Геобиологические сети Хартмана и Карри в испытательных культурах ели сибирской

Автор: Рогозин Михаил Владимирович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 2, 2011 года.
Бесплатный доступ
Исследовано влияние на рост растений элементов геобиологических энергетических сетей Хартмана и Карри. В усиленных узлах этих сетей с диаметрами 0.55-2.0 м деревья в 21-летнем возрасте снижали рост в высоту на 13.4-14%. Зоны сильного депрессивного влияния данных сетей занимают до 1.9%, среднего и слабого влияния - примерно 10-11% площади. Поэтому необходим поиск и исключение таких зон в полевых опытах с выращиванием растений на небольших делянках, а также в лабораторных опытах.
Лесные культуры, ель сибирская, потомство, геобиологические сети
Короткий адрес: https://sciup.org/147204551
IDR: 147204551 | УДК: 630.5(075),630
Geobiological network of Hartman and Curry in test cultures spruce Siberian
The effect on plant growth elements of geobiological energy networks Hartman and Curry. In reinforced nodes of the networks with diameters of 0.55-2.0 m trees in 21 years of age reduced the growth in height of a 13.4-14%. The zone of strong depressive influence of these networks take up to 1.9%, medium and low impact - about 10-11% of the area. It is necessary to search and deletion of such zones in field experiments with the cultivation of plants on small plots, as well as in laboratory experiments.
Текст научной статьи Геобиологические сети Хартмана и Карри в испытательных культурах ели сибирской
Краткие сведения о геобиологических сетях
Геоактивные энергетические структуры Земли – геобиологические сети Хартмана и Карри являются одними из множества геофизических полей планеты Земля. В монографии «Экологический мониторинг в геофизике» (Поносов и др., 2009) даны объяснения механизмов возникновения геоактивных (геопатогенных) зон с позиций геофизики. В кратком изложении эти механизмы выглядят следующим образом.
Особенности природных объектов и процессов (геологических, геофизических, геохимических и др.) и сопутствующих им геофизических полей вызывают неоднородности в литосфере Земли. Эти неоднородности вызывают устойчивые патологии в живых организмах, и такие места называют аномальными или патогенными зонами. Вопрос о них в середине 20 столетия привлек внимание широкого круга специалистов – медиков в первую очередь, а также биологов, геологов и геофизиков. Геопатогенные зоны можно объединить, по крайней мере, в два типа.
Первый тип связан преимущественно с нахождением токсичных химических элементов и повышенным выделением радона и других газов из недр Земли, распространением патогенных бактерий. Этот тип легче поддается изучению, поскольку очевиден механизм патогенеза – вещественный обмен между источником и объектом воздействия.
Второй тип связан преимущественно с локальными аномалиями геофизических полей. Механизм патогенного влияния здесь изучен слабее, хотя сам факт наличия на поверхности Земли таких аномалий сомнений не вызывает. Наиболее сложным в данном случае представляется установление природы физических полей и механизм передачи патогенного воздействия на живые организмы. В.А. Рудник (1996, 1999, по Поносову и др., 2009) связывает наличие геоактивных зон с неоднородностями геологического состава и строения земной коры, развитием активных разломов различной природы и приуроченных к ним карстовых полостей и геологических тел, обладающих аномальными, по отношению к окружению, физическими свойствами.
Наряду с этим существуют попытки иного рода, уводящие нас в область «чистой» биолокации, полей неизвестной природы, в область изучения стационарных и мигрирующих «энергетических сеток и узлов» (сети Альберта, Витмена, Карри, Манфреда, Стальчинского, Хартмана, Швейцера, Шульги и др. (по Поносову, 1993), являющиеся, по мнению экстрасенсов-биолокаторов, источниками геопатогенеза. Однако в силу отсутствия на сегодняшний день геологического обоснования и надежного инструментального обнаружения подобного рода геопатогенных зон, их следует рассматривать как гипотетические и нуждающиеся в дальнейшем изучении. Поскольку геоактивные (геопатогенные) зоны могут оказывать прямое или опосредованное (через разрушения и аварии на различного рода объектах) негативное воздействие на биоту, их следует относить к местам повышенного экологического риска (Поносов и др., 2009).
