Генетическое разнообразие естественных популяций сосны обыкновенной в северной части Республики Карелии

Автор: Мельчакова Е.В., Малеева Н.А.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Материалы 7-го международного совещания "Сохранение лесных генетических ресурсов"
Статья в выпуске: 1, 2023 года.
Бесплатный доступ
Проанализировано генетическое разнообразие и структура 5 популяций сосны обыкновенной (Pinus sylvestris L.) из северной части Республики Карелии с использованием 13 микросателлитных локусов. Установлено высокое аллельное разнообразие популяции сосны обыкновенной на исследованных участках, несмотря на то что по степени равномерности распределения аллельных вариантов внутри выборок все исследуемые популяции достаточно однородны. Результат AMOVA-анализа показал относительно невысокий уровень межпопуляционной дифференциации: внутрипопуляционная изменчивость составляет 93 %, а межпопуляционная - 7 %. По результатам кластерного анализа, проведённого на основе матрицы генетических расстояний М. Неи, установлена популяционная структура P. sylvestris в северной части Карелии. Все изученные популяции подразделены на две большие группы.
Pinus sylvestris l, естественные популяции, генетическая структура, пцр, микросателлиты, генетическое разнообразие, est-ssr
Короткий адрес: https://sciup.org/143179856
IDR: 143179856 | УДК: 575.174.015.3 | DOI: 10.24419/LHI.2304-3083.2023.1.07
Genetic diversity of natural populations of scotch pine in the northern part of the Republic of Karelia
In this study, we assessed the genetic diversity and population structure of Scotch pine (Pinus sylvestris L.) using 13 microsatellite loci in 5 populations from the northern part of the Republic of Karelia. Our analysis showed a high allelic diversity of the Scots pine population in the studied areas. Although, in terms of the degree of uniformity of the distribution of allelic variants within the samples, all the studied populations are quite homogeneous. The result of the AMOVA analysis showed a relatively low level of interpopulation differentiation, within the population the variability is 93 %, and the interpopulation share of variability is 7 %. According to the results of cluster analysis carried out on the basis of the matrix of genetic distances by M. Nei, we revealed the population structure of P. sylvestris in the northern part of Karelia. All studied populations were divided into two large groups.
Текст научной статьи Генетическое разнообразие естественных популяций сосны обыкновенной в северной части Республики Карелии
Сосна обыкновенная ( Pinus sylvestris L.) – одна из наиболее распространённых и значимых хвойных пород Республики Карелии. Экологическое и экономическое значение этого вида стимулировало обширные исследования в отношении его происхождения, а также множество исследовательских проектов по морфологии и генетике. Однако эксплуатация лесов Карелии, наиболее активно происходившая в начале 1930-х гг. [1], привела к сильной трансформации и фрагментации коренных сосновых лесов в регионе [2]. Это не могло не отразиться на генетическом разнообразии и внутрипо-пуляционной дифференциации современной популяции сосны обыкновенной в исследуемом регионе.
Естественные популяции сосны обыкновенной длительное время изучали с помощью изо-ферментного анализа, как правило, в пределах отдельных территорий. Проанализированные изоферментные системы имеют внутрипопу-ляционную изменчивость менее 8 % [3–5]. Использование ДНК-маркеров в течение последних 10–15 лет привело к более глубокому пониманию структуры популяции и истории распространения сосны [6–9].
В настоящее время при изучении генетического разнообразия хвойных пород широко применяют EST-SSR-маркеры, поскольку они имеют двуродительское наследование, многочисленны в геноме сосны, кодоминантны и вариабельны [10]. Проводившиеся ранее молекулярно-генетические исследования в Карелии показали отсутствие какой-либо закономерности в изменчивости частот аллелей в направлении север–юг и мозаичный характер аллельного разнообразия, не выявлено четкой закономерности в распределении генетического разнообразия карельских популяций сосны обыкновенной [2].
Наше исследование дополняет предыдущие работы по изучению генетического разнообразия и популяционной структуры сосны обыкновенной с помощью ядерных микросателлитных локусов.
Цель исследования – оценка генетического разнообразия и структуры естественных популяций сосны обыкновенной в северной части Республики Карелии.
Материалы и методы
Исследования проведены в 5-ти естественных популяциях сосны обыкновенной, расположенных в северной части Республики Карелии (табл. 1, рис. 1):
-
V близлежащий населенный пункт Чупа;
-
V сельский населенный пункт Амбарнское;
-
V поселок Муезерский;
-
V поселок Юшкозеро;
-
V поселок Кевятозеро.
