Генетическое разнообразие и географическое распределение гаплотипов митохондриальной ДНК криптических видов лесных мышей подрода Sylvaemus Западного Кавказа

Автор: Амшокова Альбина Хасмановна, Темботова Фатимат Асланбиевна, Кучинова Екатерина Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 4-2 т.17, 2015 года.
Бесплатный доступ
Результаты генетического анализа лесных мышей Западного Кавказа, показали, что в его западной части, в пределах Республики Адыгея, симпатрично и симбиотопично обитают две генетически дивергированные линии мышей подрода Sylvaemus (A. uralensis и A. ponticus). В более теплых и влажных условиях равнины и предгорий, а также в лесостепном и поясе широколиственных лесов богатых разнообразными кормовыми ресурсами доминирует A. ponticus. В поясе субальпийских лугов и в пределах верхней границы широколиственных лесов, где биотопы, представлены каменистыми россыпями, напротив, малая лесная мышь. В целом вид A. ponticus на Западном Кавказе генетически более разнороден в сравнении с A. uralensis. Для A. uralensis всего Западного Кавказа установлено наличие сходных гаплотипов, отсутствие географического структурирования выборок, а также минимальные генетические дистанции между ними, что указывает на генетическое единство вида на исследованной территории.
Генетическое разнообразие, цитохром b, митохондриальная днк, генетическая дистанция, западный кавказ
Короткий адрес: https://sciup.org/148203894
IDR: 148203894 | УДК: 575:599.323.4
Genetic diversity and geographical distribution of mitochondrial DNA haplotypes in the cryptic species of common field mice from the subgenus Sylvaemus in the Western Caucasus
Genetic analysis of the common field mice in the Western Caucasus has shown that two genetically diverged lines of the mice from the subgenus Sylvaemus (A. uralensis and A. ponticus) in-habit sympatrically and symbiotically its western part, within the Republic of Adygei. A. ponticus dominates under warmer and more humid conditions of the plain and foothills, and in the forest steppe and broad-leaved belts abundant in varied food resources. A. uralensis prevails in the subalpine meadow belt and within the upper boundary of the broad-leaved forests where biotopes are represented by stony lands. In the Western Caucasus A. ponticus is on the whole genetically more heterogeneous as compared to A.uralensis. In A. uralensis from all Western Caucasus the existence of similar haplotypes, absence of geographical structure for the samples, and minimal genetic distances between them indicating genetic uniformity of the species in the territory under study, are determined.
Текст научной статьи Генетическое разнообразие и географическое распределение гаплотипов митохондриальной ДНК криптических видов лесных мышей подрода Sylvaemus Западного Кавказа
рома b мтДНК оценить уровень генетического разнообразия и степень внутри- и межвидовой генетической дифференциации лесных мышей подрода Sylvaemus Западного Кавказа; 2) исследовать географическое распределение гаплотипов лесных мышей подрода Sylvaemus Западного Кавказа; 3) изучить численное соотношение, особенности высотного и биотопического размещения видов-двойников лесных мышей в условиях Западного Кавказа.
В качестве молекулярного маркера выбран фрагмент гена цитохрома b митохондриальной ДНК, широко используемый для оценки внутри и межвидовой изменчивости позвоночных животных и реконструкции их филогении [1, 3-6].
МАТЕРИАЛЫ И МЕТОДЫ
В качестве материала для молекулярно-генетических исследований послужили образцы мышечной ткани мышей подрода Sylvaemus , фиксированные в 96% этиловом спирте . Сбор материала проводили на Западном Кавказе в разных, удаленных друг от друга локалитетах (рис. 1).
Общее количество образцов составило 178 экз. лесных мышей (Карачаево-Черкесская Республика: окр. с. В.Теберда – 10 экз.; Республика Адыгея: равнинные территории – окр. п. Северный – 3 экз., окр. ст. Кужорская – 5 экз., окр. г. Майкопа – 4 экз.; предгорные территории: окр. п. Усть – Сахрай – 7 экз., окр. ст. Даховская – 10 экз., окр. п. Хамышки – 58 экз., окр. п. Гузерипль – 13 экз.; среднегорные территории: окр. пл. Верхнее
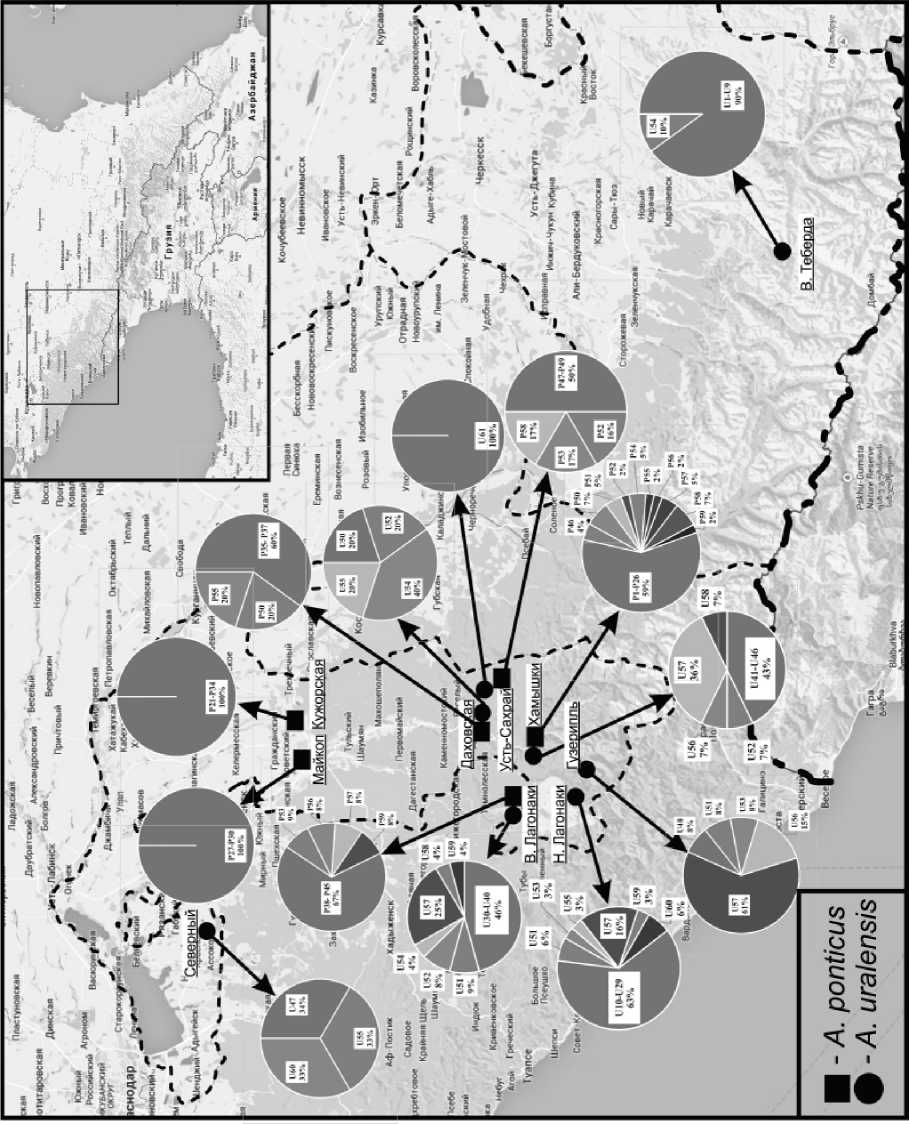
Рис. 1. Места сбора материала и распределение гаплотипов мтДНК криптических видов лесных мышей Западного Кавказа Примечание: гаплотипы обозначены: U - A. uralensis, Р - A.ponticus.