В 1996 г. разработан переносной прибор для обнаружения геофизических аномалий, геопатогенных зон, сетей Хартмана и Карри, подземных водяных жил и других объектов. Прибор предназначен для измерения электромагнитной составляющей геомагнитного поля Земли, фиксирует «ауру» человека и ее искажения, связанные с патологическими изменениями в организме. Прибор разработан в
Влияние сетей Хартмана и Карри на деревья
Отрицательное влияние геобиологических сетей (ГБС) Хартмана и Карри на здоровье человека известно с 1970-х гг. и используется в настоящее время при диагностике геопатогенных зон в жилище человека. Менее изучено их влияние на растения, хотя знания о «плохих» местах рекомендуется использовать при посадке садовых деревьев и кустарников.
Наши наблюдения за растениями в питомниках и лесных насаждениях показали, что влияние данных ГБС обнаруживается глазомерно чаще всего в местах пересечения силовых линий этих сетей, называемых узлами. Узлы в плане имеют форму круга и бывают простые и усиленные. Диаметры обнаруживаемых нами узлов в сети Хартмана составляли: простых 0.3 м и усиленных 0.55 м; в сети Карри простые узлы имели диаметр 0.5 м и усиленные имели размеры 1.1 и 2.0 м.
Из множества зафиксированных нами аномалий развития деревьев разных пород на таких усиленных узлах упомянутых сетей в качестве примера приводим фото аллеи лиственниц Сукачева и группу растений ели колючей на территории учебного ботанического сада Пермского госуниверситета

Рис. 1. Ель колючая на узле ГБС Карри диаметром 1.1 м
(рис. 1 и 2). Липа мелколистная на нахождение в таких узлах этих сетей реагирует усыханием вершины и образованием двух вершин в виде перевернутой буквы «П» (рис. 3), а кедр сибирский – мно-говершинностью (рис. 4). Реакция взрослых деревьев дуба и березы была схожей с последним примером, но эти деревья обнаруживали признаки де-

Рис. 2. Лиственницы на полосе и узлах ГБС Хартмана искривлены и уклоняются от центра ряда

Рис. 3. Липа мелколистная на двойной зоне депрессии – совмещенных усиленных узлах сетей Хартмана и Карри прессии роста кроны. На основании около 60 случаев подобных наблюдений мы пришли к предварительному выводу о том, что если в естественных или искусственных насаждениях обнаруживались деревья с аномальной формой кроны, то основание ствола таких уродливых и искривленных растений попадало чаще всего в зону влияния усиленных узлов одной или сразу обеих сетей и только иногда – в зоны влияния простых узлов этих сетей. Был сделан также и явно обнаруживаемый вывод: растения стремятся уйти из зоны влияния узлов и силовых линий ГБС: вершины стволов отклоняются, разветвляются, либо отмирают и замещаются двумя и более вершинами. Ярко выраженные отличия в формы кроны являются результатом реакции растений на неблагоприятное воздействие этих сетей.

Рис. 4. Кедр сибирский на узле сети Карри диаметром 1.1 м в пос. Орел Березниковского района
Растения описанного облика встречаются в лесах и парках не так уж и редко. Мы решили выяснить повреждающее влияние сетей Хартмана и Карри более детально. Исследование было интересно с позиции их влияния на полевые опыты и возможностей исключения из выборки растений, попавших в депрессивные зоны.
Влияние ГБС на деревья в культурах ели
Полевой опыт представлен испытательными культурами ели, созданными в 1991 г. на среднесуглинистой почве на площади 11 га в кв. 41 Ильинского лесничества в Пермском крае. Участок расположен в 1.0 км на восток от автодороги Пермь-Ильинск, на расстоянии 72.7 км по ней от г. Перми, в 4 км на север от пос. Васильевское, в начале восточного склона плоского водораздела возвышенности с отметками высот 225–250 м.
