Гетерогенность свойств копролитов дождевых червей Aporrectodea caliginosa и Lumbricus rubellus в модельном опыте на черноземе

Автор: Фролов О.А., Якушев А.В., Милановский Е.Ю.
Журнал: Бюллетень Почвенного института им. В.В. Докучаева @byulleten-esoil
Рубрика: Статьи
Статья в выпуске: 99, 2019 года.
Бесплатный доступ
Проведена оценка изменений физических и микробиологических свойств копролитов дождевых червей Aporrectodea caliginosa и Lumbricus rubellus в лабораторных опытах с использованием пахотного горизонта миграционно-мицелярного чернозема. В копролитах определяли физические параметры: гранулометрический состав, удельную поверхность, рассчитанную по эффективному диаметру частиц; микробиологические параметры: общую численность культивируемых бактерий и ферментативную кинетику липаз по модели Михаэлиса - Ментен с расчетом максимальной скорости реакции (Vm) и константы Михаэлиса (Km). Модельный эксперимент включал два этапа: первый этап - дождевые черви двух видов содержались вместе, во втором - раздельно. Контролем была исходная почва и почва с опадом. Анализировали и сравнивали с контролем копролиты и почву, в которой жили дождевые черви. Два вида дождевых червей дают неаддитивный эффект на физические и микробиологические свойства. В зависимости от видового состава характер изменения отличается...
Элементарные почвенные частицы, гранулометрический состав, удельная поверхность частиц, липазная активность
Короткий адрес: https://sciup.org/143168559
IDR: 143168559 | УДК: 631.4 | DOI: 10.19047/0136-1694-2019-99-92-116
The heterogeneity of the properties of the coprolites Aporrectodea caliginosa and Lumbricus rubellus in model experiment with chernozem soil
The changes in the physical and microbiological properties of the coprolites of earthworms Aporrectodea caliginosa and Lumbricus rubellus were evaluated in laboratory experiments using the arable horizon of migratory-mycelial chernozem. In coprolites the following physical parameters were determined: particle size distribution, specific surface area, calculated by the effective particle diameter; microbiological parameters: the total number of cultured bacteria and the enzymatic kinetics of lipases according to the Michaelis -Menten model with the calculation of the maximum reaction rate (Vm) and Michaelis constant (Km). The model experiment included two stages: the first stage - earthworms of two species were kept together, in the second - separately. The control was the source soil and the soil with litter. Coprolites and the soil in which earthworms lived were analyzed and compared with the control. Two types of earthworms give a non-additive effect on physical and microbiological properties...
Текст научной статьи Гетерогенность свойств копролитов дождевых червей Aporrectodea caliginosa и Lumbricus rubellus в модельном опыте на черноземе
Плодородие почв напрямую связано с жизнедеятельностью почвенной биоты (Звягинцев и др., 2005). Работ, посвященных изучению взаимосвязей в почве и модификаций почвенной биотой свойств почвы, достаточно много. Тем не менее методы и подхо- ды сильно варьируют. Например, почвенная биота рассматривается как центральный объект исследования (Чернов и др., 2017) или как действующий агент, влияющий на почву и формирующий ее свойства (Кутовая др., 2018; Bi et al., 2018). Можно рассматривать почвеннаую биоту как биоиндикатор изменений свойств почв (Frouz et al., 2013; Ma et al., 2013) или как ключевое звено в глобальных циклах вещества и энергии в экосистеме (Blouin et al., 2013; Edwards, 2004; Römbke et al., 2005). Дождевые черви являются одними из самых изучаемых представителей почвенной фауны (Huang, Xia, 2018; Peigné et al., 2018; Sanchez-Hernandez et al., 2018; Sanchez-Hernandez, 2018; Фролов, Якушев, 2018). Изучается их физиология (Laverack, 1963; Satchell, 1983), экология (Тихонов и др., 2011; Кутовая, 2012; Юрков и др., 2008; Храмченкова и др., 2012), использование в сельскохозяйственной отрасли (Бызов, Якушев, 2008). Однако в исследованиях обычно рассматриваются не собственно копролиты, а почва, где обитают черви. Более точное понимание вклада дождевых червей в формирование почвы возможно при разделении таких объектов как почва, где обитают черви, и непосредственно копролиты. Однако учитывая сложность и изменчивость копролитов во времени, их “старение” (Bal, 1970), подобное разделение осложнено и часто не проводится.
Целью работы является изучение модификации свойств элементарных почвенных частиц (ЭПЧ) при пассаже через кишечник дождевых червей Aporrectodea caliginosa и Lumbricus rubellus в модельном опыте. Для этого дана оценка гетерогенности внутри микрокосмов и между объектами; определены степени различий в свойствах копролитов и почвы, в которой обитают черви; выявлено, является ли значащим фактором видовая принадлежность дождевых червей для оценки свойств копролитов; сделано заключение о возможности достоверно отличить свойства копролитов от фоновой почвы.
ОБЪЕКТЫ И МЕТОДЫ
Для модельного опыта был выбран миграционно-мицелярный чернозем Курской области (51°37'17.1" N; 36°15'42.0" E). Данный тип почвы является одним из наиболее распрстраненных объектов исследования за счет высокой агроно- мической ценности. Образцы почвы после отбора хранились в воздушно-сухом состоянии. Перед закладкой опыта влажность почвы доводилась до полевой влагоемкости, почва перемешивалась и равномерно распределялась по микрокосмам, представляющим собой цилиндрические сосуды объемом 600 мл.