Точки сбора образцов обусловлены разными условиями произрастания и достаточной удалённостью друг от друга (не менее 50 км), чтобы охватить разнообразные по микроклиматическим
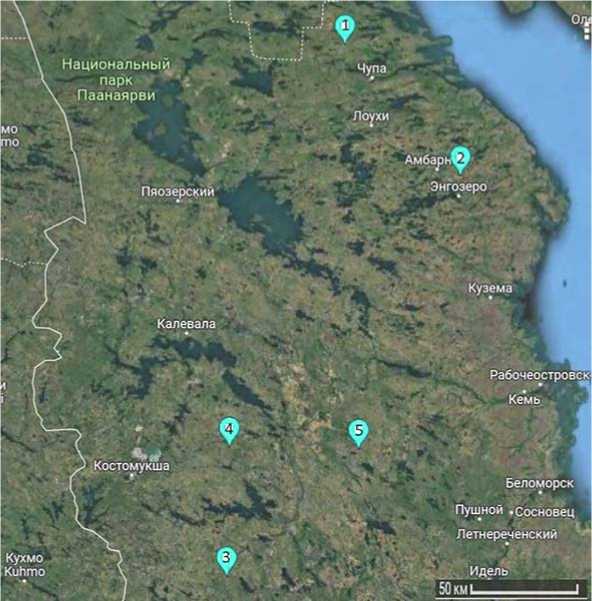
Рис. 1. Карта-схема мест сбора образцов сосны обыкновенной:
1 – Чупа, 2 – Амбарнское, 3 – Муезерский, 4 – Юшкозеро, 5 – Кевятозеро
Таблица 1. Характеристика точек отбора проб сосны обыкновенной карельских популяций
Выделение образцов геномной ДНК сосны осуществлялось с помощью набора GenElute Plant Genomic DNA Miniprep Kit (Sigma). Для анализа генетического разнообразия популяций сосны обыкновенной было отобрано 13 ядерных микро-сателлитных локусов (табл. 2).
Протоколы амплификации взяты из оригинальных источников [11–15]. Амплификацию осуществляли на термоциклире T100TM (BioRad, США). Для проведения полимеразной цепной реакции (ПЦР) использовали набор ScreenMix-HS (ЗАО Евроген, Россия). Визуализация продуктов амплификации проводилась с помощью вертикального электрофореза в 6 %-м полиакриламидном геле с последующей окраской фрагментов ДНК красителем NonTox (AppliChem, Германия). В качестве стандартного маркера длин фрагментов использована ДНК плазмиды pBR322 (Сибэн-зим, Россия), обработанная рестриктазой HPAII (Сибэнзим, Россия). Генотипирование проводили в программе Photo-Capt 12.4 (Vilber Lourmat, Франция).
Основные показатели генетической изменчивости: среднее число аллелей на локус (Na), среднее эффективное число аллелей (Ne), наблюдаемая (Но) и ожидаемая (Не) гетерозиготность, коэффициент фиксации Райта F, критерий χ2 для оценки соответствий наблюдаемых и ожидаемых распределений генотипов по правилу Харди–Вайнберга, анализ молекулярной дисперсии (АMOVA – Analysis of Molecular Variance), показатели
Таблица 2. Характеристики микросателлитных праймеров для сосны обыкновенной
|
Локус (праймер) |
Мотив |
Ожидаемые размеры ампликона, п.н. |
Температура отжига (Ta), °C |
Литературный источник |
|
lw_isotig21953 |
(ATGGG)7 |
208 |
60 |
Fang et al.,2014 [12] |
|
lw_isotig04306 |
(TCC)7 |
196 |
55 |
Fang et al., 2014 [12] |
|
lw_isotig04195 |
(GAG)5 |
189 |
55 |
Fang et al., 2014 [12] |
|
lw_isotig27940 |
(TGGA)5 |
231 |
55 |
Fang et al., 2014 [12] |
|
PtTx 2123 |
(AGC) 8 |
192–207b |
60–50 ↓ |
Elsik et al., 2000 [11] |
|
PtTx 2146 |
(GAG)5… (CAG)8CGG(CAG)7CGG(CAG)4 |
168–249 |
60–50 ↓ |
Elsik et al., 2000 [11] |
|
PtTx 3107 |
(CAT) 14 |
150–182 |
55–45 ↓ 1.0 °C |
Liewlaksaneeyanawin et al., 2004 [14] |
|
PtTx 3116 |
(TTG)7(TTG)5 |
122–226 |
55 |
Liewlaksaneeyanawin et al., 2004 [14] |
|
PtTx 4001 |
(CA) 15 |
201–224 |
60–50 ↓ 1.0 °C |
Gonzalez-Martinez et al., 2004 [13] |
|
Spag 7.14 |
(TG)17(AG)21 |
187–211 |
55–50 ↓ 1.0 °C |
Gonzalez-Martinez et al., 2004 [13] |
|
psy117 |
(TA)7 |
219–251 |
55 |
Sebastiani et al., 2012 [15] |
|
psy142 |
(TC)9 |
171–179 |
55 |
Sebastiani et al., 2012 [15] |
|
psy144 |
(CGG)5 |
166–175 |
55 |
Sebastiani et al., 2012 [15] |
F-статистики Райта и генетические дистанции по Нею определяли с помощью программы GenAlEx 6.5 [16]. Оценку степени однородности распределения аллельных вариантов внутри выборки проводили с помощью индекса Шеннона (I) [17]. Построение дендрограммы на основе матрицы генетических расстояний осуществляли с помощью метода невзвешенного попарного арифметического среднего UPGMA [18]. Для обнаружения и исключения возможных вариантов ошибок генотипирования ядерных микросателлитных локусов использована программа Micro-Checker [19].