Лагонаки – 36 экз., окр. плато Нижнее Лагонаки – 32 экз.).
Сбор материала в Карачаево-Черкесской Республике проводили в окр. с. В. Теберда (правый борт р. В. Теберда) в сосново-разнотравно-злаковом лесу на высоте 1350 м над ур. м.
В республике Адыгея исследованиями охвачены равнинные, предгорные и среднегорные территории. На равнине исследованы три географических пункта окр. г. Майкопа, ст. Кужорская и п. Северный. В окр. г. Майкопа исследованиями охвачен участок леса на высоте 148 м над ур. м. В видовом составе древесной растительности преобладали дуб и береза, также встречались лещина, боярышник, чубушник кавказский. Под пологом леса хорошо выражен подрост лиственных деревьев. В ст. Кужорская отлов зверьков проводили в пойменном мертвопокровном лесу на высоте 229 м над ур. м. Среди древесной породы доминирующими видами являлись дуб, береза, осина и клен. Травянистая растительность представлена слабо, под деревьями значительный опад, состоящий из сухой листвы, веточек, кусочков коры и т.д. В п. Северный материал собирался в лесополосе, рядом с агроценозом на высоте 279 м над ур. м. Древесная растительность в основном была представлена плодовыми деревьями (алыча, яблоня, груша, ясень, боярышник), основную долю травянистой растительности составляли злаки.
В предгорных условиях Адыгеи в п. Усть -Сахрай исследования проводили в широколиственном лесу. Основными образующими породами являются дуб черешчатый, клен канадский. Высота над уровнем моря – 525 м. В окр. ст. Даховская первичный материал собирался на высоте 534 м над ур. м в грабово-буковом лесу с примесью лещины, дуба черешчатого, ивы. На данном участке из травянистой растительности произрастают – мятлик луговой, тонколучник обыкновенный, цикорий обыкновенный, репешок аптечный, тмин рассеченнолистный, мята длиннолистная. В п. Хамышки отлов зверьков проводили в буково-дубовом лесу на высоте 604 м. над ур. м. Травянистая растительность представлена тимофеевкой луговой, мятликом, борщевиком, ясноткой, клевером ползучим и т.д. В отличие от описанных выше точек в окр. п. Гузерипль (653 м над ур. м) биотоп представлен каменистыми россыпями. Валуны, частично покрытые мхом, достигают в диаметре 1,5-2 метров. Древесная растительность представлена дубом черешчатым, мелколиственницей, ольхой, дикой яблоней, грушей и т.д. Травянистый ярус – дикая астра, мята, крапива двудомная, белокопытник.
В среднегорных экосистемах республики Адыгея сбор первичного материала проводили в субальпийском поясе – окр. пл. Лагонаки на высоте 1800 м над ур. м. Учитывая биотопиче-ское разнообразие плато Лагонаки было условно разделено на два участка Нижнее и Верхнее
Лагонаки. В Нижнем Лагонаки биотоп представлен каменистыми россыпями. Древесная растительность в основном состоит из березы Радде с примесью сосны Коха, ивы козьей и т.д. Из травянистой растительности на данной территории произрастают – малина Буша, мятлик луговой, подмаренник, крушина ломкая, герань Роберта, мятлик луговой и т.д. В Верхнем Лагона-ки материал собирался в смешанном лесу (пихта, бук, клен, ясень). Травянистая растительность представлена следующими видами – папоротник, девясил высокий, щавель конский, ежа сборная, просо куриное, крапива двудомная и т.д.
Для молекулярно-генетического анализа использовали участок гена цитохрома b мтДНК длиной (732 пн). Выделение ДНК проводили с использованием прибора для автоматического выделения нуклеиновых кислот King Fisher Flex (Финляндия) и набора для выделения ДНК «Magna DNA Prep100» (Изоген, Москва), основанном на применении магнитных частиц. Амплификация фрагментов ДНК проводилась с использованием набора MasterMix Х5 (Диалат, Москва). Использованные в работе праймеры (L-14724 CGAAGCTT GATATGAAAAACCATCGTTG и H-15573 AATAGGAAGTATCATTCGGGTTTGATG) взяты из статьи А.Е. Балакирева с соавторами (2007) [1]. Полимеразную цепную реакцию (ПЦР) проводили в следующем режиме:
-
1) первичная денатурация при 95 о С – 4 мин.,
-
2) 35 циклов с 60 секундной денатурацией при 94 о С, 40 секундным отжигом при 50 о С и 60 секундной элонгацией при 72 о С,
-
3) 10 минутная элонгация при 72 о С.
