Геногеографическая изменчивость гемоглобина у красной полевки (Myodes rutilus Pallas 1779) юга Дальнего Востока России
 юга Дальнего Востока России")
Автор: Фрисман Л.В., Картавцева И.В., Павленко М.В., Капитонова Л.В.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Фаунистика. Эволюционная генетика
Статья в выпуске: 3 т.19, 2016 года.
Бесплатный доступ
В статье представлены результаты электрофоретического исследования гемоглобина в популяциях красной полевки (Myodes rutilus) юга Дальнего Востока России. Исследование геногеографической изменчивости показало наличие пространственной дифференциации. Были обнаружены две географические группы. Популяции островов Большой Шантар, Сахалин, Хоккайдо и соседних территорий Тихоокеанского побережья принадлежат к одной и той же группе. К этой же группе относятся популяции Восточного Сихотэ-Алиня. Другая группа объединяет материковые популяции Приамурья и прилежащих территорий. Показана парапатричность ареалов этих групп при узкой зоне наложения их ареалов.
Красная полевка, гемоглобин, ареал, юг дальнего востока России, генетическая дифференциация
Короткий адрес: https://sciup.org/14328972
IDR: 14328972 | УДК: 575.22:
Gene-geographical variation of hemoglobins in red-backed voles (Myodes rutilus Pallas 1779) from the south of the russian far east
The article presents the results of electrophoretical analysis of hemoglobin in northern red-backed voles (Myodes rutilus) from the south-eastern part of Russia. The authors studied a gene-geographic variation of the species and determined its differentiations. It was identified two geographical groups. The Myodes rutilus populations of Big Shantar Island, Sakhalin Island, Hokkaido Island (Japan), and populations of red voles from neighboring Pacific coast belong to the same group. East populations of the Sikhote Alin Mountains also belong to this group. Another group includes continental populations of the Amur region and adjacent areas. The two groups are effectively parapatric, their ranges only narrovly overlapping.
Текст научной статьи Геногеографическая изменчивость гемоглобина у красной полевки (Myodes rutilus Pallas 1779) юга Дальнего Востока России
Красная полевка (Moydes rutilus Pallas, 1779) широко распространена по территории Голаркти-ки: от Швеции и Норвегии в Евразии до побережья Гудзонова залива в Северной Америке. Она заселяет темнохвойные, смешанные и хвойно-широколиственные леса, обитает в лесных участках лесо-, луго- и тундростепей, в горах до верхних пределов леса, а также в пойменных лесах степной зоны. На севере Евразии ареал заходит в лесотундру и южную тундру, на юге – в лесостепные участки Казахстана и Западной Сибири, горные леса Северной Монголии, Северо-Восточного Китая, полуострова Корея и Северной Японии. Для территории бывшего СССР известно до 20 подвидов. Из них не более 7–8 представляют реальные таксономические категории (Громов, Ербаева, 1995). В дальневосточной части ареала систематики выделяют четыре географических подвида (Громов, Ербаева, 1995; Костенко, 2000). M. r. jacuten-sis Vinogradov, 1927 объединяет красных полевок центральных районов Якутии, к западу и югу от Верхоянского хребта и верховьев р. Мая. Ареал M. r. jochelsoni G. Allen, 1903 охватывает территорию от зоны типичной тундры и лесотундры на севере Магаданской области до зоны типичной лиственничной тайги на севере Хабаровского края и Амурской области, включает п-ов Камчатка, о-ва Карагинский, Парамушир. Обитатели хвой- но-широколиственных и елово-пихтовых лесов Амурской области, Хабаровского и Приморского краев, Шантарских островов, северо-востока Китая и полуострова Корея объединены в подвид M. r. amurensis Schrenck, 1859. В подвид M. r. micado Tokuda, 1935 выделены красные полевки о-ва Сахалин и острова Хоккайдо (Япония).