Результаты и обсуждение
Анализ генетической структуры карельских популяций сосны обыкновенной показал, что все использованные микросателлитные локусы оказались полиморфны (Р99 % = 100 %).
Всего выявлено 114 аллельных вариантов, из которых 18 % являются уникальными. Наибольшее аллельное разнообразие обнаружено в популяции Кевятозеро, которое в среднем составило
12 аллельных вариантов на локус (табл. 3). Наименьшее количество зафиксировано в популяциях Амбарнское и Чупа, которое составило по 8 аллелей на локус. Стоит отметить, что данные популяции находятся в самых северных точках исследования – 65° и 66° с. ш. соответственно. В целом по всем изученным выборкам аллельное разнообразие составляет 10±0,565 аллелей на локус, что показывает высокое аллельное разнообразие популяции сосны обыкновенной на исследованных участках (см. табл. 3).
Эффективное число аллелей (Ne) варьирует от 4 до 7 на локус в среднем для всех рассматриваемых популяций. Наибольшие показатели эффективного числа аллелей отмечены для локусов PtTx 4001, PtTx2146, PtTx2146, lw_isotig21953, которые являются наиболее характерными по частоте встречаемости.
По степени равномерности распределения аллельных вариантов внутри выборок все исследуемые популяции достаточно однородны: индекс Шеннона варьирует от 1,672 до 1,884, за исключением популяций Кевятозеро и Юшкозе-ро, для которых индекс Шенона составляет 2,096 и 1,990 соответственно. Это указывает на более
Таблица 3. Уровень генетического разнообразия в популяциях P. sylvestris в северной части Карелии
|
Популяция |
Локус |
Na Ne |
I Ho |
He |
F |
||
|
Кевятозеро |
lw_isotig21953 |
15,000 |
9,164 |
2,405 |
0,204 |
0,891 |
0,771 |
|
lw_isotig04306 |
10,000 |
5,532 |
1,899 |
0,688 |
0,819 |
0,161 |
|
|
lw_isotig04195 |
5,000 |
3,441 |
1,416 |
0,458 |
0,709 |
0,354 |
|
|
lw_isotig27940 |
15,000 |
8,711 |
2,376 |
0,220 |
0,885 |
0,751 |
|
|
PtTx 3107 |
12,000 |
7,042 |
2,153 |
0,543 |
0,858 |
0,367 |
|
|
PtTx 3116 |
9,000 |
4,439 |
1,744 |
0,097 |
0,775 |
0,875 |
|
|
PtTx 4001 |
21,000 |
12,285 |
2,720 |
0,780 |
0,919 |
0,151 |
|
|
PtTx2123 |
8,000 |
4,299 |
1,692 |
0,313 |
0,767 |
0,593 |
|
|
PtTx2146 |
18,000 |
7,825 |
2,428 |
0,180 |
0,872 |
0,794 |
|
|
psyl 17 |
16,000 |
11,246 |
2,544 |
0,898 |
0,911 |
0,014 |
|
|
psyl 42 |
14,000 |
7,407 |
2,203 |
0,560 |
0,865 |
0,353 |
|
|
psyl 44 |
8,000 |
4,662 |
1,681 |
0,102 |
0,786 |
0,870 |
|
|
Spag 7.14 |
10,000 |
6,135 |
1,992 |
0,920 |
0,837 |
-0,099 |
|
|
Среднее |
12,385 |
7,091 |
2,096 |
0,459 |
0,838 |
