Генетический анализ наследования высоты растений риса, длины метелки, числа и массы колосков в ней

Автор: Костылев Павел Иванович, Попов Сергей Сергеевич
Журнал: Вестник аграрной науки Дона @don-agrarian-science
Рубрика: Агрономия, лесное хозяйство и биологические науки
Статья в выпуске: 2 (22), 2013 года.
Бесплатный доступ
Рассмотрены генетические различия между родительскими формами гибридов второго поколения риса по количественным признакам, тесно связанным с урожайностью.
Рис, наследование, масса и длина метелки, длина стебля, количество колосков, гибридные популяции
Короткий адрес: https://sciup.org/140204229
IDR: 140204229 | УДК: 575.1:633.18
Genetic analysis of rice heights inheritance, length of panicle rice, quantity and mass of cones in it
Genetic distinctions between parental forms of the rice second generation hybrids on the quantitative signs which have been closely connected with productivity are considered.
Текст научной статьи Генетический анализ наследования высоты растений риса, длины метелки, числа и массы колосков в ней
Продуктивность сельскохозяйственных культур - это количественный признак, на который большое влияние оказывают другие количественные признаки. Оптимальное их сочетание позволяет получать наибольшие урожаи с единицы площади. По высоте растений предпочтение отдается низкорослым сортам риса, которые устойчивы к полеганию. Низкорослость растений может быть одним из самых важных агрономических признаков, потому что часто сопровождается устойчивостью к полеганию и таким образом можно применять большие дозы удобрений. Полукарликовость контролируется преимущественно одним мутантным геном с неполным доминированием. Она может быть обусловлена неполным доминированием высокорослости, варьированием числа модификаторов, взаимодействием генов полукарликовости sd с другими генами (Chang Т„ 1964).
Длина метелки и ее плотность имеют большое значение для формирования зерновой продуктивности растений. Исследования ряда мутантов открыли некоторое количество генов, участвующих в регулировании формирования метелки. У мутантов метелки ген lax 1 ограничено инициирует развитие веточек, боковых и терминальных колосков, указывая на то, что этот ген необходим для закладки пазушных меристем в метелке риса (Komatsu М. и др., 2001). Маленькая метелка (small panicle spa^ является другим главным регулятором формирования пазушной меристемы (Komatsu К. и др., 2003). Подобно lax мутантам, количество ветвей метелки и колосков у spa мутантов сильно уменьшено.
В дополнение к генам, управляющим закладкой меристемы, гены, влияющие на быстрое увеличение клеток, которое в свою очередь влияет на размер меристемы, в конечном счете, регулируют скорость
Вестник аграрной науки Дона дифференциации колосков, важную для размера метелки и числа колосков (Ikeda К. и др., 2005).
Согласно концепции апикального доминирования, отношения между ростом побега и ветвлением регулируются балансом между ауксином, который подавляет рост пазушных почек, и цитокинином, который снижает подавление. Такой баланс фитогормонов также имеет место при регулировании ветвления метелки. Молекулярное клонирование и анализ QTL для числа зерен (grain number 1 Gnla^ показали роль цитокина в регулировании размера метелки (Ashikari М. и др., 2005). Gnla кодирует фермент (OsCKX2), который понижает цитокинин. Когда проявление OsCKX2 снижено, цитокинин накапливается в меристемах соцветия и увеличивает число репродуктивных органов, которые увеличивают число зерен и приводят к увеличению сбора зерна, не влияя на фенологию растения риса. Различие в 34 зерна на главной метелке было отмечено между двумя линиями, гомозиготными по двум аллелям Gn 1 а. Наоборот, мутант lonely guy QogY У которого есть дефект в синтезе активного цитокинина, дает метелку намного меньшую, чем дикий тип (Kurakawa Т. и др., 2007). Таким образом, цитокинины необходимы в контроле формы метелки у риса.
Для селекционной модели сорта важно знать оптимальные величины этих признаков. Для этого необходимо изучить наследование и экспрессию генов, возможность комбинирования со скороспелостью, оптимальной высотой растений и др.
Цель и задачи исследований - определение типа наследования, числа и силы генов, участвующих в проявлении количественных признаков высоты растений, длины метелки, числа и массы колосков на ней у гибридов от скрещивания образцов риса Ил. I4 и Ил. 28 с сортом Кубояр.