Амплификацию проводили в термоциклере Tetrad 2 (Bio-Rad). Очистку продукта ПЦР проводили с использованием реакционной смеси ацетата аммония с этанолом (NH 4 Ac+EtOH конечная концентрация аммония – 0,125М, этанола – 70%).
Результаты амплификации ДНК анализировали с помощью электрофореза в 1,5 % агарозном геле и буфере ТВЕ с добавлением бромистого этидия.
Для проведения секвенирования использовались те же праймеры. Определение нуклеотидных последовательностей ДНК проводили на секвенаторе ABI 3130 (Applied Biosystems, США) с использованием набора BigDye Terminator Kit 3.1 (Applied Biosystems, США). Выравнивание полученных последовательностей осуществляли вручную с помощью программы BioEdit [7]. Генетические дистанции и значения нуклеотидного разнообразия (π) рассчитывались в программе Mega 4.3 [8]. Разнообразие гаплотипов в выборке рассчитывали по формуле Нея [9]: H=N (1-∑ xi2)/ (N-1), где N – число особей в выборке, а xi – частота гаплотипа в выборке. Построение филогенетических деревьев осуществляли методом ближайшего связывания (Neighbor-Joining) c использованием модели Тамура-Нея. Медианную сеть гаплотипов строили с помощью программы Network 4.6.1[10].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Нуклеотидный состав исследованного фрагмента гена цитохрома b (аденин – 31.4%, тимин – 29.0%, цитозин – 25.8%, гуанин – 13.8%) сходен с таковыми других мелких млекопитающих [11-12], включая разные виды лесных и полевых мышей, особенностью которых является низкое содержание гуаниновых оснований [13]. При этом соотношение оснований в разных позициях различается. Встречаемость азотистых оснований в первой позиции – аденин – 30.4%, тимин – 25.4%, цитозин – 20.3%, гуанин – 23.8%, вторая позиция характеризуется высоким содержанием тимина (37.5%). Соотношение нуклеотидных оснований в третьей позиции сдвинуто в сторону избытка аденина (41.5%) и недостатка гуанина (3%). Всего для всей выборки выявлено 174 вариабельных сайтов из которых 133 были парсимониально-информативными, синглетонов было 41. По данному фрагменту описано 120 гаплотипов.
Для выяснения филогенетических отношений между описанными гаплотипами было построено дерево ближайшего связывания. Дендрограмма четко демонстрирует отделение гаплогруппы 1 от гаплогруппы 2. В пределах каждого кластера отмечается совершенно беспорядочное распределение особей, т.е. отсутствуют какие-либо региональные группировки, имеющие достоверную бутстреп-поддержку. Хотя большинство гаплотипов лесных мышей образуют две основные группы, тем не менее в пределах каждого кластера выделяются отдельные гаплотипы несколько дистанцированные от этих групп. Генетическая дистанция между двумя основными гаплогруп-пами составила 12%, что соответствует уровню межвидовых различий. Согласно данным ряда авторов [14-15], межвидовая дивергенция гена cyt b мышевидных грызунов варьирует в пределах от 6 до 15%. Таким образом, полученная дистанция достигает уровня межвидовых различий, а выявленные гаплогруппы соответствуют двум видам – A. uralensis и А. ponticus , что не противоречит существующим на сегодняшний день данным касательно видового состава лесных мышей Западного Кавказа [1, 16] . Однако, в отличие от нас, данные авторы для обозначения географической формы A. uralensis населяющей Кавказ используют эпитет « A. сiscaucasicus ». Ниже приводятся результаты молекулярно-генетического анализа и географическое распределение гаплотипов каждого вида.
Внутривидовая изменчивость малой лесной мыши (А. uralensis) Западного Кавказа.
Из 178 исследованных экз. лесных мышей Западного Кавказа 103 экз. относились к виду (А.
uralensis). В исследованной выборке обнаружено 60 гаплотипов гена цитохрома b, которые определяются полиморфизмом в 77 позициях мтДНК, 29 из которых являются парсимониально-инфор-мативными. Большинство мутации составляют транзиции (59), трансверсии выявлены в 18 позициях. У выявленных 60 гаплотипов значимую долю составляют уникальные гаплотипы (83%) и только 10 (17%) встречаются в нескольких образцах (2-24). Каждая исследованная выборка характеризуется уникальными гаплотипами свойственными только ей (рис. 1). Наибольшее количество уникальных гаплотипов (90%) отмечается в выборке из окр. с. В. Теберда. Гаплотипы U1-U9 встречаются только в данной выборке. У животных ст. Даховская отмечается наименьшее количество уникальных гаплотипов – 20%. В данной выборке отмечен только один оригинальный гаплотип U50 не характерный для других выборок. К числу таковых в выборке из окр. пл. В. Лагонаки относятся гаплотипы U30-U40, а в выборке из окр. пл. Н. Лагонаки U10 – U29. Доля уникальных гаплотипов в отмеченных выборках в первом случае составляет 46%, а во втором 63%. В выборке из п. Хамышки уникальными являются 6 гаплотипов (U41-U46), в выборке из равнинной Адыгеи - п. Северный уникальным является 1 гаплотип (U47) и одним уникальным гаплотипом (U49) отличается от всех остальных выборка из окр. п. Гузерипль. Уникальными являются и гаплотипы зверьков из п. Усть-Сахрай (n=1) и равнинной Адыгеи – ст. Кужорская (n=1).
Как видно из рис. 1, лесные мыши с идентичными гаплотипами отмечены в выборках из окр. пл. В. Лагонаки, Н. Лагонаки и п. Гузерипль (U51), с гаплотипом (U52) в выборках из окр. п. Хамышки, пл. В. Лагонаки и ст. Даховской. Гаплотип U53 встречается в двух – окр. пл. Н. Лагонаки и п. Гузерипль, а U54 в трех выборках - окр. пл. В. Лагонаки, ст. Даховской и с. В. Теберда. Сходство по гаплотипу U55 отмечается в трех выборках – ст. Даховская, п. Северный, пл. Н. Лагонаки, а по гаплотипу U56 в двух выборках п. Хамышки и п. Гузерипль. Гаплотип U57 является общим для четырех – п. Хамышки, пл. В. Лагонаки, пл. Н. Ла-гонаки и п. Гузерипль, U 58 для двух - п. Хамышки, пл. В. Лагонаки, U59 также для двух выборок - пл. В. Лагонаки, пл. Н. Лагонаки и U60 встречается в выборках из п. Северный и пл. Н. Лагонаки.