0,458 |
|
|
Ошибка среднего |
±1,269 |
±0,756 |
±0,109 |
±0,082 |
±0,018 |
±0,094 |
|
Продолжение табл. 3
|
Популяция |
Локус |
Na |
Ne |
I Ho |
He |
F |
|||
|
Юшкозеро |
lw_isotig21953 |
12,000 |
9,191 |
2,306 |
0,060 |
0,891 |
0,933 |
||
|
lw_isotig04306 |
8,000 |
6,642 |
1,969 |
0,327 |
0,849 |
0,616 |
|||
|
lw_isotig04195 |
6,000 |
3,757 |
1,437 |
0,540 |
0,734 |
0,264 |
|||
|
lw_isotig27940 |
10,000 |
6,949 |
2,052 |
0,184 |
0,856 |
0,785 |
|||
|
PtTx 3107 |
9,000 |
6,859 |
2,032 |
0,320 |
0,854 |
0,625 |
|||
|
PtTx 3116 |
10,000 |
5,959 |
1,987 |
0,340 |
0,832 |
0,591 |
|||
|
PtTx 4001 |
18,000 |
11,737 |
2,630 |
1,000 |
0,915 |
-0,093 |
|||
|
PtTx2123 |
8,000 |
4,250 |
1,610 |
0,225 |
0,765 |
0,706 |
|||
|
PtTx2146 |
22,000 |
12,563 |
2,771 |
0,840 |
0,920 |
0,087 |
|||
|
psyl 17 |
11,000 |
7,278 |
2,141 |
0,480 |
0,863 |
0,444 |
|||
|
psyl 42 |
9,000 |
7,331 |
2,074 |
0,286 |
0,864 |
0,669 |
|||
|
psyl 44 |
3,000 |
1,994 |
0,731 |
0,020 |
0,499 |
0,960 |
|||
|
Spag 7.14 |
14,000 |
6,180 |
2,135 |
1,000 |
0,838 |
-0,193 |
|||
|
Среднее |
10,769 |
6,976 |
1,990 |
0,432 |
0,822 |
0,492 |
|||
|
Ошибка среднего |
±1,378 |
±0,814 |
±0,143 |
±0,091 |
±0,031 |
±0,103 |
|||
|
Муезерский |
lw_isotig21953 |
16,000 |
9,796 |
2,489 |
0,064 |
0,898 |
0,929 |
||
|
lw_isotig04306 |
7,000 |
3,993 |
1,595 |
0,375 |
0,750 |
0,500 |
|||
|
lw_isotig04195 |
6,000 |
4,558 |
1,622 |
0,600 |
0,781 |
0,231 |
|||
|
lw_isotig27940 |
11,000 |
6,558 |
2,092 |
0,214 |
0,848 |
0,747 |
|||
|
PtTx 3107 |
9,000 |
6,252 |
1,951 |
0,292 |
0,840 |
0,653 |
|||
|
PtTx 3116 |
14,000 |
3,962 |
1,862 |
0,143 |
0,748 |
0,809 |
|||
|
PtTx 4001 |
22,000 |
16,234 |
2,913 |
0,860 |
0,938 |
0,084 |
|||
|
PtTx2123 |
7,000 |
3,256 |
1,382 |
0,118 |
0,693 |
0,830 |
|||
|
PtTx2146 |
16,000 |
6,766 |
2,220 |
0,128 |
0,852 |
0,850 |
|||
|
psyl 17 |
9,000 |
4,717 |
1,756 |
0,000 |
0,788 |
1,000 |
|||
|
psyl 42 |
12,000 |
8,322 |
2,235 |
0,592 |
0,880 |
0,327 |
|||
|
psyl 44 |
2,000 |
1,855 |
0,653 |
0,000 |
0,461 |
1,000 |
|||
|
Spag 7.14 |
8,000 |
4,582 |
1,716 |
0,571 |
0,782 |
0,269 |
|||
|
Среднее |
10,692 |
6,219 |
1,884 |
0,304 |
0,789 |
0,633 |
|||
|
Ошибка среднего |
±1,465 |
±1,024 |
±0,153 |
±0,076 |
±0,033 |
±0,087 |
|||
|
Чупа |
lw_isotig21953 |
10,000 |
5,678 |
1,980 |
0,200 |
0,824 |
0,757 |
||
|
lw_isotig04306 |
6,000 |
3,986 |
1,545 |
0,586 |
0,749 |
0,217 |
|||
|
lw_isotig04195 |
6,000 |
3,657 |
1,433 |
0,517 |
0,727 |
0,288 |
|||
|
lw_isotig27940 |
9,000 |
5,263 |
1,875 |
0,467 |
0,810 |