Исходный материли - среднерослый сорт Кубояр, высотой 90-100 см, с компактной прямостоячей метелкой длиной 12,5 см, полученый от скрещивания сортов Кубань и Боярин. Ил. 14 - низкорослый образец высотой 50-60 см, несущий гены устойчивости к пирикуляриозу и Ил. 28 -высокорослый образец высотой 120-130 см, с поникающей метелкой, также с генами устойчивости к пирикуляриозу. Анализировали по 500 гибридных растений F2 от каждой комбинации скрещиваний и по 150 родительских растений.
Для генетического анализа использовали методики А С. Серебровского (1981) и компьютерную программу поиска моделей расщепления Полиген А (Мережко А.Ф., 1984). Исследования проводили на полях ФГУП «Пролетарское» Ростовской области.
Результаты исследований. Установлено, что высота растений и длина метелки взаимосвязаны. Коэффициент корреляции между этими признаками (0,65±0,07) в расщепляющихся гибридных популяции F2 был средним положительным.
Распределение по высоте в обеих комбинациях имело незначительную правостороннюю асимметрию (As=-0,15), что свидетельствует об отрицательном доминировании (рис. 1).
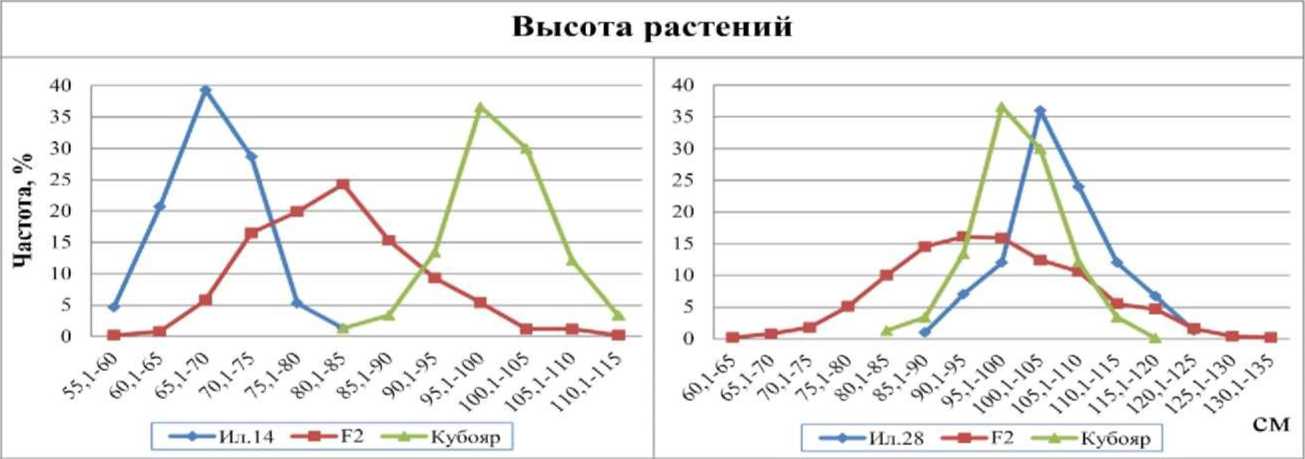
Рис. 1. Распределение частот признака «высота растений» у гибридов риса F2 Ил. 14 х Кубояр, Ил. 28 х Кубояр и их родительских форм, 2012 г.
В комбинации Ил. 14 х Кубояр родительские формы значительно различались между собой. Кубояр - среднерослый сорт с высотой 100 см, а Ил. 14 - низкорослый с высотой 68,6 см. Кривая распределения частот гибрида находилась в пределах распределения обеих родительских форм, а её вершина оказалась примерно посередине между ними, что говорит о промежуточном наследовании признака. На долю рецессивного родителя приходилась 1/16 часть частот распределения гибрида, что свидетельствует о различии родительских форм по аллельному состоянию двух пар генов, детерминирующих длину стебля. Расщепление происходило в соотношении 9:6:1. При этом наблюдалось частичное отрицательное доминирование (һр=-0,5). Средняя сила одного аллеля, удлиняющего стебель - 15,7 см.
В комбинации Ил. 28 х Кубояр родительская форма Ил. 28 немного, в среднем на 2,5 см выше, чем Кубояр. Среди гибридов выщепилось небольшое количество трансгрессивных форм, что свидетельствует о различном аллельном состоянии генов. Родительские формы различались по аллельному состоянию двух пар генов.