Кроме того, для исследования был выбран эндогейный червь Apporectodea caliginosa (собственно почвенный червь, обитающий в минеральной толще почвы (60–100 см) и питающийся опадом средней степени разложенности) и эпигейный червь Lumbricus rubellus (почвенно-подстилочный вид, обитающий на поверхности почвы и питающийся слаборазложившимся опадом). Отбор дождевых червей проводили согласно ISO 23611-1, видовую принадлежность червей устанавливали по определителю Все володовой – Перель (1997). Среди червей преобладали ювенильные особи. После эксперимента количество червей не изменилось.
Лабораторный эксперимент состоит из двух этапов. На первом этапе дождевых червей двух видов A. caliginosa и L. rubellus содержали совместно в сосудах с добавлением опада в виде свежих листьев клена остролистного ( Acer platanoides ). В качестве контроля в модельном опыте использовали сосуды с исходной почвой, а также с почвой и опадом. Таким образом, на первом этапе были проанализированы образцы: 1П – почва; 1ПО – почва с опадом; 1П-AL – почва из сосуда, где содержались черви A. caligi-nosa и L. rubellus ; 1К-AL – копролиты червей A. caliginosa и L. ru-bellus . Вес почвы в сосудах – 496 г. Общий вес червей – 12 г в равных долях.
Второй этап аналогичен первому, но виды червей содержались раздельно. На втором этапе проанализированы следующие образцы: 2П – почва; 2ПО – почва с опадом; 2П-A – почва из сосуда, где содержались черви A. caliginosa ; 2П-L – почва из сосуда, где содержались черви L. rubellus ; 2К-A – копролиты червей A. caliginosa ; 2К-L – копролиты червей L. rubellus . Вес почвы в сосудах с A. caliginosa – 470 г, с L. rubellus – 370 г. Вес червей A. calig-inosa – 16 г, L. rubellus – 18 г.
Экспериментальные сосуды содержали при температуре 17– 22 °С и весовой влажности почвы 35–40 % на протяжении четырех месяцев.
Для фракции частиц копролитов и почвы меньше 250 мкм определяли гранулометрический состав методом лазерной дифракции на дифрактометре Mastersize 3000 (предварительное просеивание через сито с размером ячеек 250 мкм и диспергация в течение 3 мин. ультразвуком, при энергии ультразвуковой диспер-гации 450 Дж/мл). Гранулометрический анализ проводили без удаления органических частиц при 5-кратной аналитической повторности по размерным фракциям: ил (< 1 мкм), мелкая пыль (1– 5 мкм), средняя пыль (5–10 мкм), крупная пыль (10–50 мкм), тонкий песок (50–250 мкм). Для каждой фракции рассчитывали среднее арифметическое (% объема) и доверительные интервалы при p = 0.95. Образцы, взятые для второго и первого этапа эксперимента, изначально имели небольшие отличия в гранулометрическом составе при оценке выбранным методом. Сравнение проводилось не по абсолютным значениям, а по соотношению долей фракций.
Удельную поверхность почвенных частиц рассчитывали по средневзвешенному диаметру частиц (установленному методом дифракции) из допущения, что частицы имеют форму сфер.
Общую численность культивируемых гетеротрофных аэробных и факультативно-анаэробных бактерий определяли чашечным методом Коха на среде “Rich” – разбавленной глюкозопептонной дрожжевой среде (ГПД). Пробоподготовка: почвенная суспензия (1 : 10), десорбция клеток на вортексе “Multi Reax” фирмы Heidolph 20 мин. при 2000 об./мин. Анализ численности культивируемых бактерий проводился в 3-кратной повторности; результаты представлены как медиана и квартили, так как при малой выборке непараметрический анализ является более корректным, хотя и менее точным. Ферментативная кинетика по Михаэлису – Ментен для липаз определялась по гидролизу флуоресцеина диацетата (ФДА) (Schnürer, Rosswall, 1982) . Для расчета константы Михаэлиса ( Km ) и максимальной скорости реакции ( Vm ) применялась серия растворов из 20 возрастающих концентраций ФДА (Бызов, Якушев, 2008) в диапазоне от 0 до 160 мкмоль/л. Каждая концентрация ФДА для каждого исследуемого образца была воспроизведена в 6-кратной повторности.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Контроль (почва и почва с опадом) по гранулометрическому составу не отличались. Опад в отсутствие животных не оказал никакого влияния на соотношение фракций.
В ходе первого этапа эксперимента в копролитах происходит увеличение доли фракций тонкого песка (50–250 мкм), и, как следствие, увеличивается удельная поверхность почвенных частиц.
□ копролиты A.caliginosa & L.terrestris
□ почва с червями A.caliginosa & L.rubellus
□ почва с опадом
□ почва
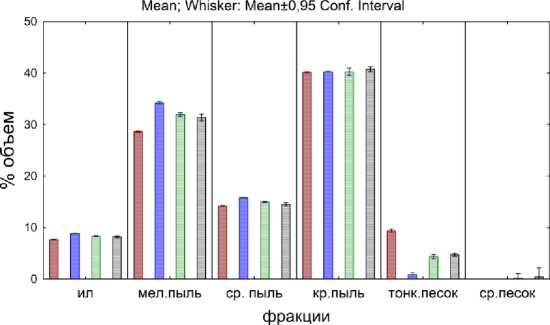
Рис. 1 . Гранулометрический состав первого этапа эксперимента.