0,424 |
|||
|
PtTx 3107 |
9,000 |
6,667 |
2,022 |
0,500 |
0,850 |
0,412 |
|||
|
PtTx 3116 |
10,000 |
4,931 |
1,871 |
0,357 |
0,797 |
0,552 |
|||
|
PtTx 4001 |
13,000 |
7,287 |
2,226 |
0,933 |
0,863 |
-0,082 |
|||
|
PtTx2123 |
4,000 |
1,772 |
0,818 |
0,103 |
0,436 |
0,763 |
|||
|
PtTx2146 |
16,000 |
9,836 |
2,501 |
0,700 |
0,898 |
0,221 |
|||
|
psyl 17 |
6,000 |
3,830 |
1,477 |
0,633 |
0,739 |
0,143 |
|||
|
psyl 42 |
7,000 |
4,462 |
1,670 |
0,586 |
0,776 |
0,244 |
|||
|
psyl 44 |
2,000 |
1,724 |
0,611 |
0,000 |
0,420 |
1,000 |
|||
Окончание табл. 3
Популяция Локус Na Ne I Ho He F
|
Spag 7.14 |
8,000 |
4,467 |
1,707 |
0,933 |
0,776 |
-0,203 |
||
|
Среднее |
8,154 |
4,889 |
1,672 |
0,501 |
0,743 |
0,364 |
||
|
Ошибка среднего |
±1,024 |
±0,608 |
±0,145 |
±0,079 |
±0,041 |
±0,094 |
||
|
Амбарнское |
lw_isotig21953 |
15,000 |
9,474 |
2,462 |
0,433 |
0,894 |
0,516 |
|
|
lw_isotig04306 |
6,000 |
3,315 |
1,443 |
0,733 |
0,698 |
-0,050 |
||
|
lw_isotig04195 |
6,000 |
3,689 |
1,476 |
0,433 |
0,729 |
0,405 |
||
|
lw_isotig27940 |
8,000 |
5,422 |
1,878 |
0,267 |
0,816 |
0,673 |
||
|
PtTx 3107 |
9,000 |
6,164 |
1,957 |
0,367 |
0,838 |
0,562 |
||
|
PtTx 3116 |
14,000 |
7,157 |
2,263 |
0,483 |
0,860 |
0,439 |
||
|
PtTx 4001 |
9,000 |
2,841 |
1,436 |
0,759 |
0,648 |
-0,171 |
||
|
PtTx2123 |
8,000 |
3,564 |
1,484 |
0,300 |
0,719 |
0,583 |
||
|
PtTx2146 |
11,000 |
5,590 |
1,970 |
0,633 |
0,821 |
0,229 |
||
|
Spag 7.14 |
10,000 |
7,229 |
2,094 |
0,967 |
0,862 |
-0,122 |
||
|
psyl 17 |
10,000 |
5,233 |
1,917 |
0,833 |
0,809 |
-0,030 |
||
|
psyl 42 |
8,000 |
5,389 |
1,823 |
0,600 |
0,814 |
0,263 |
||
|
psyl 44 |
2,000 |
1,034 |
0,085 |
0,033 |
0,033 |
-0,017 |
||
|
Среднее |
8,923 |
5,085 |
1,714 |
0,526 |
0,734 |
0,252 |
||
|
Ошибка среднего |
±0,937 |
±0,613 |
±0,163 |
±0,072 |
±0,062 |
±0,083 |
||
|
Общее значение по популяциям |
10,185 |
6,052 |
1,871 |
0,445 |
0,785 |
0,440 |
||
|
Ошибка среднего |
±0,565 |
±0,356 |
±0,065 |
±0,036 |
±0,018 |
±0,043 |
||
Примечание. Nа – среднее число аллелей на локус, Nе – эффективное число аллелей на локус, I – индекс Шенона, Hо – наблюдаемая гетерозиготность, Hе – ожидаемая гетерозиготность, F – индекс фиксации.
неоднородное распределение аллелей по сравнению с другими исследуемыми популяциями.
Наблюдаемая гетерозиготность (Ho = 0,445±0,036) во всех рассматриваемых нами популяциях ниже ожидаемой (He = 0,785±0,018). Исключение составляют локусы PtTx 4001 и Spag 7.14 для популяций Юшкозеро и Чупа, где наблюдаемая гетерозиготность выше ожидаемой.