Известно, что привлечение генетических характеристик не только оказывается полезным для рассмотрения таксономической подразделенности и эволюционной истории вида, но зачастую является решающим инструментом в спорных воспросах систематики. Исследование электрофоретической изменчивости 13 ферментов и 3 неэнзимных белков, контролируемых 25 локусами (Фрисман и др., 2002), показало генетическую близость красной полевки островов Сахалин, Хоккайдо ( M. r. mikado ) как между собой, так и с популяцией западного побережья Татарского пролива ( M. r. amurensis ) и их значимое отличие от популяций юга Магаданской области и полуострова Камчатка ( M. r. jochelosni ). Наиболее значимая геногеографическая изменчивость (смена основного аллеля между группами популяций соседствующих географических регионов) была обнаружена по двум маркерам. Это маркеры гемоглобин (Hb) и глюкозо-6-фосфатдегидрогеназа (G6PD). Выборки Магаданской области и п-ова
Камчатка были мономорфны по Hb-а. В качестве единственного или основного аллеля в этих выборках был G6PD-A. В выборках материкового побережья Татарского пролива, о-вов Сахалин и Хоккайдо были обнаружены Hb-b и G6PD-В. Иными словами, наблюдалось разделение всех исследованных популяций красной полевки на две географические группы, которые мы обозначили как «северная» и «южная». Наиболее интересные и информативные картины геногеографии были обнаружены по гемоглобину (Hb). Именно гемоглобин был выбран нами для последующего более детального исследования геногеографии красной полевки юга Дальнего Востока России. Идентичность Hb-а единственному варианту гемоглобина M. rufocanus позволила привлечь литературные данные и тем самым расширить регион исследования геногеографии этого белка. На основании литературных данных в «северную» группу кроме красных полевок полуострова Камчатка и юга Магаданской области были отнесены представители этого вида Чукотки, Аляски и юго-запада Магаданской области, у которых был обнаружен только вариант, идентичный гемоглобину красно-серой полевки (Nadler et al., 1978; Курышев, 1990). В «южную» группу мономорфных по Hb-b популяций, кроме красных полевок о-вов Сахалин, Хоккайдо и западного побережья Татарского пролива, вошли обитатели о-ва Большой Шантар и левобережья Амурского лимана (Коробицына и др., 1983). Было высказано предположение, что обнаруженная картина геногеографической изменчивости отражает достаточно долгую независимую историю «северной» и «южной» гемоглобиновых форм, а сходство популяций островов Сахалин, Хоккайдо и популяций прибрежных материковых территорий по гемоглобиновым спектрам возникло до отделения данных островов от материка. Возникает вопрос: как далеко на запад может быть распространена «южная» форма? Цель настоящей работы – анализ внутривидовой дифференциации и истории становления ареала красной полевки юга Дальнего Востока. Задача – исследование встречаемости вариантов гемоглобина в популяциях красной полевки территории Приамурья, Приморья и прилежащих регионов.
Материалы и методы
Материал настоящего исследования, включая ранее опубликованные нами данные по 67 животным из 7 суммарных географических выборок (Фрисман и др., 2002), объединяет 270 экземпляров красной полевки из 41-го местонахождения на территории Приамурья, Приморья, островов
Большой Шантар, Сахалин, Хоккайдо (Япония) и единичных выборок Новосибирской области, Бурятии, Забайкалья и Якутии (рис. 1). Места, годы сбора и численность выборок представлены в табл. Отбор крови проводился в лабораторных либо в полевых условиях. В последнем случае собранные образцы доставлялись в лабораторию в охлажденном виде. Был проведен электрофорез гемолизата в крахмальном геле с последующим окрашиванием на общий белок по методу Пастера с соавторами (Pasteur et al., 1988).
Практически по всей дальневосточной территории красная полевка может встречаться совместно с красно-серой. Видовая диагностика была проведена нами на основе как морфологических признаков (окрас, ширина резцов, зубы) и диагностических для данных видов трех электрофоретических маркеров – изоцитратдегидрогене-за-1(IDH-1), изоцитратдегидрогеназа-2 (IDH-2), глутамата-оксалоацетат-трансаминаза (GOT-1), межвидовые различия аллельного состава которых были описаны нами ранее (Фрисман и др., 2002). Следует отметить, что гемоглобин у всех рассматриваемых нами красно-серых полевок (более 100 экз.) был мономорфен и идентичен Hb-а.