Fig. 1. The soil grain size distribution at the first stage of the experiment.
На первом этапе эксперимента копролиты и почва, где жили черви, характеризуются меньшим разбросом значений (размаха 95 % интервала для среднего процентного объема), по сравнению с почвой и почвой с опадом. В копролитах происходит укрупнение частиц, а в почве, где жили черви, напротив, наблюдается повышение доли мелких частиц. Это хорошо заметно во фракции мелкой пыли и тонкого песка. Подобное перераспределение может иметь две причины: во-первых, оно связано с избирательностью дождевых червей, которые используют более крупные минераль- ные частицы для перетирания органических остатков в мышечном желудке (Перель, 1979); во-вторых, в кишечник дождевых червей и в копролиты попадают более крупные органические частицы опада, которыми питаются черви. Учитывая разную экологию выбранных видов дождевых червей, появилось предположение о специфике их питания – большей частью минеральными частицами (A. caliginosa) и слаборазложившимися растительными остатками (L. rubellus).
Таким образом, значимым фактором, оказывающим влияние на свойства копролитов, является видовая принадлежность дождевых червей. Чтобы это подтвердить, на втором этапе эксперимента дождевые черви содержались раздельно.
-
□ копролиты A. caliginosa
-
□ почва с A. caliginosa
-
□ копролиты L.rubellus
3 почва с L.rubellus
-
□ почва
-
□ почва с опадом
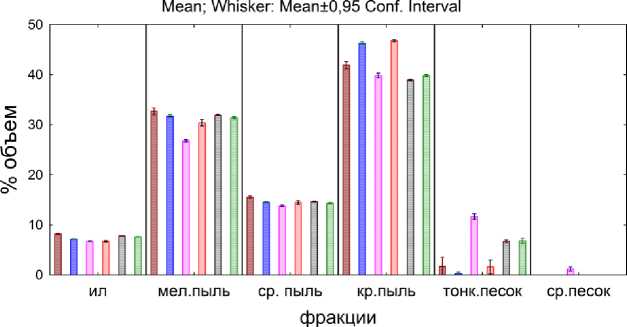
Рис. 2. Гранулометрический состав второго этапа эксперимента.
Fig. 2. The soil grain size distribution at the second stage of the experiment.
Гетерогенность копролитов и почвы, где жили черви, заметно выше, чем в контроле (почва и почва с опадом). Копролиты A. caliginosa по гранулометрическому составу состоят из частиц меньших, чем почва, в которой жили черви. Это заметно во фракциях ила, мелкой пыли, средней пыли. Процентное содержание мелких частиц в этих фракциях для копролитов выше, а для почвы, где жили черви, ниже. Во фракции крупной пыли наблюдается существенное увеличение частиц, по сравнению с контролем (на ~ 3 %). При этом увеличение содержания частиц большего размера наблюдается в почве, где жили черви, по сравнению с копролита-ми (на ~ 4.4 %). Частиц фракции тонкого песка практически нет как в копролитах, так и в почве, где жили черви A. caliginosa .
Иная картина наблюдается для L. rubellus (копролиты и почва, где жили черви). Во фракциях ила и мелкой пыли, содержание частиц ниже, чем в контроле (для фракции ила уменьшение на ~ 1 %, и для копролитов, и для почвы, где жили черви; для фракции мелкой пыли уменьшение составило ~ 4.5 % для копролитов и ~ 1 % для почвы, где жили черви). При этом доля фракции мелкой пыли в копролитах ниже, чем в почве, где жили черви. Фракция средней пыли не отличается от контроля. Во фракции крупной пыли почва с червями содержит большее количество частиц, чем в контроле и копролитах. Интерес представляет то, что в копроли-тах L. rubellus во фракции тонкого песка сдержится намного больше частиц, чем в контроле и почве, где жили черви (последняя представленна единичными частицами из этой фракции). Вероятно, это связано с присутствием в копролитах органических частиц из опада. Наибольшее же внимание привлекает наличие в копролитах частиц крупнее 250 мкм. Это частицы, которых не было в образцах исходной почвы из-за специфики пробоподготовки (просеивание образцов через сито 250 мкм). В данном случае частицы крупнее 250 мкм представляют собой органический материал, который переработали черви (по данным микроскопирования). Предположительно, из-за своей несферичности подобные частицы смогли пройти сквозь сито. Поскольку такие частицы являются результатом преобразования опада, прошедшего через кишечник дождевых червей, они найдены именно в копролитах.
Наличие опада на поверхности почвы (вариант ПО) не оказало влияния на гранулометрические показатели. Из чего можно заключить, что все зафиксированные отличия в гранулометрическом составе произошли за счет деятельности дождевых червей.
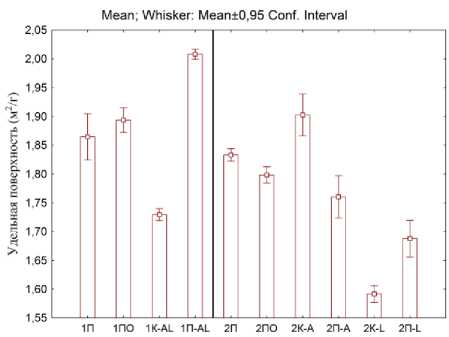
Рис. 3 . Удельная поверхность для двух этапов эксперимента: слева (первый этап 1П – почва, 1ПО – почва с опадом, 1K-AL – копролиты двух видов, 1П-AL – почва, где жили черви), справа (второй этап 2П – почва, 2ПО – почва с опадом, 2K-A – копролиты A. caliginosa ; 2П-А – почва, где жили A. caliginosa , 2K-L – копролиты L. rubellus ; 2П-L – почва, где жили L. rubellus ).