Целью создания культур было получение оценок по быстроте роста у потомства 525 материнских деревьев и их селекция. Было высажено 22.4 тыс. растений, распределенных в 6 повторностей. Посадка проведена 4-летними саженцами на раскорчеванную вырубку по схеме 2.5×1.0 м (Рогозин,
2008). Изучением ГБС мы тогда не занимались и при посадке их не учитывали.
В 2007 г. в возрасте растений 21 год мы измеряли их высоту и нанесли деревья на план с точностью ± 0.2 м на одном из участков размером 100 × 62 м. Координаты левого нижнего угла участка: N 58˚27΄20.9΄΄Е 55˚50΄17.8΄΄. Спустя 2 года, 30 и 31 июля 2009 г., на этот план были нанесены некоторые элементы ГБС Хартмана и Карри. Биолокация осуществлялась одним оператором с помощью маятника и соблюдением определенных правил таких работ (Поносов, 1993). Двумя угловыми засечками определялось положение усиленных узлов сети Хартмана диаметром 0.55 м (61 шт.), а на следующий день – усиленных узлов сети Карри с диаметром 1.1 м (48 шт.) и с диаметром 2.0 м (19 шт.). Определялось вращение энергии в периферии узла – против (–) или по часовой стрелке (+) и связи с соседними узлами. Связи существовали только между узлами с противоположными знаками, и это служило проверкой правильности построения сети.
Площадь усиленных узлов составила: в сети Хартмана 14.49 м2 или 0.23%, а в сети Карри – 105.3 м2 или 1.69% площади участка. Усиленные узлы были соединены сетью полос шириной 0.3 м: в сети Хартмана протяжением 1106 м и площадью 332 м2 или 5.35%, а в сети Карри протяжением 1142 м и площадью 343 м2 или 5.53% площади участка. Общая площадь ГБС, на которой мы изучали их влияние на растения, составила: сети Хартмана 5.58% и сети Карри 7.22% площади участка.
На рис. 5 и 6 показано расположение растений и фрагменты структуры ГБС: усиленные узлы и связь между ними в виде линий (полос).
Обычные узлы, как более слабые, мы не наносили на схему, но в правой верхней части рис. 5 частично показана полная структура сети Карри.
Общий рисунок сетей сильно отличается от теоретических моделей этих сетей, приводимых в литературе. Практически нет одинаковых по форме ячеек, а их площади различаются до 8–10 раз. Кроме того, имеются по крайней мере два места, где ячейки приобрели явно искаженную форму (в левой части участка вблизи границы и чуть ниже и правее центра). Можно предполагать, что искажения структуры сетей вызваны влиянием более сильных геофизических полей. По-видимому, здесь эти поля имеют возмущения, вызываемые неоднородностями состава и строения земной коры, напряжением в пластах, подземными водными потоками и другими особенностями литосферы (Поносов и др., 2009).
На рисунках каждая делянка, в виде отрезка ряда длиной 10 м, имеет свой номер. На нее высаживалось обычно от 7 до 10 саженцев из потомства какого-либо дерева и в ее пределах растения генетически более однородны. Первые два растения высаживали через 0.5 м и остальные – через 1.0 м.