В таб. 1 представлено гаплотипическое и нуклеотидное разнообразие выборок A. uralensis . Разнообразие гаплотипов в выборках варьировало от 0.63 до 1.00 (ср. 0.91).
Из таб. 1 видно, что выборка лесных мышей из п. Гузерипль отличается от других несколько меньшими показателями гаплотипического и нуклеотидного разнообразия, а выборка лесных мышей из окр. с. В. Теберда напротив характеризуется повышенными значениями нуклео-
Таб. 1. Генетическое и гаплотипическое разнообразие A. uralensis Западного Кавказа
|
Географические группировки (размер выборки) |
Число гаплотипов |
Нуклеотидное разнообразие |
Гаплотипическое разнообразие |
|
A. uralensis |
|||
|
окр. п. Хамышки (n=14) |
10 |
0.006±0.00 |
0.89 |
|
окр. ст. Даховская (n=5) |
4 |
0.005±0.002 |
0.90 |
|
окр. п. Северный (n = 3) |
3 |
0.007±0.002 |
1.00 |
|
окр. п. Гузерипль (n=13) |
5 |
0.004±0.001 |
0.63 |
|
окр. пл. В. Лагонаки (n=24) |
17 |
0.006±0.002 |
0.94 |
|
окр. пл. Н. Лагонаки (n=32) |
25 |
0.008±0.002 |
0.98 |
|
окр. с. В. Теберда (n=10) |
10 |
0.010 ±0.002 |
1.00 |
тидного и гаплотипического разнообразия по сравнению с другими выборками. В целом для выборок, характерны невысокие значения нуклеотидного разнообразия при высоких значениях гаплотипического разнообразия. Высокие значения гаплотипического и низкие нуклеотидного разнообразия характерны для популяций с быстрым ростом численности от изначально небольшого числа основателей в случае, когда прошло достаточное время для восстановления гаплотипической изменчивости за счет мутационного процесса, но недостаточное для того, чтобы накопились значительные различия между нуклеотидными последовательностями [5, c. 328].
Генетические дистанции, полученные при попарном сравнении выборок A. uralensis из разных локалитетов Западного Кавказа, показали, что дистанции низки и колеблются в пределах от 0.000 до 0.008 (табл. 2). Исключение составляет одна особь из КЧР, с. В. Теберда (Teb.1) которая в несколько большей степени отличается от особей всех изученных выборок.
Дистанции, полученные при сравнении данной особи с особями из остальных географических выборок, варьировали от 0.015 до 0.021 (табл. 2). Однако считаем преждевременным, делать какие–либо выводы о наличии внутривидовых группировок, поскольку в данном случае речь идет о единственной особи.
Внутривидовая изменчивость кавказской лесной мыши (А.ponticus) Западного Кавказа.
В условиях Западного Кавказа исследовано 75 экз. кавказской лесной мыши. Для исследованного фрагмента последовательности гена цитохрома b мт ДНК длиной 732 п.н. выявлено 97 вариабельных сайтов (мутаций). Среди вариабельных сайтов отмечены 79 транзиции (Ts) и 22 трансверсии (Tv).
Состав нуклеотидных оснований гена цитохрома b – T-29.3%, A-25.5%, C-31.4%, G-13.8%. Как и у Apodemus uralensis насыщенность разных позиции кодона гуаниновыми основаниями различна: в первой – 23.2%, во второй – 14.9 %, в третьей – 3.3. В проанализированной выборке А.ponticus обнаружено 59 гаплотипов. Доля уникальных гаплотипов варьировала от 50 до 100 %. Из рис. 1 видно, что все особи, встреченные в условиях равнинной Адыгеи (окр. г. Майкопа и ст. Кужорская) являются носителями оригинальных не отмеченных в других выборках гаплотипов. Гаплотипы Р1-Р26 встречаются только в выборке из п. Хамышки и отсутствуют в других выборках.
Таб. 2. Средние генетические дистанции между выборками Apodemus uralensis Запаного Кавказа , рассчитанные по 732-пн участку гена цитохрома b
|
Обозначения выборок |
Ham. |
Guz. |
US |
Dah. |
VL |
NL |
Sev. |
Kuch. |
Teb. |
Teb1. |
|
Ham. |
0.000 |
0.002 |
0.001 |
0.000 |
0.000 |
0.001 |
0.003 |
0.001 |
0.005 |
|
|
Guz. |
0.000 |
0.002 |
0.002 |
0.000 |
0.000 |
0.002 |
0.003 |
0.001 |
0.005 |
|
|
US |
0.004 |
0.005 |
0.001 |
0.002 |
0.001 |
0.002 |
0.003 |
0.001 |
0.005 |
|
|
Dah. |
0.003 |
0.004 |
0.002 |
0.001 |
0.001 |
0.001 |
0.003 |
0.000 |
0.004 |
|
|
VL |
0.000 |
0.000 |
0.003 |
0.002 |
0.000 |
0.001 |
0.002 |
0.001 |
0.005 |
|
|
NL |
0.000 |
0.001 |
0.003 |
0.001 |
0.000 |
0.001 |
0.002 |
0.000 |
0.004 |
|
|
Sev. |
0.003 |
0.004 |
0.005 |
0.003 |
0.003 |
0.001 |
0.003 |
0.001 |
0.005 |
|
|
Kug. |
0.007 |
0.008 |
0.008 |
0.007 |
0.006 |
0.006 |
0.008 |
0.002 |
0.005 |
|
|
Teb. |
0.002 |
0.003 |
0.002 |
0.000 |
0.001 |
0.001 |
0.003 |
0.005 |
0.004 |
|
|
Teb1. |
0.017 |
0.018 |
0.016 |
0.015 |
0.016 |
0.015 |
0.018 |
0.021 |
0.015 |
Примечания: под диагональю – значения межгрупповых дистанций, над диагональю – соответствующие значения стандартной ошибки; обозначения выборок: 1. Нam. – п. Хамышки; 2. Guz. – п. Гузерипль; 3. US – п. Усть–Сахрай; 4. Dah. - ст. Даховская; 5. VL – пл. В. Лагонаки; 6. NL – пл. Н. Лагонаки; 7. Sev. – равнинная Адыгея, п. Северный; 8. Кug. – равнинная Адыгея, ст. Кужорская; 9. Teb. – КЧР, c. В. Теберда; 10. Teb1. – КЧР, c. В. Теберда
Таковыми в выборке из окр. пл. В. Лагонаки являются Р38-Р45. По три уникальных гаплотипа описаны и в выборках из окр. ст. Даховская (Р35-Р37) и п. Усть-Сахрай (Р47-Р49).