Fig. 3. The specific surface for the two stages of the experiment: on the left (first stage 1П – soil, 1ПО – soil with litter, 1K-AL – two types of coprolites, 1П-AL – soil where the worms lived), on the right (second stage 2П – soil, 2ПО – soil with litter, 2K-A – coprolites of A. caliginosa ; 2П-А – the soil where A. caliginosa lived, 2K-L – coprolites of L. rubellus ; 2П-L – soil where L. rubellu s lived).
Удельная поверхность почвы (рис. 3) рассчитана исходя из данных гранулометрического анализа. Для совместного содержания двух видов дождевых червей характерно уменьшение удельной поверхности в копролитах и увеличение в почве, где жили черви, по сравнению с контролем (т. е. после прохождения через кишечник наблюдалась бóльшая агрегированность почвы). Для вида A. caliginosa наблюдается увеличение удельной поверхности копролитов, по сравнению с контролем (т. е. черви размельчили почву, пропуская ее через кишечник). Почва, где жил A. caliginosa практически не отличается от почвы контроля. Для L. rubellus наблюдается уменьшение удельной поверхности, как для копролитов, так и для почвы, где жил L. rubellus
(предположительно за счет массового добавления к почве органических частиц опада). При этом удельная поверхность копролитов ниже, чем почвы.
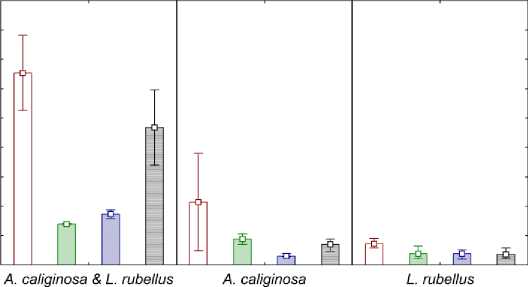
Анализируя микробиологические данные можно отметить, что для копролитов во всех вариантах опыта наблюдается увеличение численности бактерий (рис. 4). На рисунке первая треть отображает численность для первого этапа эксперимента ( A. caliginosa и L. Rubellus , содержавшиеся вместе), а последующие две трети – для второго этапа ( A. caliginosa и L. Rubellus , содержавшиеся раздельно). Для каждого определения показателей численности брался свой контроль (почва без червей и почва только с опадом).
Общая численность культивируемых бактерий @ копролиты @ почва 0 почва с опадом 0 почва с червями
Median; Whisker: 25%-75% 4,5Е9 4Е9
3.5Е9 ЗЕ9
2.5Е9 2Е9
1.5Е9 1Е9 5Е8 О
Рис. 4. Общая численность культивируемых бактерий. Fig. 4. The total number of cultivated bacteria.
Несмотря на то, что точность метода микробиологического посева позволяет определять изменение численности при разнице более чем на порядок, можно сказать, что наибольшее увеличение численности наблюдается при содержании двух видов дождевых червей вместе. Также наблюдается тенденция усиления этого эф- фекта в ряду L. rubellus и A. caliginosa > A. caliginosa > L. rubellus.
Проведена оценка липазной активности, на основании реакции гидролиза ФДА (флуоресцеина диацетата). На рисунке 7 приведен пример экспериментально полученных кривых зависимости скорости фементативной реакции от концентрации субстрата реакции, используемых для расчета кинетических параметров V m и Km липаз с помощью аппроксимации экспериментальных данных кинетической математической моделью Михаэлиса – Ментен.
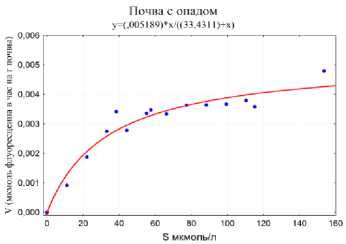
Рис. 5. Пример кривой насыщения липаз субстратом ферментативной реакции (ФДА), используемой для расчета кинетических параметров ферментов по модели Михаэлиса – Ментен.
Fig. 5. An example of a lipase saturation curve with an enzyme reaction substrate (FDA) used to calculate the kinetic parameters of enzymes according to the Michaelis – Menten model.
На основании подобных кривых рассчитаны значения Vm (рис. 6) и K m (рис. 7) для липаз.
Для первого этапа эксперимента характерно увеличение Vm в копролитах и почве, где содержались черви, что говорит о повышении дождевыми червями активности микроорганизмов при содержании двух видов червей вместе. При этом увеличение в копролитах ниже, чем в почве с L. rubellus и A. сaliginosa . Это может быть связано с действием кожных выделений дождевых червей. Во второй части эксперимента при раздельном содержании червей разных видов наблюдается увеличение Vm для копролитов A. сaliginosa , по сравнению с контролем, и уменьшение в почве, где жили A. caliginosa.