По длине метелки родительские формы в комбинации Ил. 14 х Кубояр практически не различались, что говорит об отсутствии различий по генам между ними. В комбинации Ил. 28 х Кубояр различия между родителями существенны. У Кубояра средняя длина метелки - 13,9 см, у Ил. 28 - 21,2 см. Гибрид имел промежуточную длину метелки 17 см. Кривая распределения частот признака гибрида находилась в пределах изменчивости родительских форм, асимметрия почти отсутствовала (As=0,15), а вершина гибрида находилась посередине между вершинами родительских форм, что свидетельствовало об отсутствии доминирования (рис. 2). На долю рецессивного родителя приходилась 1/16 часть частот гибрида, что указывает на аллельные различия по двум парам генов. Сила одного аллеля составила 3,6 см.
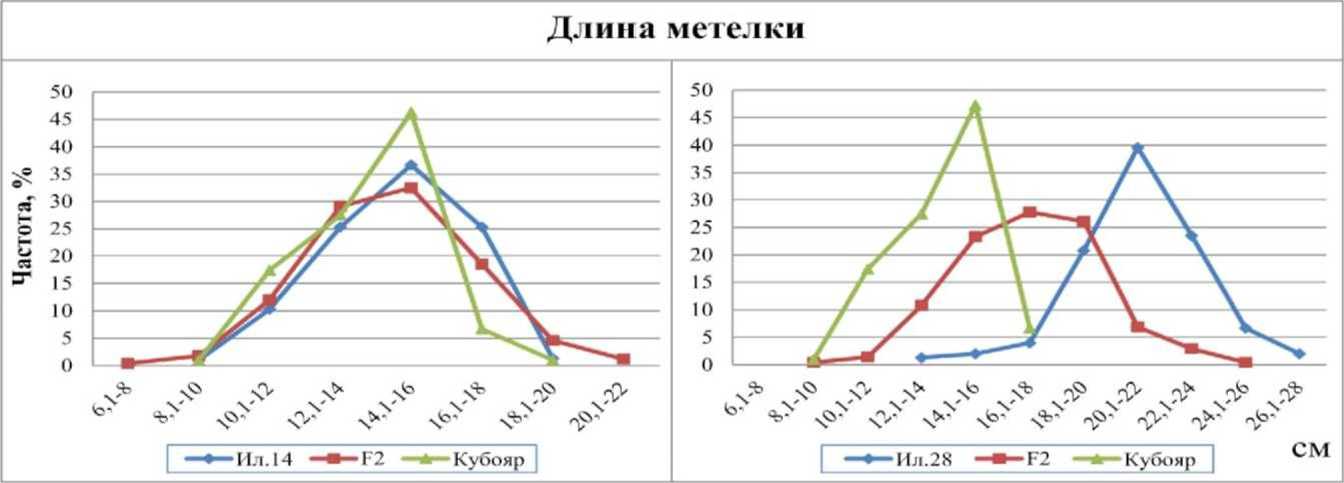
Рис. 2. Распределение частот признака «длина метелки» у гибридов риса F2 Ил. 14 х Кубояр, Ил. 28 х Кубояр и их родительских форм, 2012 г.
Изменчивость по признаку «число колосков в метелке» очень высока и варьирует от 40 до 380 колосков у гибридов до 220 у Кубояра и до 300 у Ил. 14 и Ил. 28. В комбинации Ил. 14 х Кубояр кривые распределения частот родителей и гибрида идентичны, что свидетельствует об отсутствии различий между ними.
По числу колосков на метелке Ил. 28 (154 шт.) и Кубояр (114 шт.) различались на 40 штук. У гибрида значение этого признака составило 144 колоска, приближая его к Ил. 28, тем самым свидетельствуя о частичном доминировании большего значения (hp=0,46). Выщепилось большое количество трансгрессивных форм (рис. 3). На долю рецессивного родителя приходилась 1/4 часть частот гибрида, что указывает на аллельные различия по одной паре генов. Сила гена составила 40 колосков.