Результаты и обсуждение
В результате электрофоретического разделения и последующего окрашивания гемолизата гемоглобин у красных полевок выявляется в анодной области геля в виде двух зон различной яркости. Наиболее интенсивно окрашенная зона обладает большей подвижностью. Визуальные различия гемоглобиновых вариантов (электроморф) представлены различной анодной подвижностью двузонных комплексов (рис. 2). Электрофоретический анализ 270 красных полевок из 41 географической локальности позволил идентифицировать 3 гемоглобиновых варианта. Наиболее часто представлены Hb-а и Hb-b (табл.). Кроме того, у трех полевок их популяций Среднего Приамурья был выявлен Hb-с, обладающий промежуточной электрофоретической подвижностью по отношению к двум вышеуказанным вариантам. Распространение Hb-а и Hb-b подтверждает полученную ранее нами картину геногеографии красной полевки в дальневосточной части ее ареала, а именно существование на этой территории двух географических групп популяций, различающихся по набору гемоглобиновых вариантов. Геногеографическая изменчивость характеризуется единообразием внутри групп и сменой аллельного состава на стыке их ареалов. Только Hb-а был обнаружен нами в выборках Амурской области (точки 8-12, здесь
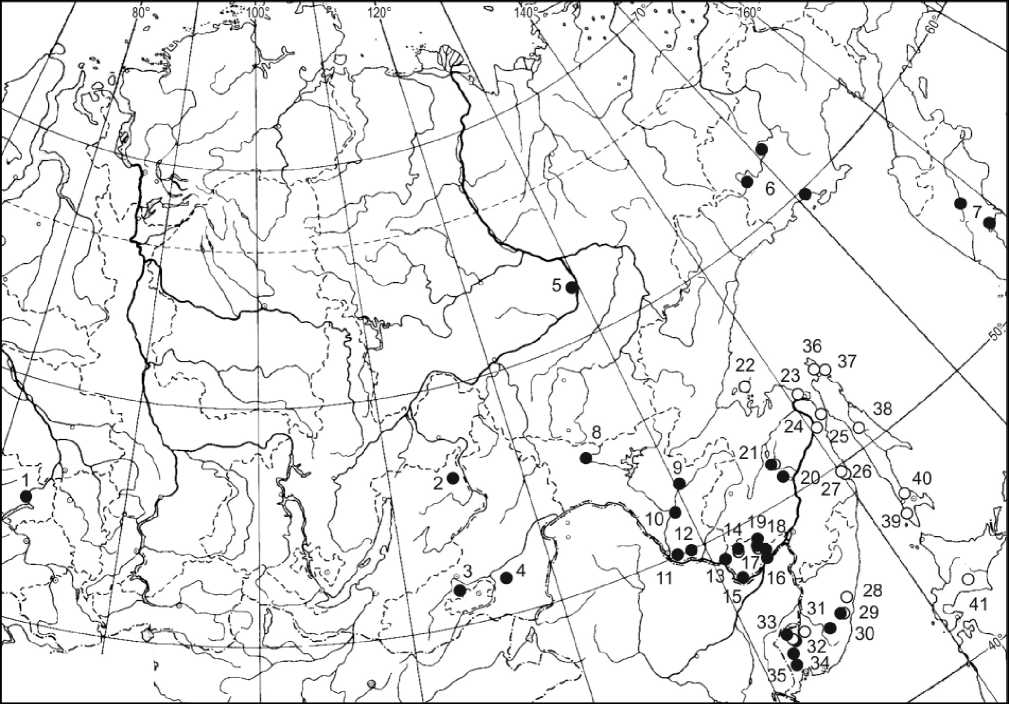
Рис. 1 Места отлова красной полевки Myodes rutilus и встречаемость вариантов гемоглобина в популяциях. Символы соответствуют вариантам гемоглобина: черный кружок – Hb-a, белый кружок – Нb-b, полосатый кружок – Hb-c
Fig. 1. Capture places of northern red-backed vole Myodes rutilus and geographical distribution of Hb variants in populations. The symbols correspond to hemoglobin variants: black circle – Hb-a, white circle – Hb-b, striped circle – Hb-c и далее – табл.) и Еврейской автономной области (точки 13-19), а также в единичных выборках Западной Сибири, Забайкалья и Якутска (точки 1-5). Ранее этот вариант был найден в исследованных популяциях Магаданской области и п-ова Камчатка (суммарные точки 6-7) (Фрисман и др., 2002), Омолона, Чукотки и Аляски (Курышев, 1978; Nadler, 1978). В «южную» группу, мономорфную по Hb-b, кроме красных полевок островов Сахалин, Хоккайдо (точки 36-41) и западного побережья Татарского пролива (точка 24 ), входят также красные полевки о-ва Большой Шантар, (точка 22), право- и левобережья нижнего Амура (точки 2325) и восточных склонов Сихотэ-Алиня (точки 26-28). Данная группа заселяет юго-восточную оконечность азиатского ареала красной полевки и, по-видимому, представляет собой «юго-восточную» линию эволюционной истории вида. Между популяциями «юго-восточной» линии с одной сто- роны и популяциями остальных дальневосточных территорий ареала обнаружены