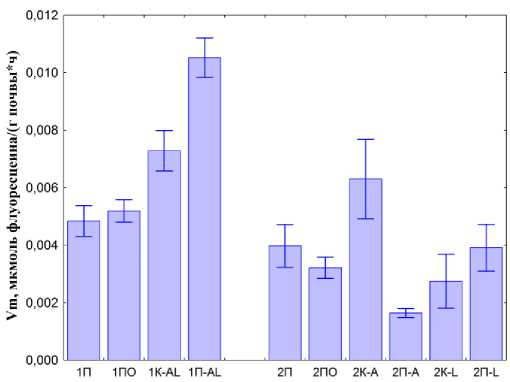
Рис. 6. Максимально возможная скорость гидролиза ( Vm ) ФДА, осуществляемого липазами при полном насыщении их активных центров субстратом реакции (первый этап 1П – почва, 1ПО– почва с опадом, 1K-AL – копролиты двух видов, 1П-AL – почва, где жили черви), справа (второй этап 2П – почва, 2ПО – почва с опадом, 2K-A – копролиты A. caliginosa ; 2П-А – почва, где жили A. caliginosa , 2K-L – копролиты L. rubellus ; 2П-L – почва, где жили L. rubellus ).
Fig. 6. The maximum possible rate of hydrolysis (Vm) of FDA carried out by lipases at full saturation of their active centers with the reaction substrate (first stage 1П – soil, 1ПО – soil with litter, 1K-AL – two types of coprolites, 1П-AL – soil where the worms lived), on the right (second stage 2П – soil, 2ПО – soil with litter, 2K-A – coprolites of A. caliginosa ; 2П-А – the soil where A. caliginosa lived, 2K-L – coprolites of L. rubellus ; 2П-L – soil where L. rubellu s lived).
В копролитах L. rubellus происходит снижение V m , а почва, где жил L. Rubellus , достоверно не отличается от контроля. Значения Vm вариантов контроля с почвой и почвой с опадом практически не отличаются. Присутствие опада не дает изменения липазной активности, соответственно, все изменения вызваны деятельностью дождевых червей.
Таким образом, можно сделать вывод, что Vm уменьшается в ряду копролитов L. rubellus и A. сaliginosa > A. caliginosa > L. rubellus. Это явление подтверждается увеличением общей чис- ленности бактерий в копролитах, достигающей своего максимума при совместном содержание дождевых червей.
Важно заметить, что абсолютные значения липазной активности ( Vm ) существенно ниже в нашем эксперименте, чем они представлены в литературных данных (Киреева и др., 2006; Zhimin et al., 2015) . Причины столь низких значений, возможно, кроются в длительном хранении исходной почвы в воздушно-сухом состоянии. Для некоторых ферментов подобные условия хранения могут привести к значительному снижению исходной ферментативной активности (Даденко, Казеев, 2004) .
Эффективность работы ферментов характеризует константа полунасыщения Михаэлиса K m (рис. 7). Для первого этапа эксперимента характерно незначительное увеличение K m для копролитов и почвы, где жили черви. При этом увеличение в копролитах ниже чем в почве с L. rubellus и A. сaliginosa. Из этого можно сделать предположение, что два вида дождевых червей мало влияют на эффективность работы липазы.
На втором этапе наблюдается увеличение Km для копроли-тов A. caliginosa и, особенно, для L. rubellus и уменьшение K m в почве, где жил A. сaliginosa, по сравнению с контролем . Эффективность работы ферментов в почве, где жили L. rubellus , увеличивается не столь сильно и достигает тех же значений, что и в ко-пролитах A. caliginosa .
Контроль с почвой и почвой с опадом существенно не отличаются по данному показателю. Поскольку присутствие опада не дает изменения K m , значит, все изменения вызваны деятельностью дождевых червей. Таким образом, можно заключить, что по отдельности каждый вид дождевых червей повышает эффективность работы фермента липаз, но при совместном содержании эффективность фермента падает, однако значительно возрастает скорость ферментативной активности.
Для интегрального анализа полученных физических и микробиологических параметров и для выявления скрытых биологических факторов был проведен дискриминантный анализ отдельно для физических и микробиологических параметров.
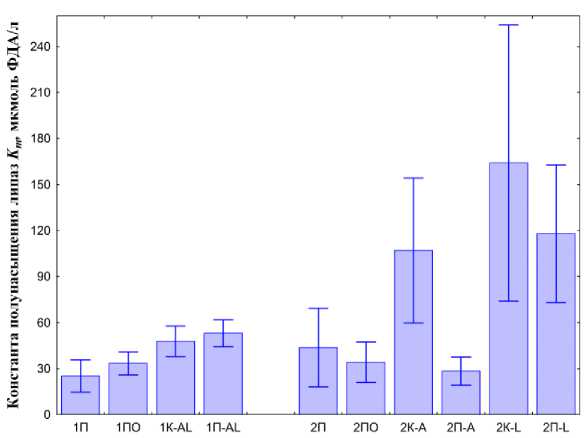
Рис. 7. Сродство липаз к субстрату, оцененное по константе полунасыщения Михаэлиса ( Km ) (первый этап 1П – почва, 1ПО – почва с опадом, 1K-AL – копролиты двух видов, 1П-AL – почва, где жили черви), справа (второй этап 2П – почва, 2ПО – почва с опадом, 2K-A – копролиты A. caliginosa ; 2П-А – почва, где жили A. caliginosa , 2K-L – копролиты L. rubellus ; 2П-L – почва, где жили L. rubellus ).
Fig. 7. The affinity of lipase to the substrate, estimated by the Michaelis constant of half-saturation (Km) (first stage 1П – soil, 1ПО – soil with litter, 1K-AL – two types of coprolites, 1П-AL – soil where the worms lived), on the right (second stage 2П – soil, 2ПО – soil with litter, 2K-A – coprolites of A. caliginosa ; 2П-А – the soil where A. caliginosa lived, 2K-L – coprolites of L. rubellus ; 2П-L – soil where L. rubellu s lived).