|
196 |
,98 |
’ "/99* |
|||||||||||
|
У/84* ‘ \ ' |
'85 \ |
,86 |
,87 7 |
188 |
,89 |
,90 / |
,91 X |
Х^з* * * |
|||||
|
467 \ |
469 \ |
72 |
473 / |
Д75 |
Д77 |
48/ |
|||||||
|
76 |
,7? |
,78 у |
,79 |
/ \ \ ^'"/83‘ / ' |
|||||||||
|
4/5 |
\ 4/7 / |
428 \ |
-V Д29 Y |
430 у |
43/ |
433 |
Х,^ 435/ / |
V3F Т |
\ Ч4У / |
||||
|
154 \ |
V */65 * * |
166 / |
* * 167 / |
168 / |
/69 |
/ко |
|||||||
|
/ \ \ |
|||||||||||||
|
\ ЗА7 / |
ЯА.9 / |
390 |
393 |
402 |
405 \ 1 |
у.— 2(Н |
|||||||
|
/59 |
j /60 |
• • /п- |
" /52 * |
||||||||||
|
*542 * * * |
\543 е е А* е |
* 5ДД Y" ’ |
5Д5 О-"---— |
■-----"347^ |
Х^ДЗ * * * |
* \ /55/ * * |
|||||||
|
/46 / |
/47 / |
/49 |
/50 ^Xz |
X /53 |
|||||||||
|
‘«К Г |
/459 * |
*46*0 / * * |
* * 46*1 7 * * * * |
* /*462 * * |
“дез * |
*468 * "Л |
X70* ’ " — |
||||||
|
угз Г |
422 * |
*432 * * |
*/ ЧзЬ^х^ |
./ .х^ \. |
|||||||||
|
у*1^ * / |
* "/25 |
"X /2/ |
/27 |
,28 |
129 |
УУо * |
/ /3/ |
/33* * |
|||||
|
* у(^ * * |
* цд *, |
/20 J |
/2/ |
||||||||||
|
"*5/" ЛЛ* |
52 / |
* \5: |
54 1 |
55 |
56 X |
X 58 |
59f |
" 50 ' / |
|||||
|
V |
\ 496 |
49$ |
59° < |
У 501 |
367/( ) |
368 / |
372 |
/373 |
375 / |
||||
|
Д1 * * ' |
* *43 |
* * /44 |
• 45 " |
Уб / |
47 / |
да \ |
/ Д9 |
50/ |
|||||
|
*46*7 ' * |
469 |
472 |
7 473 |
/ А77 / |
48ГХ |
482 ( J |
_ 483 |
Д8Д |
|||||
|
* * **з/ / ■ |
зК |
/ 34 |
35 |
—У |
36 |
37/ X |
38 / |
39 |
|||||
|
428 |
429 Y |
430 |
43/ |
483 |
X 435 / |
437 |
/ 440 |
||||||
|
*2/* * |
22 |
/ 22 |
23 / |
24 / |
А5 |
/26 |
X / |
28 |
/ 29 |
||||
|
356 / |
" 369 " ' ' " * * |
370 |
37/ / |
374 / |
J 376 |
• • • . |
' 377 |
379 |
380 |
/ 363 |
|||
|
/2 7 |
ХЗ |
лГг |
16 |
,8 / |
19 |
||||||||
О 5 10 15 20 м
Рис. 5. Фрагменты геобиологической сети Карри (усиленные узлы и связи между ними) и расположение растений в испытательных культурах ели в 21-летнем возрасте.