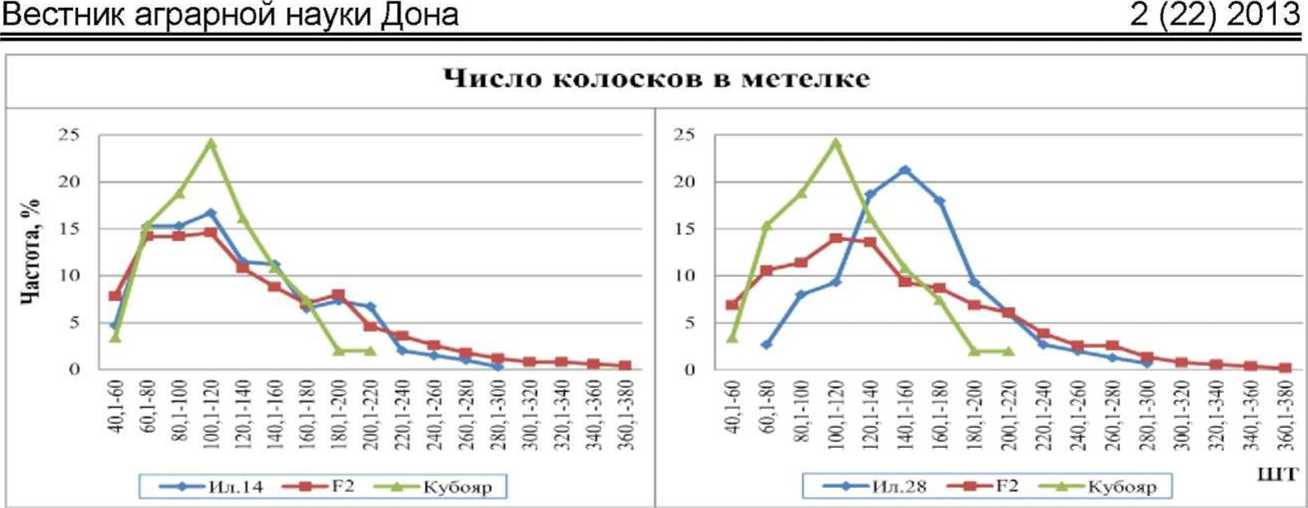
Рис. 3. Распределение частот признака «число колосков в метелке» у гибридов риса Ғ2 Ил. 14 х Кубояр, Ил. 28 х Кубояр и их родительских форм, 2012 г.
Масса зерна с метелки в значительной степени связана с числом колосков в ней (г=0,8-0,9), поэтому графики распределения частот этого признака были похожи на предыдущие. В комбинации Ил. 14 х Кубояр родительские формы отличались друг от друга на 1,1 г. Кривая распределения частот признака гибрида была асимметричной (As=l,04), а среднее значение гибрида находилось между родительскими формами, что свидетельствовало о промежуточном наследовании признака (рис. 4). На долю обеих родительских форм приходилось по 1/4 частот гибрида, что указывает на аллельные различия по одной паре генов, которые расщеплялись в соотношении 1:2:1. Сила действия гена составила 0,55 г.
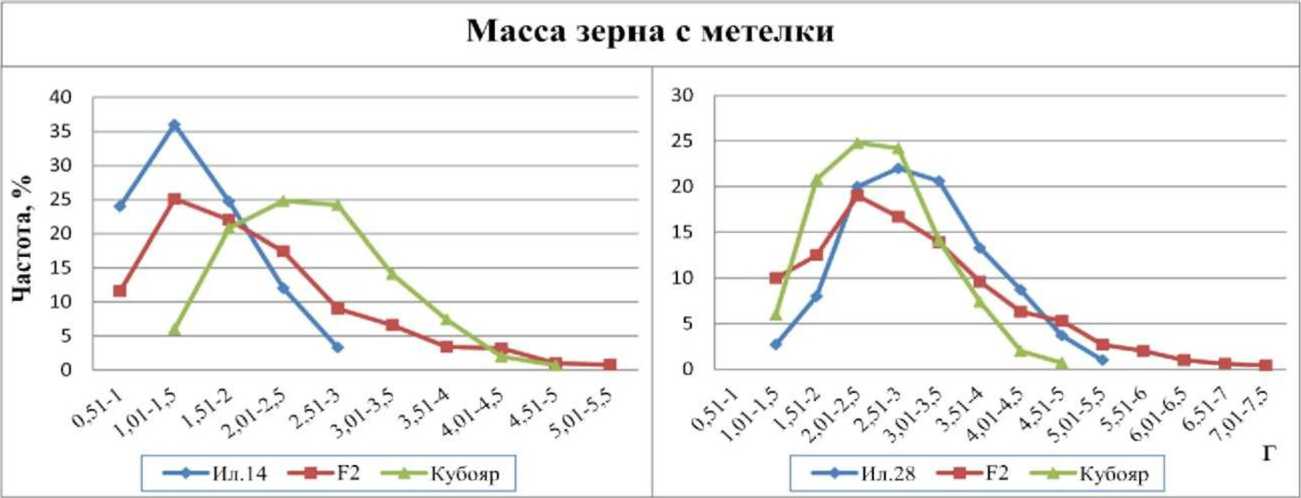
Рис. 4. Распределение частот признака «масса зерна с метелки» у гибридов риса F2 Ил. 14 х Кубояр, Ил. 28 х Кубояр и их родительских форм, 2010 г.