Кроме того, во всех изучаемых популяциях наблюдается дефицит гетерозигот. Значения индекса фиксации (F) находятся в диапазоне 0,252–0,633. Наиболее выраженный дефицит обнаружен у популяций Амбарнское (F = 0,252) и Чупа (F = 0,364). В наиболее равновесном состоянии по распределению генотипов согласно правилу Харди–Вайнберга находится популяция Муезерский. О недостатке гетерозигот в северной части карельских популяций сосны также свидетельствуют показатели F-статистики Райта (табл. 4).
Так, значение индекса FIT – 0,482±0,079 свидетельствует о том, что в популяции присутствует дефицит гетерозигот. Предполагаем, что причиной избыточной гомозиготности популяций может быть эффект Валунда [20], когда при сильных различиях частот аллелей между популяциями доля гетерозигот уменьшается, даже если соотношение Харди-Вайнберга выполняется в каждой выборке, что ведёт к увеличению гомозигот. Но в отличие от инбридинга эффекту Валунда подвержена только частота гетерозигот исключительно в тех локусах, в которых имеется вариация аллельной частоты между субпопуляциями, когда при инбридинге может изменяться частота гетерозигот во всех локусах.
Значение индекса FIS > 0 (коэффициент инбридинга особей в субпопуляциях) указывает
Таблица 4. Значение F- статистики Райта для популяции сосны обыкновенной северной части Карелии
|
Локус |
F IS |
F IT |
F ST |
|
lw_isotig21953 |
0,781 |
0,791 |
0,043 |
|
lw_isotig04306 |
0,299 |
0,333 |
0,047 |
|
lw_isotig04195 |
0,307 |
0,328 |
0,030 |
|
lw_isotig27940 |
0,679 |
0,694 |
0,044 |
|
PtTx 3107 |
0,523 |
0,533 |
0,020 |
|
PtTx 3116 |
0,646 |
0,665 |
0,052 |
|
PtTx 4001 |
-0,012 |
0,049 |
0,060 |
|
PtTx2123 |
0,687 |
0,727 |
0,128 |
|
PtTx2146 |
0,432 |
0,465 |
0,058 |
|
psyl 17 |
0,308 |
0,364 |
0,082 |
|
psyl 42 |
0,375 |
0,399 |
0,038 |
|
psyl 44 |
0,929 |
0,956 |
0,372 |
|
Spag 7.14 |
-0,072 |
-0,034 |
0,035 |
|
Среднее |
0,453 |
0,482 |
0,078 |
|
Ошибка среднего |
±0,082 |
±0,079 |
±0,026 |
7 %
0,392
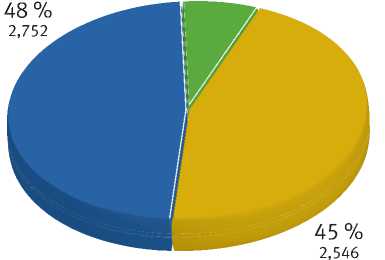
Между популяциями
Между индивидами
Внутри популяции
Рис. 2. Соотношение уровней внутривидового генетического разнообразия сосны обыкновенной северной части Карелии на редукцию гетерозиготности из-за возможного самоопыления и в среднем для всех изученных выборок составляет 0,453±0,082, хотя по отдельным локусам PtTx 4001 и Spag 7.14 редукция гетерозигот не отмечена.
Значение индекса FST – 0,078±0,026 (коэффициент инбридинга субпопуляций относительно всей популяции) указывает на слабую дифференциацию популяций и высокий поток генов внутри субпопуляций.
Результат AMOVA-анализа показал, что большая часть общей генетической дисперсии, обнаруженной на основе исследования 13 микроса-теллитных локусов, приходится на изменчивость внутри популяций. Так, поток генов между особями внутри популяции – 48 %, а внутри индивидов (самоопыление) – 45 % (рис. 2). Таким образом, внутрипопуляционная изменчивость составляет 93 %, а межпопуляционная – 7 %.
Для оценки уровня генетической дифференциации изучаемых популяций сосны обыкновенной проведены вычисления генетических дистанций М. Неи [21]. Результаты вычислений приведены на дендрограмме сходства, которая построена на основе матрицы генетических расстояний методом невзвешенного попарного арифметического среднего UPGMA [22] (рис. 3).