Одна точка соответствует одному растению в рядах культур, номера обозначают варианты опыта (номера потомств). Левый край участка ориентирован на северо-запад 38˚
|
495 |
о 196 |
/ 9? |
,98 |
368 |
"Ч?2 |
/ 202 * * |
||||||
|
496 |
500 \ |
/ 373 |
||||||||||
|
46^—— |
1,69 / |
Д72 j |
473 1 |
,79 |
/80 |
X / 482 ____^ |
Г 1,83 |
/83* * |
||||
|
"4/6" " |
4/7/ |
428 / |
*429 " * |
430 |
/43/ |
433 |
\ 435 |
437 |
440 |
|||
|
16^ ' |
ks * * |
,66 1 |
167 |
168 |
,70 Г |
\/7/ |
,72 |
,73 |
||||
|
384 |
^х/387 |
389 |
390 |
393 |
1 402 |
дозг |
Д08 |
|||||
|
*542 " " * |
* *543 * * " * |
‘ " ' 544 ..... |
*545 ‘ " " |
' *546 * * " |
1 547 |
5Д8 |
/ 5Д9 |
*550 * * * |
||||
|
"/44 " * ’ |
7 */*45* * * |
/46 * |
* */*47 Л |
/48 |
1 /49 |
/ /50 / /5/ |
152 |
153 |
||||
|
*423 * ' / |
Чу */*35* * |
462 |
*432 ✓ |
468 |
' 438 * * * * |
Д39 * * * * Y |
||||||
|
**/2*4 * / |
5 |
/27 |
,28 |
/3/ |
/32 , ' |
,33 |
||||||
|
* Нй/ * |
",15 Хч |
" " U6 ‘ |
117 " |
‘/,8 ' " |
*1/9 |
\ /20 \ |
/2/ |
/22 Ч |
123 |
|||
|
1 495 " *~" |
52 |
1 *499 * * |
*50*0 * * / |
*\ *50/ * / ' |
* * * 357* |
' " *368 |
372 |
/373* |
||||
|
■дб?" * |
464 |
-А73 * 7* * ' |
*47*7 * * ‘ |
" *37 " ' |
||||||||
|
*2/*\* "Л |
22 |
\ 22 |
\ / 24 |
261 |
27 / |
28 |
’9 |
|||||
|
3661 |
369 |
\ 370 |
\374 |
379 |
20 , _________ |
|||||||
X 10 /5 20 м
Рис. 6. Фрагменты геобиологической сети Хартмана (усиленные узлы и связи между ними) и расположение растений в испытательных культурах ели в 21-летнем возрасте.
Обозначения и пояснения те же, что и для рис. 5
Из высаженных растений на данном участке сохранилось 1648 шт. или 79.7%. При этом в узлы и на полосы сетей попало 157 растений, в том числе в сеть Хартмана 60 и в сеть Карри 73 саженца. Через
17 лет из них сохранилось 123 шт. или 78.3 % , в том числе в сети Хартмана сохранность составила 83.3%, а в сети Карри – 75.3%. Достоверного изменения сохранности в узлах ГБС не наблюдается и связано это, по-видимому, с высоким качеством посадочного материала – отсортированных саженцев.
В качестве контроля использованы растения, не попадающие в узлы и полосы ГБС. Контроль был двух видов: из 1–2 соседних деревьев делянки и из растений всей делянки, на которой выращивалось семенное потомство от свободного опыления какого-либо одного дерева. То есть сравнивались расте- ния только одного потомства (братья и сестры).
В зоне влияния полос и усиленных узлов сети Карри в культурах сохранилось 73 растения, которые мы измеряли, в том числе в полосах – 29, на краях узлов – 12 и в узлах 32 шт. Для последней группы (в узлах) приводим сравнение их высот с контролем (таблица). Далее в тексте мы приводим анализ средних высот растений в сетях ГБС в сравнении с высотами 1–2 соседних деревьев, как с контролем более строгим, чем контроль по средней высоте всех растений на делянке.