Сходные гаплотипы отмечаются как в пределах одной выборки, так и в разных выборках. Так, гаплотип Р46 отмечен у двух особей выборки п. Хамышки, то же самое касается и гаплотипов Р51 и Р54. Сходство по трем гаплотипам (Р56, Р57, Р59) отмечается между выборками лесных мышей из окр. п. Хамышки и пл. В. Лагонаки, еще два гаплотипа (Р52, Р58) являются общими для выборок из окр. п. Хамышки и п. Усть-Сахрай (рис. 1). Гаплотипы (Р50, Р55) встречены у зверьков из окр. п. Хамышки и ст. Даховская, а гаплотип Р53 у животных п. Усть-Сахрай и пл. В. Лагонаки.
Гаплотипическое разнообразие, отражающее количество и частоту встречаемости разных гаплотипов в выборах A. ponticus достаточно высокое 0.99-1.0 (табл. 3). Также для данных выборок характерны и высокие значения нуклеотидного разнообразия, существенно превышающие таковые A. uralensis. Таким образом, в отличие от A. uralensis практически все выборки A. ponticus характеризуются высокими значениями нуклеотидного и гаплотипического разнообразия, что имеет место в двух случаях: 1) при стабильности в течение длительного времени генетически целостных и экспансивных популяции с высокими значениями эффективной численности; 2) для популяций сформировавшихся путем объединения раннее изолированных генетически неоднородных группировок [17-19].
Филогенетическое дерево, построенное для географических выборок A. ponticus показало, что 15 особей, происходящие из различных пунктов Западного Кавказа, отделяются от общего пула, образуя одиночные кластеры с разным уровнем бутстреп-поддержки (рис. 2). При попарном сравнении отмеченных особей между собой дистанции варьировали от 0.008 до 0.057, примерно такого же порядка дистанции (0.002 – 0.044) получены при сравнении этих особей с особями из основных географических выборок из п. Усть-Сахрай, ст. Даховская, п. Хамышки и пл. В. Лагонаки (табл. 4). Минимальными оказались дистанции (0.000), полученные при сравнении между собой отмеченных выше четырех выборок.
Таким образом, внутривидовые различия в пределах вида А. ponticus значительны и в некоторых случаях достигают генетических дистанции выявленных между расами A. uralensis по фрагменту цитохрома b (0.057) [20].
Распространение. Как видно из рис. 1, на Западном Кавказе в пределах Республики Адыгея оба вида A. ponticus и A. uralensis имеют широкое распространение на всей территории республики, причем не только симпатрическое, но и симби-отопическое. Изложенное, касается как равнинных, так и горных (предгорных и среднегорных) территорий. Так, в большинстве изученных точек в пределах Республики Адыгея, оба вида регистрировались вместе (окр. ст. Кужорская, с. Хамышки, ст. Даховская, п. Усть-Сахрай, окр. п. В. Лагонаки). При этом они занимали самые разные ландшафты: степной зоны (окр. ст. Кужорская), лесные предгорные (с. Хамышки, ст. Даховская, п. Усть-Сахрай) и среднегорные (пл. В. Лагонаки). Численное соотношение видов на отмеченных территориях различается, но практически во всех предгорных и равнинных биотопах доминирует кавказская лесная мышь: окр. п. Усть-Сахрай 6:1, окр. с. Хамышки 3:1, окр. ст. Кужорская 4:1, или равна по численности с малой лесной, как это установлено в окр. ст. Даховской 1:1. В среднегорьях Западного Кавказа (окр. пл. В. Лагонаки), напротив, многочисленна малая лесная мышь 3:1, а в Н. Лагонаки встречен только A. uralensis.
Учитывая, что районы В. Лагонаки и Н. Ла-гонаки находятся на одном и том же высотном уровне, можно предположить, что одним из факторов, лимитирующих распространение A. ponticus в окр. пл. Н. Лагонаки является отсутствие подходящих биотопов для данного вида. Исследованные территории окр. пл. Н. Лагонаки в основном были представлены каменистыми биотопами, наиболее подходящими для обитания полевок, нежели лесных мышей. Примерно такие же ландшафты характерны и для п. Гузерипль, где вид A. ponticus также не обнаружен. Все 13 экз. лесных мышей, отловленных на данной территории и генетически датированных, относились к виду A. uralensis . В среднегорьях (окр. с. В. Теберда) восточной части Западного Кавказа в пределах Карачаево-Черкессии также представлен только один вид – A. uralensis .