По результатам кластерного анализа популяций сосны обыкновенной можно выделить 2 большие группы. В состав самой большой группы (DN = 0,16) вошли 4 выборки – Муезерский, Амбарнское, Чупа и Юшкозеро. Эти популяции оказались генетически наиболее близкими, хотя внутри них можно выделить ещё одну более обособленную подгруппу (DN = 0,026), состоящую из двух популяций – Чупа и Юшкозеро. В отдельную группу выделена популяция Кевя-тозеро, которая выделяется высоким уровнем генетической обособленности (DN = 0,35). Кроме того, эта популяция отличается самым большим аллельным разнообразием из всех изученных популяций.
В целом выявленная с помощью 13 ядерных микросателлитных локусов популяционная структура сосны обыкновенной в северной части Республики Карелии свидетельствует о генетической близости исследованных популяций. Обособленность популяции Кевятозеро может отражать отличия в уровне генетического разнообразия этих популяций. Эти отличия могут быть связаны как с особенностями расселения сосны в послеледниковый период, так и с современными изменениями генетической структуры популяций, обусловленными такими условиями, как возрастающая мозаичность ареала сосны обыкновенной, восстановление лесов смешанным лесосеменным материалом, изменение климата.
Выводы
С помощью 13 ядерных микросателлитных локусов проведен анализ генетической структуры 5 естественных популяций: Чупа, Ам-барнское, Юшкозеро, Кевятозеро, Муезерский. Все использованные микросателлитные локусы полиморфны (Р99 % = 100 %) во всех популяциях и пригодны для использования в целях генетической паспортизации.
Всего обнаружено 114 аллелей. Наибольшее аллельное разнообразие выявлено в популяции Кевятозеро (Na = 12,385±1,269), наименьшее – в популяциях Амбарнское (Na = 8,923±0,937) и Чупа (Na = 8,1±1,024).
Эффективное число аллелей (Ne) варьирует от 4 до 7 на локус в среднем для всех рассматриваемых популяций. В целом по всем изученным п. Кевятозеро
|
0,35 |
п. Муезерский |
|||
|
0,16 |
Амбарнское с.п.н |
|||
|
0,13 |
0,026 |
|||
|
б.н.п. Чупа |
||||
|
п. Юшкозеро |
||||
|
0,4 |
Illi 0,35 0,3 0,2 0,15 0,1 0,05 0 DN |
|||
Рис. 3. Дендрограмма сходства популяций сосны обыкновенной в северной части Республики Карелии по матрице генетических расстояний Неи (DN )
выборкам аллельное разнообразие составляет 10±0,565 аллелей на локус, что показывает высокое аллельное разнообразие популяции сосны обыкновенной на исследованных участках.
Степень равномерности распределения аллельных вариантов внутри выборок всех исследуемых популяций достаточно однородна, индекс Шенона варьирует от 1,672 до 1,884.
Уровень наблюдаемой гетерозиготности (Ho)
во всех рассматриваемых популяциях ниже ожидаемой (He) за исключением некоторых локусов (PtTx 4001 и Spag 7.14) в популяциях Юшкозеро и Чупа, где она оказалась выше ожидаемой.
Результат AMOVA-анализа показал относительно невысокий уровень межпопуляционной дифференциации, внутрипо-пуляционная изменчивость составляет 93 %, а межпопуляционная – 7 %.
На основе кластерного анализа по матрице генетических расстояний М. Неи выявлена популяционная структура P. sylvestris в северной части Карелии. Все изученные популяции разделены на 2 группы. В состав первой группы вошли 4 популяции – Муезерский, Амбарнское, Чупа и Юшкозеро. Среди этих популяций можно идентифицировать подгруппу, состоящую из двух популяций – Чупа и Юшкозеро. В отдельную группу выделяется популяция Кевятозеро, эта группа отличается высоким уровнем генетической обособленности (DN = 0,35) в сравнении со всеми изученными популяциями.
Список литературы Генетическое разнообразие естественных популяций сосны обыкновенной в северной части Республики Карелии
- Гаврилова, О.И. Лесовосстановление вырубок и продуктивность лесных культур хвойных пород Республики Карелия / О.И. Гаврилова. - Петрозаводск, 2012. - 351 с.
- Ильинов, А.А. Состояние генофонда сосны обыкновенной Pinus sylvestris L. в Карелии / А.А. Ильинов, Б.В. Раевский // Сибирский лесной журнал. - 2016. - № 5. - С. 45-54.
- Генетическая изменчивость и дифференциация сосны обыкновенной Pinus sylvestris L. на территории Карелии / Ю.А. Янбаев, В.В. Тренин, З.Х. Шигапов, Б.А. Чистяков, Р.М. Бахтиярова // Научные основы селекции древесных растений Севера. - Петрозаводск: КНЦ РАН, 1998. - С. 25-32.