географические точки, где совместно представлены гемоглобиновые маркеры, характеризующие эти различающиеся группы популяций. Такими местонахождениями являются в Нижнем Приамурье – окрестности озера Эворон в Эворон-Чукчагирской впадине (точка 21), а в Южном Приморье – юго-восточное побережье оз. Ханка и южные отроги хребта Сихотэ-Алинь в районе пос. Кавалерово (точки 30-31). Материал из этих районов позволил предположить существование гибридной зоны, протянувшейся вдоль западных отрогов Сихотэ-Алиня (Фрисман, Бородин, Фрисман, 2010).
Ископаемые остатки лесных полевок известны с позднего плиоцена (Громов и др., 1963). Общность лево- и правобережных популяций нижнего Амура по Hb-b не позволяет рассматривать Амур в его современном русле в качестве
Таблица
Данные электрофоретического исследования гемоглобина Myodes rutilus из различных районов
Table
Data on the electrophoretical research of hemoglobinin Myodes rutilus from different regions
|
№ пп |
Точки отлова |
Годы сбора |
Объем выборки |
Количество носителей варианта Hb |
Источник |
||
|
Hb-a |
Hb-b |
Hb-c |
|||||
|
Myodes rutilus rutilus Pallas, 1779 |
|||||||
|
1. |
г. Новосибирск |
1999 |
2 |
2 \ |
Наши данные |
||
|
M. r. baicalensis Ognev, 1924 |
|||||||
|
2. |
Республика Бурятия, Муйский р-н, пос. Таксимо |
2007 |
2 |
2 |
Наши данные |
||
|
Забайкальский край |
|||||||
|
3. |
Дульдургинский район, с. Бальзино |
2010 |
3 |
3 |
Наши данные |
||
|
4. |
Балейский р-н, пос. Подойницыно |
2007 |
5 |
5 |
Наши данные |
||
|
M. r. jacutensis Vinogradov, 1927 |
|||||||
|
5. |
г. Якутск |
1999 |
2 |
2 \ |
Наши данные |
||
|
M. r. jochelsoni J. Allen, 1903 |
|||||||
|
6. |
Магаданская обл., г. Магадан, пос. Дэбин, пос. Стоковое |
8 |
8 |
Фрисман и др., 2002 |
|||
|
7. |
П-ов Камчатка, г. Милково, г. Петропавловск-Камчатский |
9 |
9 |
Фрисман и др., 2002 |
|||
|
М. r . amurensis Shrenk, 1859 |
|||||||
|
Амурская область |
|||||||
|
8. |
г. Тында |
2001 |
1 |
1 |
Наши данные |
||
|
9. |
Мазановский р-н, с. Новокиевский Увал |
2010 |
2 |
2 |
Наши данные |
||
|
10. |
Заповедник Норский |
2006 |
7 |
7 |
Наши данные |
||
|
11. |
Михайловский р-н, с. Михайловка, с. Арсеньевка |
2005 |
2 |
2 |
Наши данные |
||
|
12. |
Тамбовский р-н, с. Тамбовка |
2005 |
1 |
1 |
Наши данные |
||
|
Еврейская автономная область |
|||||||
|
13. |
Облученский р-н, с. Пашково |
2010 |
1 |
1 |
Наши данные |
||
|
14. |
Облученский р-н, с. Кульдур, пос. Известковый |
2010 |
5 |
4 |
1 |
Наши данные |
|
|
15. |
Октябрьский р-н, с. Амурзет, с. Садовое |
2010 |
3 |
3 |
Наши данные |
||
|
16. |
Ленинский р-н, с. Ленинское |
2007, 2012 |
12 |
12 |
Наши данные |
||
|
17. |
г. Биробиджан, с. Валдгейм |
2004– 2012 |