Таб. 3. Генетическое и гаплотипическое разнообразие A. ponticus Западного Кавказа
|
Географические группировки (размер выборки) |
Число гаплотипов |
Нуклеотидное разнообразие |
Гаплотипическое разнообразие |
|
A. ponticus |
|||
|
окр. п. Хамышки (n=44) |
36 |
0.009±0.001 |
0.99 |
|
окр. ст. Даховская (n=5) |
5 |
0.022±0.004 |
1.00 |
|
окр. г. Майкоп (n = 4) |
4 |
0.025±0.004 |
1.00 |
|
окр. ст. Кужорская (n=4) |
4 |
0.031±0.005 |
1.00 |
|
окр. пл. В. Лагонаки (n=12) |
11 |
0.013±0.002 |
0.99 |
|
окр. п. Усть-Сахрай (n=6) |
6 |
0.007±0.002 |
0.99 |
2586H1
2585H1
2164H1
34us1

2193VL1
2167H1
2150H1
2169H1
2375VL1
2177H1
2203H1
2204H1
35us1
2154H1
2189VL1
2361H1
2163H1
43Dah1
2455H1
2155H1
2166H1
2172H1
2184H1
2178H1
33us1
2170H1
2205H1
2161H1
2409H1
2202H1
2453H1
32us1
2454H1
2395VL1
2201H1
2160H1
2520H1
2152H1
2175H1
2376VL1
2H1
2174H1
273Les1
2403us1
2165H1
2168H1
2430VL1
2149H1
2151H1
2182VL1
2186VL1
2411H1
37Dah1
2195VL1
36us1
40Dah1
2158H1
2450VL1
2618H1
2410H1
2176H1
2437VL1
240Kug1
2171H1
2156H1
275Les1
272Les1
263Kug1
39Dah1
44Dah1
274Les1
2396VL1
261Kug1
252Kug1
2162H1
0.005
Рис. 2. Дендрограмма сходства гаплотипов фрагмента гена цит. b мтДНК кавказской лесной мыши, построенная методом ближайшего связывания (Neighbor-Joining) с использованием двухпараметрической модели Кимуры (Kimura, 1980). Места сбора животных обозначены геометрическими фигурами:
- п. Хамышки 1 (H1), □ - п. Усть-Сахрай 1 (Usl); □ - пл. В. Лагонаки1 (V.Lag.l),
О - ст. Кужорская (Kugl), О - окр. г. Майкопа (Les1), В - ст. Даховская1 (Dahl)
Таб. 4. Средние генетические дистанции между выборками лесных мышей Apodemus ponticu s ( Sylvaemus) Западного Кавказа , рассчитанные по 732-пн участку гена цитохрома b
|
_ 1 ^ CXI 5 CXI |
О О о о |
о о о о |
о о о о |
о о о о |
^f о о о |
о о о о |
о о о |
со о о о |
со о о о |
о о о |
о о о |
\О о о о |
о о о |
о о о |
со о о о |
to о о о |
о о о |
о о о |
о о о |
|
|
| S ^ ^ CX] |
хО О о о |
о о о |
о о о |
\О о о о |
о о о |
о о о |
о о о |
со о о о |
\О о о о |
\О о о о |
о о о |
о о о |
о о о |
\О о о о |
о о о о |
о о о |
\О о о о |
о о о |
о СХ] о о |
|
|
oo S 2 xi |
СХ] О о о |
СХ] о о о |
СХ] о о о |
СХ] о о о |
\О о о о |
СХ] о о о |
со о о о |
о о о |
о о о |
\О о о о |
^t о о о |
о о о |
со о о о |
со о о о |
о о о |
\О о о о |
\О о о о |
о to о о |
о СХ] о о |
|
|
H S (M |
\О О о о |
о о о |
о о о |
о о о |
о о о |
\О о о о |
о о о |
о о о |
о о о |
о о о |
о о о |
\О о о о |
\О о о о |
\О о о о |
о о о |
^ о о о |
^ СХ] о о |
^ СХ] о о |
со о о |
|
|
bO О u4 О ^ № CX] |
\О О о о |
\О о о о |
\О о о о |
\О о о о |
о о о |
\О о о о |
\О о о о |
со о о о |
о о о |
\О о о о |
о о о |
о о о |
о о о |
о о о |
со о о о |
о о о |
о СХ] о о |
о СХ] о о |
со о о о |
|
|
Й о m 5 xf rH M CX| |
о о о |
о о о |
о о о |
о о о |
со о о о |
о о о |
о о о о |
о о о о |
о о о |
о о о |
о о о |
со о о о |
о о о |
со о о о |
о to о о |
^ to о о |
^ to о о |
о ^t о о |
СХ] ^ о о |
|
|
M ex, Xf 5 m ^ч M CXI |
со о о о |
со о о о |
со о о о |
\О о о о |
\О о о о |
со о о о |
\О о о о |
о о о о |
о о о |
о о о |
со о о о |
о о о |
\О о о о |
СХ] ^ о о |
о о |
^ СХ] о о |
4 О о |
о СХ] о о |
о СХ] о о |
|
|
to xO ^ № CX] |
со о о о |
со о о о |
со о о о |
со о о о |
\О о о о |
со о о о |
о о о |
со о о о |
со о о о |
\О о о о |
о о о о |
о о о |
^f СХ] о о |
о ^t о о |
о to о о |
о СХ] о о |
о о |
to to о о |
to о о |
|
|
\O CX] >—1 tO '—I H> CXI |
о о о |
о о о |
о о о |
о о о |
\О о о о |
о о о |
о о о |
со о о о |
о о о |
\О о о о |
о о о |
to о о |
о о |
СХ] ^ о о |
о СХ] о о |
^ СХ] о о |
^ to о о |
^ to о о |
to СХ] о о |
|
|
^ H ^ ^ч ?> CXI |
^f о о о |
^f о о о |
^f о о о |
^f о о о |
о о о |
^ о о о |
со о о о |
со о о о |
о о о |
о о о |
СХ] о о |
СХ] о о |
о о |
о to о о |
о to о о |
^ to о о |
^t о о |
о ^t о о |
to о о |
|
|
S 1 3 |
\О о о о |
\О о о о |
\О о о о |
\О о о о |
о о о |
о о о |
о о о |
о о о |
\О о о о |
СТ' to о о |
о to о о |
^f СХ] о о |
о to о о |
to о о |
СХ] о о |
о СХ] о о |
о о |
о to о о |
^f СХ] о о |
|
|
o Q m |
\О о о о |
\О о о о |
\О о о о |
\О о о о |
\О о о о |
о о о |
со о о о |
о о о о |
о о |
to ^ о о |
о to о о |
СХ] о о |
о to о о |
^t о о |
со о о |
о СХ] о о |
СХ] to о о |
о to о о |
to о о |
|
|
8 £ ОО К CXI |
о о о |
о о о |
о о о |
о о о |
о о о |
о о о |
со о о о |
со ^t о о |
to о о |
о ^t о о |
to ^ о о |
о о |
о ^t о о |
о о |
to о о |
to о о |
to to о о |
со ^t о о |
to о о |
|
|
▻- к ^ |
со о о о |
со о о о |
со о о о |
со о о о |
о о о |
со о о о |
о ^t о о |
СТ' ^ о о |
о СХ] о о |
о о |
о СХ] о о |
СТ' to о о |
to СХ] о о |
о о |
о СХ] о о |
to to о о |
о о о |
о ^t о о |
to о о |
|
|
8 00 cd О К CXI |
СХ] о о о |
СХ] о о о |
СХ] о о о |
СХ] о о о |
\О о о о |
СТ' о о |