Высота растений в культурах ели в возрасте 21 год в усиленных узлах геобиологической сети Карри
|
Делянка, № |
Высота растений в усиленных узлах ГБС Карри |
Контроль: высота растений, не попадающих в узлы и полосы обеих ГБС, м |
||||||
|
м |
по отношению к контролю |
два соседних дерева |
на делянке |
|||||
|
из двух соседних деревьев |
из растений на делянке |
первое |
второе |
средняя |
число растений |
средняя высота |
||
|
194 |
7.0 |
0.92 |
1.14 |
7.4 |
7.9 |
7.65 |
4 |
6.15 |
|
6.0 |
0.78 |
0.98 |
||||||
|
501 |
4.6 |
0.81 |
0.77 |
6.7 |
4.7 |
5.70 |
6 |
5.98 |
|
4.2 |
0.74 |
0.70 |
||||||
|
368 |
2.6 |
0.44 |
0.39 |
5.6 |
6.1 |
5.85 |
3 |
6.63 |
|
372 |
4.7 |
1.07 |
1.00 |
4.2 |
4.6 |
4.40 |
4 |
4.70 |
|
174 |
4.9 |
0.71 |
0.71 |
|||||
|
4.8 |
0.69 |
0.70 |
7.1 |
6.8 |
6.95 |
1 |
6.90 |
|
|
180 |
7.2 |
1.03 |
1.22 |
6.4 |
7.6 |
7.00 |
4 |
5.93 |
|
428 |
6.5 |
0.92 |
0.92 |
6.9 |
7.2 |
7.05 |
3 |
7.07 |
|
5.3 |
0.75 |
0.75 |
||||||
|
429 |
4.4 |
0.66 |
0.80 |
6.4 |
7.0 |
6.70 |
7 |
5.49 |
|
384 |
6.8 |
1.33 |
1.17 |
5.1 |
5.1 |
5.10 |
5 |
5.80 |
|
4.6 |
0.90 |
0.79 |
||||||
|
544 |
5.3 |
0.85 |
1.01 |
5.5 |
6.9 |
6.20 |
3 |
5.23 |
|
545 |
5.8 |
1.05 |
1.05 |
5.6 |
5.5 |
5.55 |
5.55 |
|
|
547 |
5.5 |
0.99 |
0.87 |
5.9 |
5.2 |
5.55 |
5 |
6.34 |
|
134 |
2.6 |
0.48 |
0.50 |
4.1 |
6.8 |
5.45 |
4 |
5.23 |
|
423 |
5.3 |
0.88 |
1.00 |
6.0 |
6.0 |
6.00 |
4 |
5.33 |
|
425 |
3.8 |
0.94 |
0.66 |
3.6 |
4.5 |
4.05 |
5 |
5.78 |
|
426 |
4.9 |
0.78 |
0.85 |
4.9 |
7.7 |
6.30 |
||
|
6.0 |
0.95 |
1.04 |
||||||
|
499 |
4.5 |
0.68 |
0.96 |
7.2 |
6.1 |
6.65 |
1 |
4.70 |
|
500 |
4.2 |
0.88 |
0.68 |
4.0 |
5.5 |
4.75 |
4 |
6.15 |
|
469 |
5.2 |
1.18 |
0.96 |
4.0 |
4.8 |
4.40 |
4 |
5.40 |
|
482 |
5.6 |
0.95 |
0.92 |
5.9 |
5.90 |
3 |
6.10 |
|
|
3.5 |
0.59 |
0.57 |
||||||
|
32 |
5.5 |
1.03 |
0.95 |
6.2 |
4.5 |
5.35 |
2 |
5.80 |
|
4.5 |
0.84 |
0.78 |
||||||
|
36 |
4.8 |
0.86 |
0.99 |
4.8 |
6.4 |
5.60 |
4 |
4.85 |
|
428 |
5.5 |
0.97 |
0.88 |
5.7 |
5.6 |
5.65 |
4 |
6.25 |
|
429 |
4.4 |
0.67 |
0.85 |
6.6 |
6.60 |
4 |
5.18 |
|
|
Число |
32 |
- |
- |
24 |
22 |
46 |
84 |
- |
|
Среднее |
5.02 |
0.854 |
0.860 |
5.83 |
5.75 |
|||
Различия в высотах оказались достоверны: растения выросли низкорослыми и по сравнению с соседями в среднем оказались ниже на 0.81 м или на 14.0% (критерий достоверности различия Стьюдента tф= 3.34>t0.05=2.00), а по сравнению с растениями своего потомства (своей делянки) – ниже на 0.73 м или на 12.7%. (tф=3.25>t0.05=2.00). Снижение высот деревьев по краям узлов Карри оказалось меньше и составило в среднем 12.2%; еще меньше оно было на полосах сети (5.2%).
Вращение энергии по периферии узлов Карри было в 17 узлах против (–) и в 15 узлах – по часовой стрелке (+). Знак вращения не оказал влияния на высоту деревьев и для обоих случаев ее снижение оказалось одинаковым.
