2. С. 218-228
- Oliver F.J., Menissierde Murcia J., and de Murcia G. Poly(ADP-Ribose)Polymerase in the Cellular Response to DNA Damage, Apoptosis and Disease//Am. J. Hum. Genet. 1999. №64. Р. 1282-1288
- Раскоша О.В. Оценка стабильности генома в клетках щитовидной железы мышей после хронического воздействия ионизирующего излучения в малой дозе, и у их потомков//Успехи современного естествознания. 2016. № 4. С. 48-52