ьо о о |
ьо ьо о о |
СТ' СХ] о о |
ьо о о |
^f to о о |
о о |
4 О о |
^ to о о |
о СХ] о о |
^ СХ] о о |
о to о о |
о о |
о о |
|
|
8 40 u-> К CXI |
о о о |
о о о |
о о о о |
о о о о |
о СХ] о о |
ьо ьо о о |
о ьо о о |
СХ] о о |
о СХ] о о |
о to о о |
о о |
о to о о |
^f СХ] О О |
to о о |
СХ] о о |
о СХ] о о |
^ СХ] о о |
^ СХ] о о |
о о о |
|
|
о о о о |
о о о о |
о о о о |
ьо СХ] о о |
ьо о о о |
со ^t о о |
СХ] ьо о о |
СТ' СХ] о о |
о СХ] о о |
^f о о |
to to о о |
to ^ о о |
СХ] О О |
^ to о о |
CN о о |
СХ] СХ] о о |
СХ] о о о |
СХ] to о о |
CN о о |
||
|
Cd to Q |
о о о о |
о о о о |
о о о о |
ьо СХ] о о |
^ о о о |
со ^t о о |
ьо ьо о о |
CN о о |
о СХ] о о |
^f о о |
to to о о |
4 О о |
СХ] ^ О О |
^ to о о |
^ СХ] о о |
to СХ] о о |
СХ] о о о |
to to о о |
^f СХ] о о |
|
|
s CXI К |
о о о о |
о о о о |
о о о о |
ьо СХ] о о |
^ о о о |
СТ' о о |
СХ] ьо о о |
о ьо о о |
о о |
^f о о |
^f to о о |
4 О о |
to о о |
to to о о |
CN о о |
to СХ] о о |
СХ] о о о |
СХ] to о о |
CN о о |
|
|
co ^ D |
о о о о |
о о о о |
о о о о |
ьо СХ] о о |
ьо о о о |
СТ' о о |
СХ] ьо о о |
СТ' СХ] о о |
о о |
ьо о о |
^f to о о |
to ^ о о |
to ^ о о |
to to о о |
CN о о |
to СХ] о о |
СХ] о о о |
СХ] to о о |
CN о о |
|
|
1—1 |
СХ] |
to |
'ф |
in |
о |
ь- |
со |
СТ' |
о |
1—1 |
СХ] |
ьо |
'ф |
1-0 |
о |
1—1 |
со |
о |
о СХ] |
Из полученных данных можно заключить вид A. uralensis широко распространен на территории Западного Кавказа и отмечается на всех исследованных участках, тогда как вид A. ponticus, встречается только на некоторых обследованных территориях (рис.1). В условиях среднегорий Западного Кавказа одним из основных факторов, ограничивающих распространение второго вида, является не столько высота над уровнем моря, сколько наличие подходящих для обитания данного вида биотопов. Полученные результаты согласуются с данными приводимыми в работе Н.М. Окуловой с соавторами (2005), согласно которым в низкогорных высокопродуктивных орехоплодных широколиственных лесах (бук, граб, дуб, каштан) кавказская лесная мышь является преобладающим по численности, а в менее продуктивных орехоплодных лесах среднегорья напротив доминирующим по численности видом являетсямалая лесная мышь [21].
ЗАКЛЮЧЕНИЕ
Полученные данные по генетическому разнообразию лесных мышей Западного Кавказа показали, что в его западной части, в пределах Республики Адыгея, обитает две генетически значительно дивергированные линии мышей подрода Sylvaemus , генетические дистанции между которыми весьма существенны (около 12%) и достигают уровня межвидовой дифференциации. Выявленные гаплогруппы соответствуют двум видам A. uralensis и A. ponticus . В восточной части Западного Кавказа, в Карачаево-Черкессии, выявлено обитание только малой лесной мыши.
Для малой лесной мыши всего Западного Кавказа установлено наличие сходных гаплотипов, отсутствие географического структурирования выборок, а также минимальные генетические дистанции между ними, что указывает на генетическое единство вида на исследованной территории.
A. ponticus на Западном Кавказе генетически более разнороден по структуре мтДНК в сравнении с A. uralensis . Среди особей A. ponticus отмечаются единичные животные, существенно выделяющиеся в выборках. Дистанции, полученные при этом в некоторых случаях, достигают нижних пределов межвидовых различий.
Генетически определенный материал на данном этапе исследования позволил с уверенностью утверждать, что на Западном Кавказе только в пределах Адыгеи оба вида обитают симпатрично, а чаще и симбиотопично, где в более теплых и влажных условиях равнины и предгорий, богатых и разнообразных кормовых ресурсов лесостепного и пояса широколиственных лесов, доминирует кавказская лесная мышь. В поясе субальпийских лугов и в пределах верхней границы широколиственных лесов, где био- топы, представлены каменистыми россыпями, напротив, малая лесная мышь.
Вероятно, кавказская лесная мышь заняла равнину и предгорья Северо-Западного Кавказа значительно раньше, чем A. uralensis , о чем свидетельствует более высокая степень внутривидового генетического разнообразия, характер распространения, биотопическая приуроченность и численность A. ponticus в районе исследования.
Малый объем выборки по КЧР не позволяет на данном этапе однозначно судить о том, как далеко на восток проникает A. ponticus , для выяснения чего необходимо включить в анализ дополнительный материал из разных географических пунктов Карачаево-Черкесской Республики.
Авторы выражают огромную благодарность зав. кабинетом методов молекулярной диагностики, дбн М.В. Холодовой и кбн И.Г. Мещерскому за постоянную помощь при освоении методики и предоставленную возможность работы на базе кабинета методов молекулярной диагностики ИПЭЭ им. А.Н. Северцова РАН.