- Bilgen, B.B. Allozyme variations in six natural populations of scots pine (Pinus sylvestris) in Turkey / B.B. Bilgen, N. Kaya // Biología. - 2007. - Т. 62. - № 6. - С. 697-703.
- Genetic differentiation of Pinus sylvestris L. and Pinus mugo aggr. populations in Switzerland / C. Neet-Sarqueda [et al.] // Silvae Genetica. - 1994. - Т. 43. - № 4. - С. 207-214.
- Structure of Pinus sylvestris L. populations in Bulgaria revealed by chloroplast microsatellites and terpenes analysis: provenance tests / K.D. Naydenov [et al.] // Biochemical Systematics and Ecology. - 2005. - Т. 33. - № 12. -С.1226-1245.
- Genetic structure of montane isolates of Pinus sylvestris L. in a Mediterranean refugial area / J.J. Robledo-Arnuncio [et al.] // Journal of Biogeography. - 2005. - Т. 32. - № 4. - С. 595-605.
- Studies of allele frequencies and inbreeding in Scots pine populations by the aid of the isozyme technique. - Text: electronic / D. Rudin [et al.]. - 1974. - Режим доступа:www.thuenen.de
- Multiple origins for Scots Pine (Pinus sylvestris L.) in Scotland: evidence from mitochondrial DNA variation / W.T. Sinclair, J.D. Morman [et al.] // Heredity. - 1998. - 80:233-240. D0I:10.1046/j.1365-2540.1998.00287.x
- Калько, Г.В. ДНК-маркеры для оценки генетических ресурсов ели и сосны / Г.В. Калько // Труды Санкт-Петербургского научно-исследовательского института лесного хозяйства. - 2015. - № 4. - С. 19-34.
- Low-copy microsatellite markers for Pinus taeda L. / C.G. Elsik, V.T. Minihan, S.E. Hall, A.M. Scarpa, C.G. Williams // Genome. - 2000. - V. 43. - P. 550-555.
- Development and characterization of 25 EST-SSR markers in Pinus sylvestris var. mongolica (Pinaceae) / P. Fang [et al.] // Applications in plant sciences. - 2014. - Т. 2. - № 1. - DOI: 10.3732/apps.1300057.
- Cross-amplification and sequence variation of microsatellite loci in Eurasian hard pines / S.C. González-Martínez [et al.] // Theoretical and applied genetics. - 2004. - Т. 109. - № 1. - С. 103-111.
- Single-copy, species-transferable microsatellite markers developed from loblolly pine ESTs / C. Liewlaksaneeyanawin [et al.] // Theoretical and Applied Genetics. - 2004. - Т. 109. - № 2. - С. 361-369.
- Novel polymorphic nuclear microsatellite markers for Pinus sylvestris L. / F. Sebastiani [et al.] // Conservation Genetics Resources. - 2012. - Т. 4. - № 2. - С. 231-234.
- Peakall, R. Genalex 6: genetic analysis in Excel. Population genetic software for teaching and research / R. Peakall, P.E. Smouse // Molecular Ecol. Notes. - 2006. - N 6. - P. 288-295.
- Кузнецов, В.М. Информационно-энтропийный подход к анализу генетического разнообразия популяций (аналитический обзор) / В.М. Кузнецов // Аграрная наука Евро-Северо-Востока. - 2022. - Т. 23. - № 2. - С. 159-173.
- Sneath, P.H.A. Numerical Taxonomy.The Principles and Practice of Numerical Classification / P.H.A. Sneath, R.R. Sokal. - San Francisco: W.H. Freeman and Co., 1973. - 549 p.
- Microchecker: software for identifying and correcting genotyping errors in microsatellite data / С. Van Oosterhout, W. Hutchinson, D. Wills, P. Shipley // Molecular Ecol. Resources. - 2004. - № 4. - P. 535-538.
- Wahlund, S. Zusammensetzung von Populationen und Korrelationserscheinungen von Standpunkt der Vererbungslehre aus Betrachet / S. Wahlund // Hereditas. -1928. - V. 11. - P. 65-106.
- Nei, M. Genetic distance between populations / M. Nei // Amer. Natur. - 1972. - V. 106. - P. 283-292.
- Sinclair, W.T. Multiple origins for Scots pine (Pinus sylvestris L.) in Scotland: evidence from mitochondrial DNA variation / W.T. Sinclair, J.D. Morman, R.A. Ennos // Heredity. - 1998. - T. 80. - №. 2. - C. 233-240.