В ходе анализа был выявлен только один статистически достоверный биологический фактор, действующий на физические и микробиологические свойства и формально математически выраженный как дискриминационная функция 1 (ДФ1микр – для микробиологических свойств; ДФ1 физ – для физических свойств).
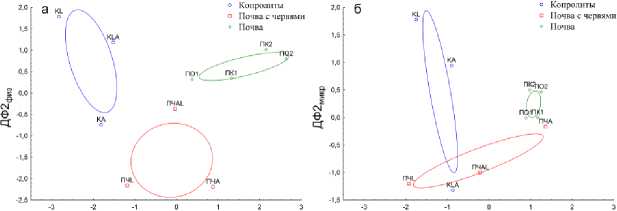
Вторая дискриминантная функция является вспомогательной, математически обусловленной проводимым анализом. По взаиморасположению объектов исследования в двумерном факторном пространстве, образованном пересечением ДФ1 и ДФ2
(рис. 8), можно заключить, что этот фактор – влияние дождевых червей на физические и микробиологические свойства субстратов. Так как слева направо на рисунке 8 наблюдается разделение объектов в ряд от копролитов, через почву, в которой жили черви, до контрольных вариантов почвы.
ДЧЧфи! ДФ1-«р
Рис. 8. Взаиморасположению объектов исследования в двумерном факторной пространстве, образованном пересечением ДФ1 и ДФ2, полученных в ходе дискриминантного анализа физических (а) и микробиологических (б) параметров.
Fig. 8. The interposition of the objects of study in a two-dimensional factor space, formed by the intersection of ДФ1 and ДФ2, obtained in the course of discriminant analysis of physical (a) and microbiological (b) parameters.
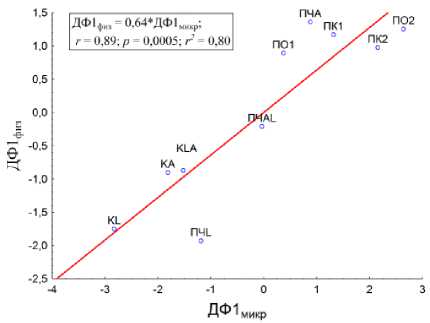
Рис. 9. Корреляция между ДФ1 микр. и ДФ1 физ. .
Fig. 9. Correlation between ДФ1микр. and ДФ1 ф из..
Проведенный корреляционный анализ (рис. 9) для ДФ1микр. и ДФ1 физ. показал идентичность действия фактора (влияние пассажа почвы через кишечник дождевых червей) на объекты исследования, что доказывает наличие корреляции между исследуемыми микробиологическими и физическими свойствами.
Для того чтобы понять роль этой корреляционной зависимости были проанализированы стандартизированные коэффициенты при физических и микробиологических параметрах, входящих в алгебраическое выражение ДФ1 физ. и ДФ1 микр. соответственно. Активность липаз ( Vm ) прямо пропорционально коррелирует с удельной поверхностью и обилием бактерий (КОЕ/г почвы) (рис. 10).
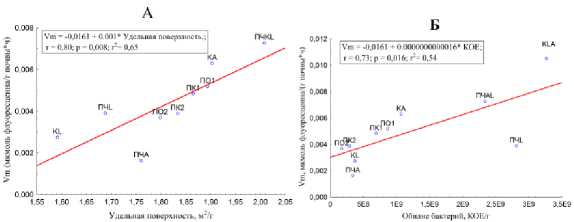
Рис. 10. Корреляция между Vm и удельной поверхностью почвы и копролитов (а) и обилием бактерий (б).
Fig. 10. The correlation between Vm and the specific surface of the soil and coprolites (a) and the abundance of bacteria (b).
Эффективность фермента ( К m ) обратно пропорциональна отношению мелкой пыли и ила к крупной пыли (рис. 11). Чем обусловлена такая корреляция, до конца не ясно, но, вероятно, это связанно с тем, что изменения, произошедшие именно в этих фракциях, существенно влияют на эффективность липаз, протекание биохимических реакций.
Однако отметим, что наличие корреляции не означает наличие прямых причинно-следственных связей между физическими и микробиологическими явлениями. Поэтому для доказательства существования подобных связей мы попытались предположить механизм подобных взаимодействий.
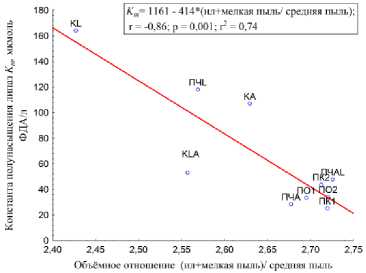
Рис. 11. Корреляция между K m липаз и отношением ил с мелкой пылью к средней пыли.
Fig. 11. The correlation between K m lipase and the ratio of sludge and fine dust to medium dust.
Увеличение удельной поверхности почвы и копролитов приводит к увеличению площади поверхности, доступной для иммобилизации внеклеточных липаз, которые стабилизируются и сохраняют свою активность преимущественно в адсорбированном состоянии. Обилие липаз связано с обилием гетеротрофных бактерий, что указывает на преимущественно микробное происхождение этих ферментов и недолгое сохранение их активности в почве (вне клеток), что и обеспечивает наблюдаемую тесную корреляцию. Уменьшение эффективности работы липаз при увеличении фракции средней пыли (ярко наблюдаемое для L. rubellus увеличение во фракции тонкого песка и снижение активности фермента ( Vm )) обусловлено привнесением червями пористых органических частиц размера средней пыли из опада (подобные частицы наблюдаются в световой микроскоп в почвенной суспензии), на которых липазы адсорбируются, в том числе и на внутренних поверхностях, для которых затруднена диффузия субстрата реакции к активным центрам липаз, иммобилизованных внутри частиц. Это приводит к увеличению Km .