В комбинации Ил. 28 х Кубояр родители незначительно отличались друг от друга, в среднем на 0,47 г. Среднее значение массы зерна с метелки у гибрида находилось между родителями, что свидетельствует о промежуточном наследовании признака. Родительские формы различались по аллельному состоянию одного гена.
Небольшие различия исходных форм по числу генов, детерминирующих длину стеблей и метелок, дают возможность с малыми затратами отбирать новые формы риса с нужным сочетанием признаков.
Выделены формы F2, сочетающие низкорослость растений с длинной, крупной метелкой, несущей много зерен, кото- рые представляют интерес для дальнейшей селекционной работы.
Выводы
-
1. По высоте растений образцы Ил. 14 и Кубояр различались по двум парам генов со средней силой аллеля 15,7 см. Расщепление происходило в соотношении 9:6:1, имелось отрицательное доминирование һр—0,5. Образцы Ил. 28 и Кубояр также различались по двум парам генов.
-
2. Образцы Ил. 28 и Кубояр различались по аллельному состоянию двух пар генов, сила действия которых в сумме составила 7,2 см. Не выявлены различия по длине метелки у образцов Ил. 14 и Кубояр.
-
3. По числу колосков между образцами Ил. 14 и Кубояр нет различий в генах. Ил. 28 отличался от Кубояра одной аллельной парой генов, сила действия которой составила 40 колосков. Имелось частичное положительное доминирование һр=0,46.
-
4. По массе зерна с метелки родительские формы обеих комбинаций гибридов различались аллельным состоянием одной пары генов.
Список литературы Генетический анализ наследования высоты растений риса, длины метелки, числа и массы колосков в ней
- Мережко, А.Ф, Система генетического изучения исходного материала для селекции растений/А.Ф. Мережко. -Ленинград: ВИР, 1984. -70 с.
- Серебровский, A.C. Генетический анализ/A.C. Серебровский. -Москва: Наука, 1981.-342 с.
- Ashikari М., Sakakibara H., Lin S.Y., Yamamoto T., Takashi T. et al. Cytokinin ox idase regulates rice grain production. Science, 2005. -309. -P. 741-45.
- Chang T. Present knowledge of rice genetics and cytogenetics//Tehnical Bullein IRRI -1964. -№ l.-P. 1-94.
- lkeda K., Nagasawa N., Nagato Y. Aberrant panicle organizati on 1 ternporally regulates meristem identity in rice. Dev. Biol., 2005. -282. -P. 349-60.
- Komatsu M., Maekawa M., Shimamoto K., Kyozuka J. The LAX1 and FRIZZY PANICLE 2 genes determine the inflorescence architecture of rice by controlling rachis branch and spikelet development. Dev. Biol., 2001.-231. -P. 364-373.
- Komatsu K., Maekawa M., Ujiie S., Satake Y., Furutani F et al. LAX and SPA: major regulators of shoot branching in rice. Proc. Natl. Acad. Sei. USA, 2003. -100. -P. 11765-11770.
- Kurakawa T., Ueda N., Maekawa M., Kobayashi K., Kojima M. et al. Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature, 2007. -445. -P.652-655.
- Li S., Qian Q., Fu Z., Zeng D., Meng X. et al. Short paniclel encodes a putative PTR family transporter and determines rice panicle size. Plant J., 2009. -58. -P. 592-605.
- Xue W., Xing Y., Weng X., Zhao Y., Tang W. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet., 2008. -40. -P. 761-767.