В сети Хартмана нами учтено 49 растений, в том числе в усиленных узлах 13 и в полосах между ними 36 шт. В полосах растения оказались ниже на 8.3% в сравнении с соседними деревьями, а в узлах – ниже на 13,4%. Достоверности различий близки к вероятности их доказательства на уровне 90–95% (критерии достоверности различия Стьюдента: t ф = 1.96 < t 0.05 = 1.98 для полос и t ф = 1.61 < t 0.10 = 1.69 для узлов). Вращение энергии по периферии узлов
Хартмана было в 8 узлах против (–) и в 5 – по часовой стрелке (+). Знак вращения не оказал существенного влияния на снижение высоты деревьев и для обоих случаев, по отношению к высоте растений контроля, оно оказалось близким: 13.3 и 13.6%.
Снижение средней высоты деревьев на усиленных узлах и в полосах ГБС Карри и Хартмана в сравнении с контролем показаны на рис. 7.
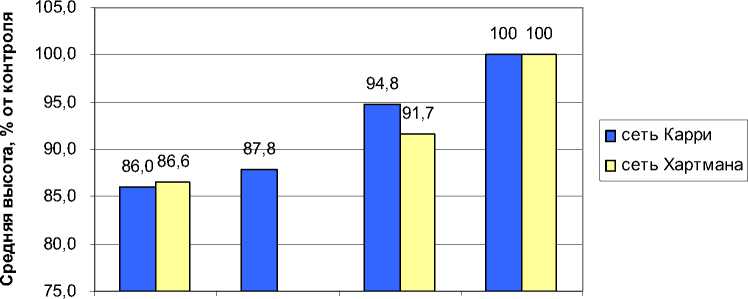
в узлах на краях в полосах контроль узлов
Рис. 7. Депрессия в росте ели в 21-летних культурах на усиленных узлах и полосах геобиологиче-ских сетей Карри и Хартмана
Нами изучаются и более сложные в поведении подвижные геобиологические сети с положительным влиянием на растения. Это структуры с узлами в диаметре по 1.0; 3.0 и 8.0 м, мигрирующие в течение месяца на несколько метров. Связи между узлами в них имеют рисунок в виде цепей, 6- и 4-х угольных структур в виде непрерывной сети. Первые два вида структур, возможно, приводят к формированию особенно крупных («плюсовых») деревьев в тех случаях, когда узлы в качестве активных биологических зон периодически возвращаются на свои прежние места.
Выводы
-
1. В культурах ели, созданных посадкой 4летними саженцами с целью испытания потомства, обнаружено отрицательное влияние геобиологиче-ских сетей Хартмана и Карри. В усиленных узлах этих сетей, имеющих диаметры от 0.55 до 2.0 м, растения в 21 год снижали рост в высоту в сравнении с контролем (высотами соседних деревьев своего потомства) в среднем на 13.4–14%. На краях узлов снижение было меньше (12.2%) и еще меньше –
-
2. Зоны сильного влияния данных сетей занимают 1.9%, а среднего и слабого влияния 10–11% площади. В опытах с делянками площадью 1–2 м2 попадание делянки в такие зоны может существенно понизить (на 5–14%) ее результаты. Необходимо исключение таких зон в опытах с выращиванием древесных растений в полевых и в лабораторных условиях.
на полосах сетей (5.2–8.3%). На сохранность растений данные сети влияния не оказали.
Список литературы Геобиологические сети Хартмана и Карри в испытательных культурах ели сибирской
- Поносов В.А. Биолокация и лозоходство. Пермь: Полиграфист, 1993. 54 с.
- Поносов В.А. и др. Экологический мониторинг в геофизике: учеб. пособие/под ред. В.А. Поносова. Пермь, 2009. 297 с.
- Рогозин М.В. Итоги 8-летних испытаний 525 семей ели сибирской в Пермском крае//Лесное хозяйство. 2008. ¢ 1. С. 37-38.