ЗАКЛЮЧЕНИЕ
Данные эксперимента охватывают гетерогенность внутри микрокосмов и позволяют говорить о порядке величин наблюдае- мых эффектов, а также позволяют выделить параметры, которые следует рассмотреть в качестве базовых для проведения дальнейших исследований, направленных на изучение свойств копролитов дождевых червей. На основании полученных результатов можно заключить, что происходит изменение гранулометрического состава и удельной поверхности при жизнедеятельности дождевых червей в почве с опадом. Наблюдается увеличение численности микроорганизмов и ферментативной активности в копролитах и почве с дождевыми червями. Разные виды червей оказывают разное влияние, а степень и характер влияния неаддитивные. Это связанно с различными экологическими группами выбранных видов. A. caliginosa – истинно почвенный (эндогейный) червь. Он в большей степени питается органическими остатками, которые находятся в толще почвы. В то время как L. rubellus – подстилочный (эпигейный) червь. Для него характерно обитание в подстилке и верхнем слое почвы. Следовательно, и преобразование опада, а также функции этих видов в почве различны. Вероятно, именно это приводит к столь сильным отличиям в определяемых параметрах. Неаддитивность же эффекта стоит отнести к устойчивости экосистем. Чем выше биоразнообразие, тем устойчивее система. Усиление эффектов при совместном содержании двух видов иллюстрирует этот принцип.
Копролиты дождевых червей отличаются как от почвы без червей, так и от почвы, в которой они обитают. Наличие опада в почве без червей практически не оказывает влияние на изученные параметры. Это может указывать на то, что в природе значительная роль в активизации почвенной микробиоты отводится не только растительным остаткам, поступающим на поверхность почвы, существенную роль играет почвенная фауна, которая размельчает листовой опад, инициирует и активизирует переработку органических веществ почвенными микроорганизмами.
Одним из важных выводов можно считать экспериментально подтвержденную низкую неоднородность (разброс значений) определяемых физических и микробиологических параметров, по сравнению с различиями между исследуемыми объектами. Это позволит проверять гипотезы и предположения, а также статистически обосновывать выводы с дальнейшей экстраполяцией на
природные экосистемы.
Ecotoxicology and Environmental Safety , 2005, Vol. 62, Issue 2, pp. 249–265, DOI: 10.1016/j.ecoenv.2005.03.027.
Список литературы Гетерогенность свойств копролитов дождевых червей Aporrectodea caliginosa и Lumbricus rubellus в модельном опыте на черноземе
- Бызов Б.А., Якушев А.В. Микробиологическая характеристика вермикомпостирования методом мультисубстратного тестирования // Почвоведение. 2008. № 11. С. 1381-1387. DOI: 10.1134/S1064229308110112
- Всеволодова-Перель Т.С. Дождевые черви: Кадастр и определитель. М.: Наука. 1997. 102 c.
- Даденко Е.В., Казеев К.Ш. Влияние различных сроков и способов хранения почвенных образцов на ферментативную активность чернозема // Известия высших учебных заведений. Северо-Кавказский регион. Естественные науки. 2004. № 6. С. 61-65.
- Звягинцев Д.Г., Бабьева И.П., Зенова Г.М. Биология почв. М.: Изд-во Моск. ун-та, 2005. 445 с.
- Киреева Н.А., Маркарова М.Ю., Щемелинин Т.Н., Рафикова Г.Ф. Ферментативная и микробиологическая активность загрязненных нефтью глееподзолистых почв на разных стадиях их восстановления // Вестник Башкирского университета. 2006. Т. 11. № 4. С. 56-60.
- Кутовая О.В. Характеристика гумусовых веществ агродерново-подзолистой почвы и копролитов дождевых червей // Бюллетень Почвенного института имени В.В. Докучаева. 2012. Вып. 69. С. 46-59.
- DOI: 10.19047/0136-1694-2012-69-46-59
- Кутовая О.В., Гребенников А.М., Тхакахова А.К., Исаев В.А., Гармашов В.М., Беспалов В А., Чевердин Ю.И., Белобров В.П. Изменение почвенно-биологических процессов и структуры микробного сообщества агрочерноземов при Разных способах обработки почвы // Бюллетень Почвенного института имени В.В. Докучаева. 2018. Вып. 92. С. 35-61.
- DOI: 10.19047/0136-1694-2018-92-35-61
- Перель Т.С. Распространение и закономерности распределения дождевых червей фауны СССР. М.: Наука, 1979. 272 c.
- Тихонов В.В., Бызов Б.А., Завгородняя Ю.А., Демин В.В. Дождевые черви - преобразователи структуры и биологической активности гуминовых кислот // Изв. РАН. Сер. биологическая. 2011. № 1. С. 24-32.
- DOI: 10.1134/S1062359010061032
- Фролов О.А., Якушев А.В. Влияние на бактериальный гидролитический комплекс гумусо-аккумулятивного горизонта техноурбанозема пассажа через кишечник дождевого червя Aporrectodea caliginosa // Бюллетень Почвенного института имени В.В. Докучаева. 2018. Вып. 94. С. 57-73.