15 |
13 |
2 |
Наши данные |
|
|
18. |
Биробиджанский р-н, с. Дубовое |
2012 |
4 |
4 |
Наши данные |
||
|
19. |
Заповедник Бастак (с. Кирга, р. Глинянка, квадрат 100) |
2010– 2012 |
26 |
26 |
Наши данные |
||
|
Хабаровский край |
|||||||
|
20. |
Солнечный р-н, пос. Солнечный |
2002 |
1 |
1 |
Наши данные |
||
|
21. |
Солнечный р-н, с. Эворон |
2002 |
6 |
5 |
1 |
Наши данные |
|
|
22. |
о-в Большой Шантар |
1983 |
21 |
21 |
Коробицына и др, 1983 |
||
|
23. |
Николаевский р-н, с. Красное |
1983 |
11 |
11 |
Коробицына и др, 1983 |
||
|
2012 |
6 |
6 |
Наши данные |
||||
|
24. |
Николаевский р-н, пос. Лазарев |
2001 |
7 |
7 |
Фрисман и др, 2002 |
||
|
25. |
Богородский р-н, с. Богородское |
2002, 2012 |
10 |
10 |
Наши данные |
||
Продолжение таблицы
Данные электрофоретического исследования гемоглобина Myodes rutilus из различных районов
The rest of the table
Data on the electrophoretical research of hemoglobinin Myodes rutilus from different regions
Для красной полевки, в том числе и на дальневосточной части ее ареала, характерна сложная комбинация разнонаправленной географической изменчивости морфологических характеристик, что в значительной степени осложняет внутривидовую таксономическую интерпретацию. В силу этого выделение систематиками дальневосточных подвидов осуществлено в основном по географическому принципу с привлечением в некоторых случаях вариаций окраски. Геногеография ге-
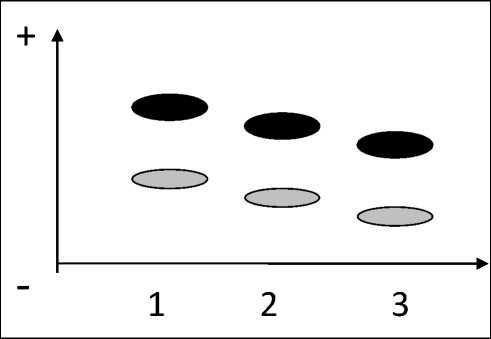
Рис. 2 Электрофоретические варианты гемоглобина красной полевки Myodes rutilus: 1) Hb-b, 2) Hb-c, 3) Hb-a
Fig. 2. Electrophoretical variants of hemoglobin in northern red-backed vole Myodes rutilus:
1) Hb-b, 2) Hb-c, 3) Hb-a моглобина подтверждает родство красных полевок о-вов Хоккайдо, Сахалин, Б. Шантар, Амурского лимана и восточных склонов Сихотэ-Алиня и показывает комплексный характер М. r. amuren-sis. В состав этого подвида, кроме представителей «юго-восточной» группы (обитатели острова Б. Шантар, Амурского лимана и востока Приморья), включены популяции, принадлежащие к «северной» гемоглобиновой форме, а также гибридные популяции. Комплексный состав М. r. amu-rensis обнаруживается и при внимательном рассмотрении построенной Костенко (Костенко, 2000) дендрограммы сходства 11 дальневосточных выборок М. rutilus, рассчитанной на основе 10 морфологических показателей тела и черепа. Если выборки М. r. jacutensis и М. r. mikado достаточно хорошо отличаются друг от друга и от остальных дальневосточных подвидов, а выборки М. r. jochelsoni создают достаточно компактные группы, то локальные выборки M. r. amurensis либо представляют собой самостоятельные линии, либо объединяются в единые линии с M. r. jo-chelsoni.