Работа выполнена при финансовой поддержке гранта РФФИ №15-04-03981 и Программы Фундаментальных исследований президиума РАН «Живая природа: современное состояние и проблемы развития».
Список литературы Генетическое разнообразие и географическое распределение гаплотипов митохондриальной ДНК криптических видов лесных мышей подрода Sylvaemus Западного Кавказа
- Балакирев, А.Е. К вопросу о таксономическом статусе формы ciscaucasicus и ее родственных отношениях с малой лесной мышью Sylvaemus uralensis по данным секвенирования гена цитохрома b мт ДНК/А.Е. Балакирев и др.//Генетика. -2007. -Т. 43. -№12. -С. 1651-1666.
- Богданов, А.С. Анализ генетической изменчивости и дифференциации малой лесной мыши Sylvaemus uralensis (Rodentia, Muridae) методом RAPD-PCR/А.С. Богданов, Д.М. Атопкин, Г.Н. Челомина//Известия РАН. Серия биол. -2009. -№3. -С. 276-292.
- Doiron, S. A comparative mitogenomic analysis of the potential adaptive value of Arctic charr mtDNA introgression in brook charr populations (Salvelinus fontinalis Mitchill)/S. Doiron, P.U. Blier, L. Bernatchez//Mol. Biol. Evol. 2002. -Vol. 19. -P. 1902-1909.
- Гащак, С. О видовом разнообразии мышей рода Sylvaemus в Украине/С. Гащак и др.//Раритетна терiофауна та її охорона. -Луганськ, 2008.-Вып. 9. -С. 80-92.
- Амбрамсон, Н.И. Филогеография: итоги, проблемы, перспективы/Н.И. Абрамсон//Вестник ВОГиС. 2007. -Т. II, -№2. -С. 307-331.
- Челомина, Г.Н. Филогенетические связи видов и внутривидовых форм лесных мышей рода Sylvaemus по данным частичного секвенирования гена цитохромa b мтДНК/Г.Н. Челомина, Д.М. Атопкин, А.С. Богданов//Генетика. -2007. -Т. 43. -№ 12. -С. 1651-1666.
- Hall, T.A. BioEdit: a user-friendly biological sequence aligment editor and analysis program for Windows 95/98/NT/T.A. Hall/Nucl. Asids. Symp. Ser. 1999. -V.41. -95-98.
- Tamura K. MEGA 4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0/K. Tamura et. al.//Molecular Biology and Evolution. -2007. -V. 24. -P. 1596-1599.
- Nei, M. Molecular Evolutionary Genetics./M. Nei//N.Y.: Columbia Univ. Press, 1987. -512 p.
- Bandelt, H.J. Median-Joining Networks for Inferring Intraspecific Phylogenies/H.J. Bandelt, P. Forster, A. Rohl//Mol. Biol. Evol. 1999. -V. 16. -№1. -Р. 37-48.
- Kotlik, P. A northern glacial refugium for bank voles (Clethrionomys glareolus)/P. Kotlik, V. Deffontaine, S. Mascheretti//PNAS. 2006. -V.103. -№ 40. -P. 14860-14864.
- Амбрамсон, Н.И. Гетическая изменчивость и филогеография рыжей полевки (Clethrionomys glareolus, Arvicolinae, Rodentia) на территории России с анализом зоны интрогрессии мтДНК близкородственного вида -красной полевки (Cl. rutilus)/Н.И. Абрамсон, Е.Н. Родченкова, Ю.А. Костыгов//Генетика. -2009. -Т. 45. -№5. -С. 610-623.
- Челомина, Г.Н. Молекулярно-генетические свидетельства глубокого филогенетического разрыва между европейской и азиатской расами малой лесной мыши по данным изменчивости гена цитохрома b мтДНК/Г.Н. Челомина, Д.М. Атопкин//Молекулярная биология. -2010. -Т. 44. -№5. -С.792-803.
- Челомин,а Г.Н. Лесные и полевые мыши: молекулярно-генетические аспекты эволюции и систематики/Г.Н. Челомина//Владивосток: Дальнаука, 2005. -205 с.
- Martin, Y. Molecular phylogeny of European muroid rodents based on complete cytochrom b sequences/Y. Martin аt. al.//Mol. Phylogenetics and Evolution. -2000. -V. 16. -P. 37-47.
- Окулова, Н.М. Биотопическое размещение мелких млекопитающих в горах и предгорьях Краснодарского края и Республики Адыгея/Н.М. Окулова и др.//Млекопитающие горных территорий. Материалы международной конференции. М.: КМК. 2007. -С. 230-233.
- Avise, J.C. Phylogeography: the history and formation of species/J.C. Avise//Cambridge; London: Harvard Univ. press, 2000. -447 p.
- Сорокин, П.А. Изолированность популяций монгольского дзерена, Procapra gutturosa (Artiodactyla, Bovidae), в прошлом: анализ фрагментов митохондриальной ДНК с разной скоростью мутирования/П.А. Сорокин, М.В. Холодова//Доклады РАН. -2006. -Т. 409(2). -С. 1-2.
- Переверзева, В.В. Генетическая структура популяций красной полевки Myodes (Clethrionomys) Rutilus Pallas, 1779 Северного Приохотья по данным об изменчивости нуклеотидных последовательностей гена цитохрома b митохондриальной ДНК/В.В. Переверзева, А.А. Примак, Е.А. Дубинин//Вавилов. журн. генет. и селекции. -2013. -Т. 17. -№ 3. -С. 435-443.
- Богданов, А.С. Анализ генетической изменчивости и дифференциации лесных мышей рода Sylvaemus по результатам секвенирования фрагмента гена первой субъединицы цитохромоксидазы/А.С. Богданов и др.//Генетика. -2012. -Т. 48. -№2. -С. 211-224.
- Окулова, Н.М. Видовое разнообразие, размещение и численность мелких млекопитающих южных территорий Краснодарского края и республики Адыгея/Н.М. Окулова и др.//Млекопитающие горных территорий. Материалы международной конференции. М.: КМК. -2005. -С. 118-121.