- DOI: 10.19047/0136-1694-2018-92-57-73
- Храмченкова О., Веремеев В., Бачура Ю. Водоросли почв и копролитов дождевых червей в луговых экосистемах // Наука и инновации. 2012. № 108. С. 67-70.
- Чернов Т.И., Холодов В.А., Когут Б.М., Иванов А.Л. Методология микробиологических исследований почвы в рамках проекта "Микробиом России" // Бюллетень Почвенного института имени В.В. Докучаева. 2017. Вып. 87. С. 100-113.
- DOI: 10.19047/0136-1694-2017-87-100-113
- Юрков А.М., Чернов И.Ю., Тиунов А.В. Влияние дождевых червей Lumbricus terrestris на структуру дрожжевого сообщества лесной подстилки // Микробиология. 2008. Т. 77. № 1. С. 121-125.
- DOI: 10.1134/S0026261708010153
- Bal L. Morphological investigation in two moder-humus profiles and the role of the soil fauna in their genesis // Geoderma. 1970. Vol. 4. Issue 1. P. 5-36.
- DOI: 10.1016/0016-7061(70)90030-3
- Bi Y.M., Tian G.L., Wang C., Zhang Y., Wang D.N., Zhang F.F., Sun Z.J. Differential effects of two earthworm species on Fusarium wilt of strawberry // Appl. Soil Ecology. 2018. Vol. 126. P. 174-181.
- DOI: 10.1016/j.apsoil.2018.02.024
- Blouin M., Hodson M.E., Delgado E.A., Baker G., Brussaard L., Butt K.R., Cluzeau D. A review of earthworm impact on soil function and ecosystem services // European Journal of Soil Science. 2013. Vol. 64. Vol. 2. P. 161-182.
- DOI: 10.1111/ejss.12025
- Edwards C.A. Earthworm ecology. CRC press. 2004. 456 p.
- Filser J., Faber J.H., Tiunov A.V., Brussaard L., Frouz J., Deyn G.D., Uvarov A.V., Berg M.P., Lavelle P., Loreau M., Wall D.H., Querner P., Eijsackers H., Jiménez J.J. Soil fauna: key to new carbon models // Soil. 2016. No. 2. P. 565-582.
- DOI: 10.5194/soil-2-565-2016
- Frouz J., Livečková M., Albrechtová J., Chroňáková A., Cajthaml T., Pižl V., Šimáčková H. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites // Forest Ecology and Management. 2013. Vol. 309. P. 87-95.
- DOI: 10.1016/j.foreco.2013.02.013
- Huang K., Xia H. Role of earthworms' mucus in vermicomposting system: Biodegradation tests based on humification and microbial activity // Sci. Total Environ. 2018. Vol. 610-611. P. 703-708.
- DOI: 10.1016/j.scitotenv.2017.08.104
- Laverack N.S. Mechanistic mathematical models of microbial growth in bioreactors and in natural soils: explanation of complex phenomena // Mathematics and Computers in Simulation. 1996. Vol. 42. Issue 2-3. P. 179-186.
- DOI: 10.1016/0378-4754(95)00127-1
- Ma Y., Filley T.R., Johnston C.T., Crow S.E., Szlavecz K., McCormick M.K. The combined controls of land use legacy and earthworm activity on soil organic matter chemistry and particle association during afforestation // Organic geochemistry. 2013. Vol. 58. P. 56-68.
- DOI: 10.1016/j.orggeochem.2013.02.010
- Peigne J., Vian J.F., Payet V., Saby N.P. Soil fertility after 10 years of conservation tillage in organic farming // Soil and Tillage Res. 2018. Vol. 175. P. 194- 204.
- DOI: 10.1016/j.still.2017.09.008
- Römbke J., Jänsch S., Didden W., Römbke J., Jänsch S., Didden W. The use of earthworms in ecological soil classification and assessment concepts // Ecotoxicology and Environmental Safety. 2005. Vol. 62. Issue 2. P. 249-265.
- DOI: 10.1016/j.ecoenv.2005.03.027
- Sanchez-Hernandez J.C. Biochar activation with exoenzymes induced by earthworms: A novel functional strategy for soil quality promotion // J. Hazardous Materials. 2018. Vol. 350. P. 136-143.
- DOI: 10.1016/j.jhazmat.2018.02.019
- Sanchez-Hernandez J.C., del Pino J.N., Capowiez Y., Mazzia C., Rault M. Soil enzyme dynamics in chlorpyrifos-treated soils under the influence of earthworms // Sci. Total Environ. 2018. Vol. 612. P. 1407-1416.
- DOI: 10.1016/j.scitotenv.2017.09.043
- Satchell J.E. Earthworm microbiology // Earthworm ecology from Darwin to vermiculture. London, N.Y, 1983. P. 351-364.
- DOI: 10.1007/978-94-009-5965-1
- Schnürer J., Rosswall T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter // Appl. Environ. Microbiol. 1982. Vol. 43(6). P. 1256-1261.
- Zhimin Y.U.A.N., Haijun L.I.U., Jun H.A.N., Jingjing S.U.N., Xiaoying W.U., Jun, Y.A.O. Monitoring soil microbial activities in different cropping systems using combined methods // Pedosphere. 2017. Vol. 27. No. 1. P. 138-146.
- DOI: 10.1016/S1002-0160(15)60100-X