Аллозимные характеристики определяются ядерным геномом и их дифференциация в общих чертах согласуется с морфологической (морфотипической) изменчивостью. Иная картина изменчивости обнаружена при анализе митохондриального генома красной полевки. Так, Абрамсон и Бодровым (Abramson, Bodrov, 2008) по результатам секвенирования гена цитохрома b (cytb) была предложена классификация филогрупп красной полевки, включающая четыре клады – западную, восточную, берингийскую и японскую. Отмечено, что границы между ними не вполне ясны и практически не совпадают с традиционно выделяемыми подвидами (Павлинов, Лисовский, 2012). М. Иваса с соавторами (Iwasa et al., 2009) объединяют в единую группу полевок п-ова Камчатки и о-ва Сахалина и выделяют в самостоятельную филетическую линию полевок о-ва Хоккайдо. Сложно структурированная картина дифференциации красной полевки по митохондриальным и ядерным генам показана и другими авторами (Kohli et al., 2015). На основе анализа изменчивости гена cytb в азиатской части ареала красной полевки выявляются три основные группировки – западная (Урал, Западная Сибирь), центральная (Восточная Сибирь, Забайкалье, Приамурье, Колыма, Чукотка) и восточная, включающая, в свою очередь, две субклады – полевок Сахалина и Хоккайдо, а полевки Камчатки относятся к североамериканской (берингийской) группировке, в то время как по ядерному гену MLR регион Колымы определятся как зона контакта между азиатскими и американскими группами. Митохондриальное единство континентальных популяций Северного Приохотья, относящихся по первой классификации к восточной кладе, при включении отдельных островных популяций Охотоморья (остров Маты-киль) в общую кладу с красными полевками Камчатки и Сахалина, показана и в работах коллег из ИБПС (Магадан) (Переверзева и др., 2011, 2013а, 2013б). В отличие от картины дифференциации по маркерам митохондриального генома, популяции Аляски и Камчатки, относящиеся к берингий-ской кладе, по вариантам гемоглобина относятся к одной группе с полевками Магаданской области, Якутии, Забайкалья, Среднего Амура. Особо отметим, что, хотя картина геногеографии гемоглобинов у красной полевки отличается от подразделенности, выявленной по другим генетическим маркерам, просматривается общий тренд – дифференциация островных популяций, в частности, Сахалина и Хокайдо.
Строгое сравнение полученных нами результатов с литературными данными затруднительно, поскольку в работах других авторов по генетической дифференциации красной полевки материал с материковой части юга Дальнего Востока России представлен единичными выборками, а прибрежные юго-восточные территории не рассматривались вообще.
Итак, проведенное исследование географической изменчивости встречаемости электрофо- ретических вариантов гемоглобина в популяциях Приамурья, Приморья и прилежащих территорий показало существование в этих регионах двух па-рапатричных групп популяций, различающихся набором гемоглобиновых вариантов. Юг Дальнего Востока России можно определить как зону контакта двух генетических «гемоглобиновых» линий красной полевки. Вопрос о путях и времени формирования этой зоны пока остается открытым.
Мы выражаем свою глубокую признательность Кире Всеволодовне Коробицыной (1936– 2011) и Владимиру Павловичу Кораблеву (1951– 2014), принимавшим активное участие в сборе материала и предварительном обсуждении результатов исследования.
Работа выполнена при частичной финансовой поддержке грантов РФФИ: 09-04-00146-а, № 12-04-98540-р_восток_а, грантов Конкурса ДВО РАН ДВО: №11-III-Д-06-025, № 12-I-П30-14, гранта 15-I-6-049 программы фундаментальных научных исследований «Дальний Восток».
Список литературы Геногеографическая изменчивость гемоглобина у красной полевки (Myodes rutilus Pallas 1779) юга Дальнего Востока России
- Артеменко Т.В., Сорокин А.П. Условия формирования и эволюция бассейна Амура//География и природные ресурсы. 2009. № 4. С. 106-111
- Громов И.М., Гуреев А.А., Новиков Г.А., Соколов И.И., Стрелков П.П., Чапский К.К. Млекопитающие фауны СССР. Ч. 1. М.; Л.: Изд-во АН СССР, 1963. 638 с
- Громов И.М., Ербаева М.А. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб., 1995. 522 с
- Гросвальд М.Г. Оледенение Русского Севера и Северо-Востока в эпоху последнего великого похолодания//Материалы гляциологических исследований. Вып. 106. М.: Наука, 2009. С. 1-152
- Коробицына К.В., Фрисман Л.В., Якименко Л.В. Кариологическое и биохимическое исследование лесных полевок Дальнего Востока//Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983. С. 92-93
- Костенко В.А. Грызуны (Rodentia) Дальнего Востока России. Владивосток: Дальнаука, 2000. 209 с
- Курышев С.В. Растворимые белки сыворотки крови и гемоглобины красной и красно-серой полевок//Генетика. 1990. Т. 36, № 10. С. 1822-1828
- Павлинов И.Я., Лисовский А.А. Млекопитающие России: систематико-географический справочник. М.: Т-во научн. изданий КМК, 2012. 604 с
- Переверзева В.В., Засыпкин М.Ю., Соловенчук Л.Л. и др. Изменчивость гена цитохрома b митохондриальной ДНКвпопуляции красной полевки Сlethrionomys rutilus Pallas, 1779 поймы среднего течения реки Колымы//Известия РАН. Серия биологическая. 2011. № 3. С. 283-288
- Переверзева В.В., Примак А.А., Дубинин Е.А. Генетическая структура популяций красной полевки Myodes (=Clethrionomys) rutilus Pallas, 1779 Северного Приохотья по данным об изменчивости нуклеотидных последовательностей гена цитохрома b митохондриальной ДНК//Вавиловский журнал генетики и селекции. 2013. Т. 17, № 3. С. 435-443
- Переверзева В.В., Примак А.А., Дубинин Е.А. Филогенетисческие отношения популяций красной полевки Myodes (=Clethrionomys) rutilus Pallas, 1779 Северного Приохотья и колымского региона//Вавиловский журнал генетики и селекции. 2013. Т. 17, № 3. С. 444-451
- Сорокин Ф.П., Махинов А.Н., Воронов Б.А., Сорокина А.Т., Артеменко Т.В. Эволюция бассейна Амура в мезозое-кайнозое и ее отражение в современной динамике рельефа//Вестник ДВО РАН. 2010. № 3. С. 82-80
- Фрисман Л.В., Бородин П.В., Фрисман Е.Я. Эволюционная динамика гибридных зон млекопитающих//Региональные проблемы. 2010. Т. 13, № 2. С. 56-62
- Фрисман Л.В., Картавцева И.В, Павленко М.В., Костенко В.А., Сузуки Н., Иваса М., Накато К., Чернявский Ф.Б. Геногеографическая изменчивость и генетическая дифференциация лесных полевок рода Clethrionomys (Rodentia, Cricetidae) Приохотья//Генетика. 2002. Т. 38, № 5. С. 655-664
- Чемеков Ю.Ф. История развития речной сети в бассейне реки Амур//Известия АН СССР. Серия географическая. 1964. № 1. С. 81-93
- Abramson N.I., Bodrov S.Yu. Genetic differentiation and phylogeography of Сlethrionomys rutilus Pallas 1811 inferred from variation of mitochondrial cytochrome b gene//11th International conference «Rodens et Spatium» on Rodent Biology. Myshkin, Russia, July 24-28. Moscow: VTO RAS, 2008. P. 64
- Iwasa M.A., Kostenko V.A., Frisman L.V., Kartavtseva I.V. Phylogeography of the root vole Microtus oeconomus in Russian Far East: A special reference to comparison between Holarctic and Palaearctic voles//Mammal study. 2009. Vol. 34. P. 123-130
- Kohli B.A. Fedorov V.B., Waltari E., Cook J.A. Phylogeography of a Holarctic rodent (Myodes rutilus): testing high-latitude biogeographical hypotheses and the dynamics of range shifts//Journal of Biogeography. 2015. Vol. 42. P. 377-389
- Nadler C.F., Zhurkevich N.M., Hoffman R.S. Biochemical relationships of the Holarctic vole genera Clethrionomys, Microtus and Arvicola (Rodentia, Arvicolinae)//Canad. J. Zool. 1978. Vol. 56, N 7. P. 1564-1575
- Pasteur N., Pasteur G., Bonchomme F. et al. Practical isozyme genetics. Hebsted Press: a division of Jone Wiley and sons. New-York Chichested Brisbane Toronto, 1988. 215 p
